甘蓝型油菜BnFUL基因家族全基因组鉴定与表达分析
2022-03-16刘营江姚兰尹泽刘佳张大为吴金锋邹子湘周定港严明理
刘营,江姚兰,尹泽,刘佳,张大为,吴金锋,邹子湘,周定港*,严明理*
(1.湖南科技大学生命科学学院,湖南 湘潭,411201;2.经济作物遗传改良与综合利用湖南省重点实验室,湖南 湘潭,411201;3.中国农业科学院油料作物研究所,湖北 武汉,430062)
甘蓝型油菜作为我国重要的油料作物,常年种植面积在700 万公顷(约1 亿亩)以上,具有适应性强,生长速度快,生物量大等优点[1]。随着农业机械化的要求越来越高,抗裂角资源的匮乏严重限制了机械化收割。因此,挖掘调控抗裂角性状相关基因,对油菜的抗裂改良意义重大。
FUL(FRUITFULL)基因属于MADS-box 转录因子家族的一员,广泛存在于真核生物中,在植物的生长发育中,特别是在被子植物的花器官分化、开花时间的调节以及果实的发育和成熟等方面起重要作用[2,3]。MADS-box 家族依据进化关系分为两种类型:type I 型和type II 型[4]。FUL基因属于type II型(又名MIKC型),通常在N端(M)具有高度保守的DNA 结合域(MADS-box),一般56~60 个氨基酸[5],“K”为第二保守结构域,具有螺旋结构,有助于蛋白之间形成二聚体,“I”和“C”结构域则保守性较低[6]。
FUL基因在模式植物拟南芥中研究较为深入,在拟南芥中仅有1 个FUL基因,且为第一个鉴定出的与拟南芥角果开裂有关的基因[7]。FUL基因在拟南芥的早期发育过程中不表达,在成熟期高表达;且突变体植株角果内皮层细胞增多,成熟的种子自动从角果中脱落[8];而过表达AtFUL基因的拟南芥植株中,角果皮不能形成离区,成熟时角果皮不开裂[9]。此外,FUL基因还参与调控拟南芥的叶片发育[10]、花序分化[11]、开花时间[12]和角果伸长[8]等。对油菜FUL基因的研究中,异位表达AtFUL基因能显著增强芥菜型油菜角果的抗裂性[13];在甘蓝型油菜中过表达与拟南芥FUL同源的白芥MADSB基因也提高了角果的抗裂性[14]。过表达GmFULa增加了大豆产量,而未改变开花时间和成熟度[15];番茄TDR4/FUL1和MBP7/FUL2这两个基因在无乙烯的条件下,参与细胞壁修饰并影响果实成熟[16];黄瓜中,FUL的旁系同源基因Csa004592和Cs004120通过控制细胞分裂与细胞大小,从而调节黄瓜果实的长度[17]。
