sRNA SaaS对沙门氏菌粘附响应规律和胞外代谢物的影响效应
2022-03-11何淑雯胡海静陆许秋王浩东王虎虎徐幸莲
何淑雯 胡海静 陆许秋 王浩东 王虎虎 徐幸莲
(南京农业大学国家肉品质量安全控制工程技术研究中心/江苏省肉类生产与加工质量 安全控制协同创新中心,江苏 南京 210095)
沙门氏菌(Salmonellaspp.)是一种重要的食源性致病菌,在全球范围内每年可造成大量的食品安全事件并造成食品行业巨大的经济损失[1-2]。在食品加工过程中的接触表面形成生物菌膜是沙门氏菌实现传播和污染的重要途径。生物菌膜是细菌粘附于接触表面后,通过增殖和分泌物胞外多聚物而形成的具有一定空间结构的生物群体[3]。越来越多的证据表明,控制生物菌膜形成过程的分子网络受到非编码sRNA的调控。非编码sRNA是一类在基因组中转录但不编码蛋白质的RNA。目前,生物菌膜的形成在转录后水平的调控机制已成为食品安全的研究热点,但现有调控通路仍不能完全解释肠炎沙门氏菌的粘附过程。sRNA SaaS (Salmonellaadhesion associated sRNA) 是近期在肉品源肠炎沙门氏菌中筛选出的一种新型调控因子,结晶紫染色试验结果显示该sRNA能够显著抑制生物菌膜的形成,但其具体功能和调控路径尚不清楚[4]。
生物菌膜的形成离不开物理接触表面的存在,在食品生产和日常生活中,机械加工设备和不同材质食品盛装器具的表面均可成为细菌粘附的载体。了解细菌在接触表面的粘附响应规律对于理解粘附因子的具体功能具有指导意义。近年来,相关研究或多以不锈钢作为接触面材质加以展开[5-6],或对环境温度的设置较为单一[7-8],而同时使用多种材质的接触面并在多个温度下进行观测的研究相对较少。
胞外代谢物是生物菌膜的重要组成部分,解析胞外代谢物有助于更直观有效地了解生物菌膜的形成过程及其机理。代谢组学通过对所有代谢物进行分析,提供代谢途径中发生变化的关键信息,从而揭示生物体生命活动发生和发展的本质[9]。广泛靶向代谢组学(widely-targeted metabolomics)结合了靶向代谢组学和非靶向代谢组学两种传统分析方法的优点,能够利用基于多反应监测模式(multiple reaction monitoring,MRM)的四极杆-线性离子阱串联质谱法(QTRAP),对代谢物实现准确的鉴定和检测[10]。近年来,代谢组学已广泛运用到微生物领域[11-13]。
针对上述现象和问题,本研究以分离自肉品加工接触面的肠炎沙门氏菌为试验对象,使用前期成功构建的sRNA SaaS基因缺失突变株为试验组,野生株为对照组,对浮游态和粘附态两种生存形式的菌体展开试验;通过研究细菌的粘附响应规律初步判定SaaS的作用条件;进一步利用广泛靶向代谢组学方法对胞外代谢物进行分析,以期揭示SaaS对生物菌膜形成的分子机制提供线索,完善生物菌膜的理论体系,为研发生物菌膜新型控制技术提供新思路。
1 材料与方法
1.1 材料与试剂
胰蛋白胨大豆琼脂(tryptic soy agar, TSA)、胰蛋白胨大豆肉汤(trypticase/tryptic soy broth,TSB)、木糖赖氨酸脱氧胆盐琼脂(xylose lysine deoxycholate agar,XLD),青岛海博生物技术有限公司;氯化钠,国药集团上海化学试剂有限公司;甲醇、乙腈、乙醇,德国Merck公司。
本试验所用肠炎沙门氏菌(S.Entertidis NCM 61,Accession:PRJNA492709)分离自冰鲜鸡肉屠宰分割台接触面,sRNA SaaS缺失突变株由λ-Red 同源重组技术构建得到[3]。
1.2 仪器与设备
BIO Ⅱ Advance 4生物安全柜,美国BAKER公司;SQL 