基于甘蓝型油菜的基因组序列,本研究在全基因组水平上系统鉴定甘蓝型油菜BnFUL 基因家族成员,并对其基本理化性质、蛋白结构、基因结构、系统进化关系及共线性关系等进行分析。同时,基于转录组表达数据对该家族基因在不同品种、不同发育时期和不同部位的表达特性进行分析,以期为甘蓝型油菜抗裂基因的功能阐释和抗裂角种质资源的培育奠定基础。
1 材料与方法
1.1 数据来源
基因组数据库来自拟南芥基因组[18](https://www. arabidopsis. org/)和甘蓝型油菜基因组[19](http://cbi. hzau. edu. cn/bnapus/index. php)。表达谱数据源自甘蓝型油菜转录组数据库[20](http://yanglab.hzau.edu.cn/BnTIR)。
1.2 BnFUL基因家族的鉴定
以拟南芥AtFUL(AT5G60910.1)基因为种子序列进行本地blastp(参数为:1e-5),以初筛选序列绘制进化树,结合Pfam(http://pfam. xfam. org/)和SMART(http://smart. emblheidelberg. de/)分析结构域,最终将与AtFUL聚为一支且均含有K-box 结构域和MADS_MEF2_like结构域的成员确定为甘蓝型油菜BnFUL基因。鉴定所得成员,根据其在染色体的位置命名。
1.3 甘蓝型油菜FUL 家族基因序列分析及蛋白特性分析
利用GSDS[21](http://gsds. gao-lab. org/)对甘蓝型油菜FUL 基因家族进行结构分析。利用MEME[22](https://meme-suite. org/meme/tools/meme)进行蛋白保守基序预测。利用NCBI-CDD[23](https://www.ncbi. nlm. nih. gov/Structure/bwrpsb/bwrpsb. cgi)工具进行BnFUL 蛋白家族保守结构域分析。利用在线网站ExPASY(https://www. expasy. org/)和SOMPA(https://prabi.ibcp.fr/htm/site/web/home)对鉴定所得成员的蛋白序列进行一级结构和二级结构分析。利 用 PLANTCARE[24](http://bioinformatics. psb.ugent. be/webtools/plantcare/html/)进行启动子区域顺式作用元件分析。利用在线网站WoLF PSOORT(https://wolfpsort. hgc. jp/)和TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)进行亚细胞定位预测和跨膜结构域预测,并用TBtools(v1.0971)[25]进行可视化。
1.4 BnFUL基因家族进化树分析及共线分析
首先在Ensembl Plants 数据库下载白菜和甘蓝基因组信息,按照1.2 的标准分别鉴定白菜和甘蓝物种中的FUL 家族基因成员;其次利用NCBI 数据库(https://www.ncbi.nlm.nih.gov/)的BLAST 比对功能,获取与甘蓝型油菜BnFUL 家族基因相似性较高的其它十字花科植物序列信息;后利用MEGA X 软件进行多序列比对,并采用邻接法(Neighbor-Joining)构建系统进化树,Bootstrap 值为1000[24]。利用TBtools 进行甘蓝型油菜自身共线性及拟南芥与甘蓝型油菜、拟南芥与甘蓝、拟南芥与白菜物种间的共线分析,参数为1e-5。
1.5 FUL基因家族多序列比对
将拟南芥AtFUL、白菜和甘蓝的FUL基因(各3个)、甘蓝型油菜的BnFUL基因(5个)及共线分析定位在ChrA09 上的基因位点(BnaA09T0074000ZS)进行多序列比对,并提交Pfam分析序列结构域特性。
1.6 BnFUL基因家族表达分析
甘蓝型油菜BnFUL 基因家族在各个部位的基因表达量,由甘蓝型油菜转录组数据库(http://yanglab. hzau. edu. cn/BnTIR)获得;不同品种甘蓝型油菜BnFUL 基因家族各个时期的基因表达量,由甘蓝型油菜泛基因组数据库获得(http://cbi. hzau. edu.cn/bnapus/index.php)。
2 结果与分析
2.1 甘蓝型油菜BnFUL 基因家族鉴定和理化特性分析
通过BLAST 方法,并结合结构域筛选,在甘蓝型油菜中共鉴定到5 个FUL基因,分别命名为Bn-FUL1(BnaA02G0391500ZS)、BnFUL2(BnaA03G040 4700ZS)、BnFUL3(BnaC02G0523700ZS)、BnFUL4(BnaC07G0377400ZS)和BnFUL5(BnaC09G0064500 ZS)。5 个BnFUL基因编码的氨基酸数目为240~242,分子量为27 365.17~27 718.66 Da,理论等电点9.34~9.46,5个BnFUL基因所编码蛋白之间理化性质差异极小,说明该家族在其扩张过程中十分保守,极其稳定。进一步分析发现,BnFUL 蛋白不稳定系数为45.89~53.87,脂溶性指数为83.29~88.63,亲水性平均系数在-0.620~-0.718(表1);亚细胞定位预测结果显示成员均定位在细胞核上。二级结构预测结果显示:BnFUL蛋白二级结构中α-螺旋比例最高,达到51.87%~58.09%;且蛋白家族的二级结构组成较为一致,均为α-螺旋>无规则卷曲>延伸链>β-转角(表2)。
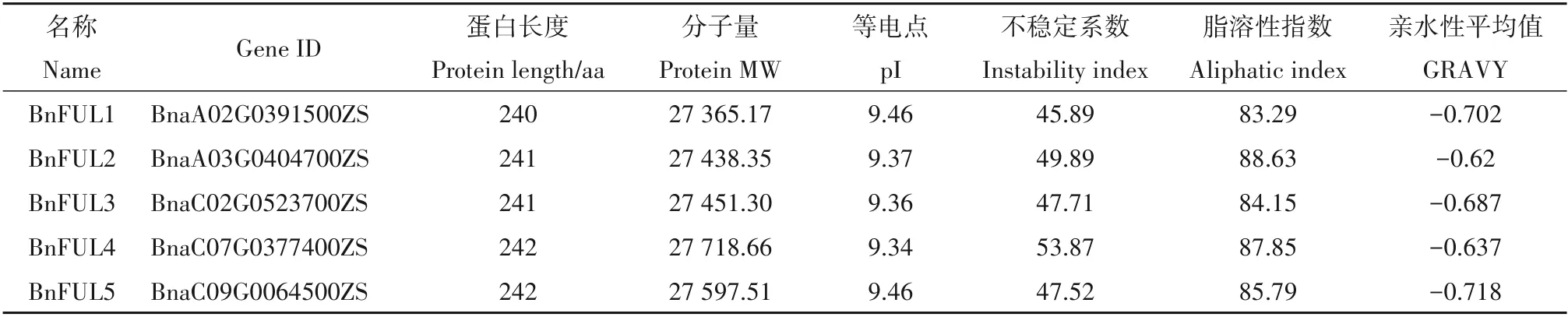
表1 BnFUL蛋白理化特性Table 1 Protein physical and chemical properties of BnFUL proteins
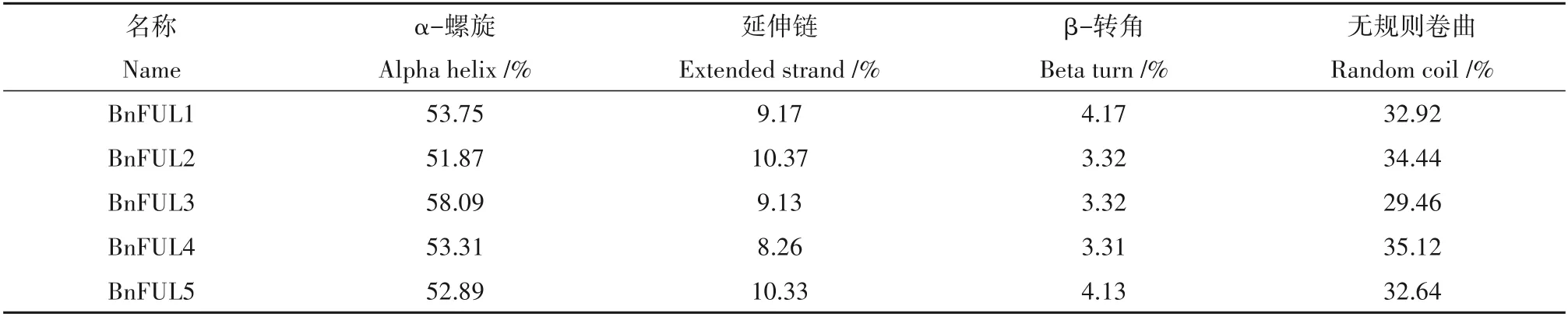
表2 BnFUL蛋白二级结构组成Table 2 Secondary structure composition of BnFUL proteins
2.2 基因结构与蛋白保守基序及结构域分析
使用MEME 对甘蓝型油菜BnFUL 蛋白进行保守Motif 预测,每个蛋白均含有7 个Motif,且排列方式与位置一致。利用Pfam 和NCBI 数据库对甘蓝型油菜5 条FUL 蛋白进行结构域分析,所有成员均含有一个K-box 结构域和一个MADS_MEF2_like 结构域,排列顺序和相对位置无明显差异。对其基因结构分析发现,5 个成员中均没有UTR(未翻译区),且均含有8 个外显子,仅呈现出外显子位置的差异(图1)。

图1 甘蓝型油菜BnFUL基因保守基序、结构域及基因结构Fig.1 Analysis of domain,motif and gene structure of BnFUL genes in Brassica napus
2.3 启动子分析
提取甘蓝型油菜BnFUL 蛋白家族的上游3 kb启动子序列进行顺式作用元件分析(图2),结果显示甘蓝型油菜BnFUL 蛋白启动子区域存在众多顺式作用元件,主要分为3大类:一是与光响应相关的元件,例如:BOX 4、G-box、MRE、I-box、ACE、GATA-motif和LAMP-element等;二是与生物胁迫或非生物胁迫相关元件,例如:ARE、LTR、MBS 和TCrich repeats 等;三是与激素响应相关的元件,例:Pbox、TGA-box、TGACG-motif、CGTCA-motif、ABRE和AuxRR-core等。
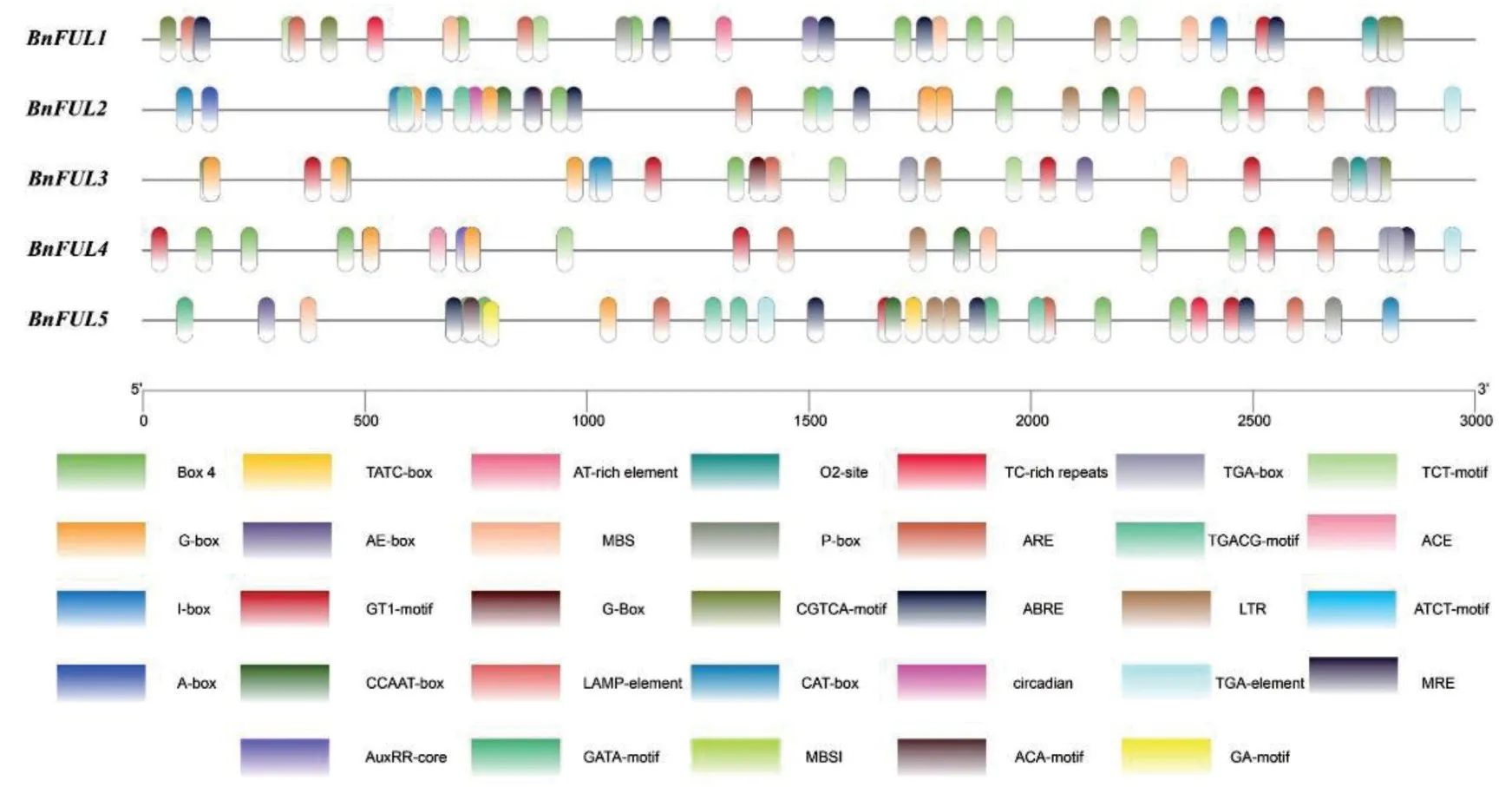
图2 甘蓝型油菜BnFUL基因启动子区域顺式作用元件预测Fig.2 Prediction of cis-acting element of BnFUL genes in B.napus
2.4 染色体定位及共线性分析
为更深入地了解甘蓝型油菜BnFUL基因家族的进化关系,利用TBtools 对甘蓝型油菜自身以及拟南芥与甘蓝、拟南芥与白菜、拟南芥与甘蓝型油菜基因组之间进行共线性分析(图3、4)。5 个BnFUL基因依次定位在染色体ChrA02、ChrA03、ChrC02、ChrC07 和ChrC09 上。共线结果显示白菜和甘蓝两物种均有3 个FUL基因,可能与白菜和甘蓝经历了全基因组三倍化事件有关。拟南芥与甘蓝型油菜的共线分析及自身共线分析显示,甘蓝型油菜有6个BnFUL基因位点,与前文仅鉴定到5 个BnFUL 基因家族成员的结论不符,提示其可能在白菜和甘蓝杂交及多倍化进程中A 基因组上的1 个拷贝丢失(BnaA09T0074000ZS)。
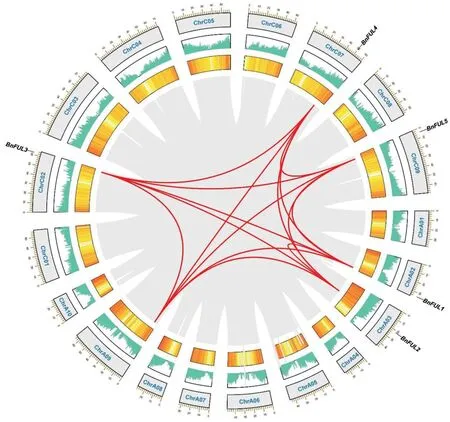
图3 甘蓝型油菜ZS11种内共线性分析Fig.3 Analysis of collinearity in B.napus cv ZS11
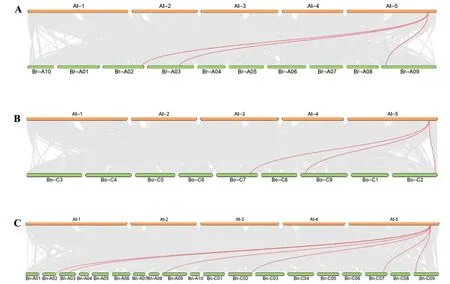
图4 拟南芥与白菜、甘蓝及甘蓝型油菜ZS11之间FUL基因家族共线性分析Fig.4 Collinearity analysis of FUL gene family in Arabidopsis thaliana with B.rapa,B.oleracea and B.napus(ZS11)
2.5 FUL基因家族多序列比对
为明确甘蓝型油菜ZS11 仅鉴定到FUL 基因家族的5个拷贝,而在A基因组上存在1个拷贝丢失的原因,我们对拟南芥、白菜、甘蓝及甘蓝型油菜ZS11的FUL 基因家族与BnaA09T0074000ZS 进行多序列比对(图5)。结果显示,潜在缺失基因(BnaA09T0074000ZS)编码蛋白序列长度(403 aa)明显大于白菜、甘蓝和甘蓝型油菜的FUL 蛋白序列(240~243 aa);进一步经过Pfam鉴定发现,该基因序列仅包含K-box 结构域,丢失了MADS_MEF2_like结构域,且有COBRA 结构域等其它序列的插入与融合,不符合FUL基因的鉴定标准,因此,该基因未被判定为BnFUL基因。
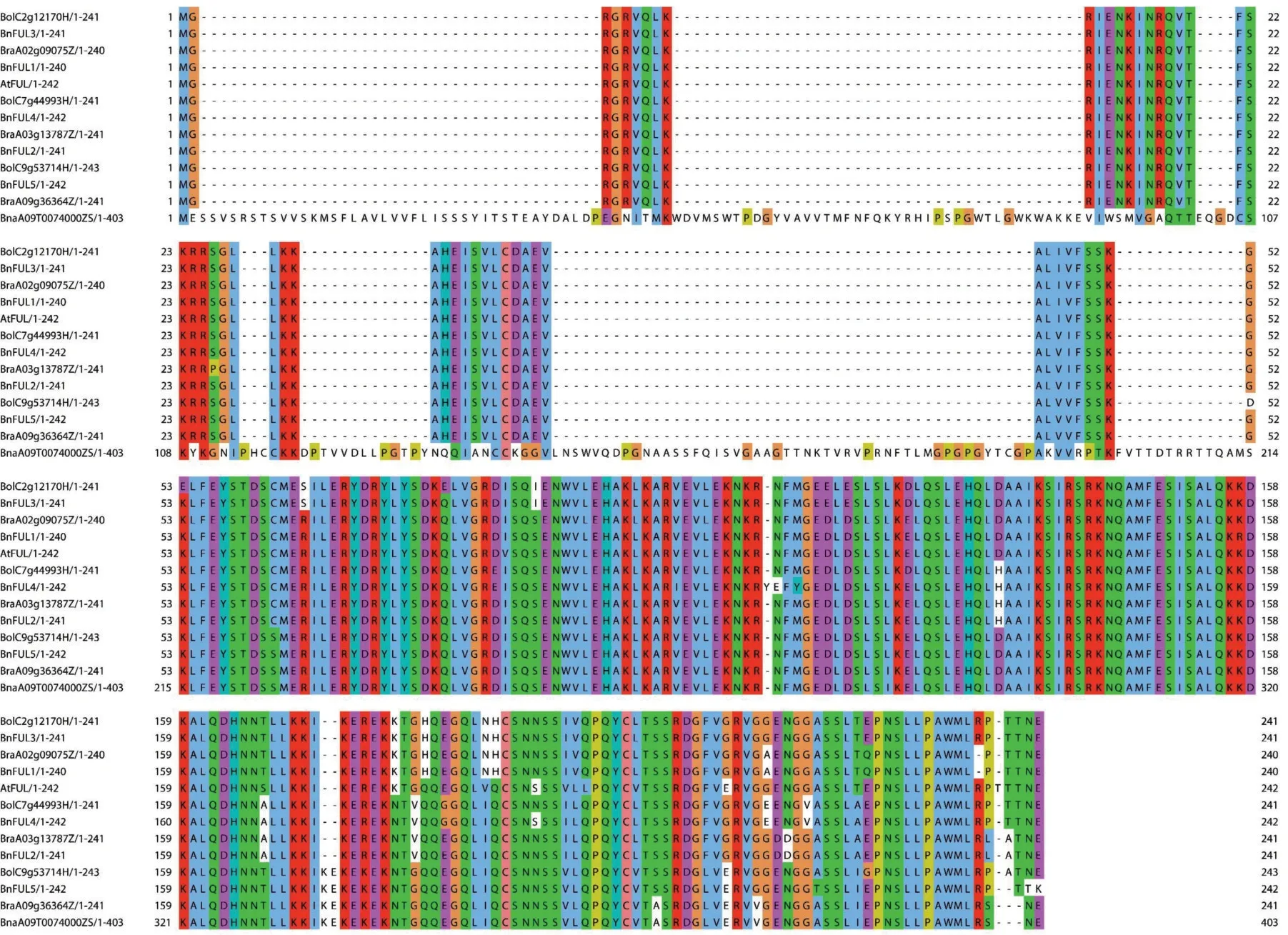
图5 拟南芥、白菜、甘蓝及甘蓝型油菜ZS11的FUL基因家族与BnaA09T0074000ZS的多序列比对Fig.5 Multiple sequence alignment of FUL gene family in A.thaliana,B.rapa,B.oleracea,B.napus(ZS11)with BnaA09T0074000ZS
2.6 系统进化分析
为进一步分析甘蓝型油菜中FUL 家族基因蛋白的进化关系,在NCBI 数据库中搜索并下载与At-FUL基因同源的其它十字花科植物氨基酸序列,并结合白菜和甘蓝全基因组鉴定到的FUL基因,利用MEGA X 软件,将5 个BnFUL与AtFUL及其它11 个FUL基因,分别为野生甘蓝(B. oleraceavar.oleracea,XP_013618458.1)、花椰菜(Brassica oleraceavar.botrytis,CAD47849.1)、甘蓝(B. oleracea)3 个、白菜(B. rapa)3 个、亚麻芥(Camelina sativa,XP_010456827.1) 、萝 卜(Raphanus sativus,XP_018454622.1)和埃塞俄比亚芥(B. carinata,KAG2316775.1)的氨基酸序列进行多序列比对及系统进化树分析(图6)。结果表明:FUL基因在进化过程中十分保守,除白菜BraA09g36363Z在甘蓝型油菜ChrA09 上无对应直系同源基因,其余2 个白菜FUL基因(BraA02g09072Z和BraA03g13786Z)及甘蓝3 个FUL基因(BolC2g12170H、BolC7g44992H和BolC9g53713H)在甘蓝型油菜中均有对应直系同源基因;这与甘蓝型油菜是由白菜和甘蓝杂交及染色体加倍而来的遗传背景一致。另外野生甘蓝(XP_013618458.1)和花椰菜(CAD47849.1)较甘蓝(BolC2g12170H)与甘蓝型油菜BnFUL3有更近的亲缘关系。与拟南芥AtFUL亲缘关系最近的是亚麻芥(XP_010456827.1)。
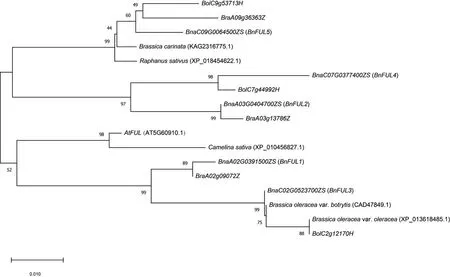
图6 十字花科植物FUL基因系统发育树Fig.6 Phylogenetic analysis of FUL gene in Brassicaceae
2.7 基因表达模式分析
对8 种甘蓝型油菜(ZS11、Quinta、Tapidor、Shengli、Zheyou、Gangan、Westar 和No2127)发育的T0(播种后24 d,温度>10℃)、T1(播种后54 d,温度<10℃)、T2(播种后82 d,温度<10℃)、T3(播种后115 d,温度<10℃)和T4(播种后147 d,温度<10℃)五个不同时期的叶片进行了表达量分析(图7)。结果发现,在8 种甘蓝型油菜的5 个发育时期中,T0 时期表达量最低,大部分未见表达;T2、T3 和T4 时期FUL基因表达量显著上升,这与其在果实发育,角果形成中有重要作用相一致;其中BnFUL3表达量最高,且表达时间跨度最大;而BnFUL4在整个发育时期表达量偏低,尤其在T0 和T1 时期,众多油菜品种该基因均未表达。另外,不同类型甘蓝型油菜在同一发育时期中的表达量有所差别,不同类型甘蓝型油菜的FUL基因家族表达模式不同。
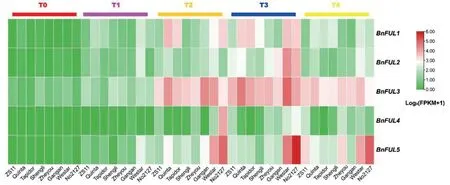
图7 BnFUL基因家族在8个甘蓝型油菜品种5个不同发育时期表达量Fig.7 Expression of BnFUL gene family in 5 different developmental stages of 8 B.napus species
为了解BnFUL 基因家族在甘蓝型油菜不同组织中的表达特征,以甘蓝型油菜品种ZS11 为例,分析了BnFUL 家族基因在甘蓝型油菜叶、茎、根以及角果等组织器官中的表达量,绘制表达模式热图(图8),结果表明BnFUL基因在根、花粉和种子生殖生长时期中表达量低或不表达,在叶片、角果、角果壳和茎中均高表达。
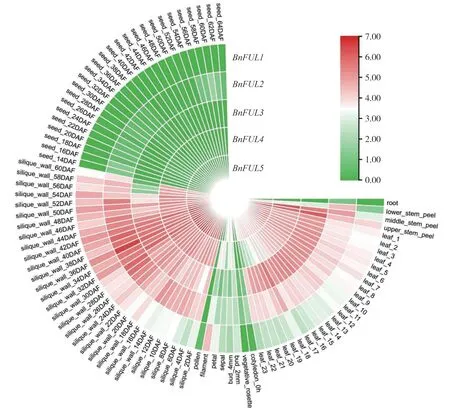
图8 甘蓝型油菜FUL基因家族各个发育时期表达量Fig.8 Expression of FUL gene family in B.napus at different developmental stages
油菜BnFUL 基因家族在不同品种、不同时期及不同部位的表达情况分析表明,BnFUL基因表达存在明显的组织特异性,且在油菜的生长发育与果实成熟过程中发挥重要作用。
3 讨论
本研究利用生物信息学的方法在甘蓝型油菜中共鉴定到5 个BnFUL基因,都属于MADS-box 转录因子家族,MADS-box 转录因子家族一般存在两个结构域:K-box 主要参与蛋白的二聚体或多聚体的特异结合,MADS-box 能与DNA 结合并调控下游基因表达[15]。对其基因结构、所编码蛋白的基序和结构域等方面进行分析,发现该家族成员在进化过程中比较保守,成员之间理化特性、蛋白结构域等均无明显差异,仅有外显子的排列位置差异较大。对其启动子区域顺式作用元件的分析显示,BnFUL基因上游启动子区域存在大量与光响应有关的元件,与其具有调控花发育的功能相一致,另有与生物胁迫、非生物胁迫和激素响应相关的元件,推测BnFUL基因可能在油菜的生长发育过程中起重要作用[2,3]。
芸薹属的白菜和甘蓝在7500 年前经全基因组三倍化后,进而杂交形成甘蓝型油菜。白菜基因组和甘蓝基因组中均鉴定到3 个FUL基因位点,理论上甘蓝型油菜FUL基因应为6 个拷贝,且共线性分析显示存在6 个共线性位点,与本文仅鉴定到5 个FUL基因结论不一致,提示其在A 基因组(ChrA09)上有该基因拷贝数的缺失。进一步分析该基因位点的缺失原因,我们将不一致的基因位点(BnaA09T0074000ZS)序列进行分析,发现该基因仅有K-box 结构域,但无MADS_MEF2_like 结构域,所编码的氨基酸长度明显大于其它成员,有COBRA 结构域等其它序列的插入与融合,表明在甘蓝型油菜ChrA09 染色体上的部分基因片段可能在染色体加倍过程中发生了基因的断裂和融合,导致部分基因的缺失。这与Sheng 等认为植物在基因组发生复制之后,往往出现大面积的基因丢失和染色体重排,用来维持代谢平衡或其它生长发育需要的观点一致[26]。
植物中FUL基因不同的表达模式与其功能有明显的相关性。例如,禾本科植物小麦(Triticum aestivum)的3 个FUL基因,WFUL1在叶片中表达,WFUL2调控外稃和外稃发育,而WFUL3则参与调控果实发育[27]。豆科植物苜蓿(Medicago truncatula)中的3 个MtFUL基因:MtFULa和MtFULb在各阶段均有表达,开花前表达量最高,主要促进开花;而MtFULc主要在开花后表达,促进花的发育;同时在拟南芥中异源过表达MtFUL造成拟南芥开花提前[28]。本次结合公开数据对BnFUL基因表达谱进行分析,8个甘蓝型油菜品种在5个不同时期的分析显示:T3 时期(播种后115 d)各个基因的表达量均高于其它时期,BnFUL3更为明显,且在不同品种间均高表达;相比之下,BnFUL4表达量较其它时期没有显著变化。进一步结合油菜品种ZS11 在各个生长阶段的表达量分析:BnFUL在叶、茎、花、长角果发育初期和角果壳中持续高表达,而在果实、花粉和根中基本不表达。BnFUL 蛋白保守基序基本相似,只有在“C”末端氨基酸区域相似度不高,结合表达谱表达模式的不同,推测BnFUL4可能与其它Bn-FUL基因间在功能上存在差异。
彭鹏飞等在拟南芥中过表达BnFUL能提高拟南芥角果的抗裂性,同时促进早开花,甚至表现出一个果柄多角果的现象[29]。因此,FUL基因可作为筛选和开发抗裂种质资源的重要候选基因,后期可利用CRISPR-Cas9 技术和基因工程等方法促进Bn-FUL在甘蓝型油菜自身中表达,来获得优质油菜新种。本文基于甘蓝型油菜基因组序列,从全基因组角度系统鉴定并对甘蓝型油菜的BnFUL 基因家族成员进行基本理化特性、进化发育关系和表达模式分析,利于阐释甘蓝型油菜BnFUL 基因家族的功能,为油菜的分子育种尤其是甘蓝型油菜抗裂基因的挖掘和抗裂角油菜新品种的培育提供依据。
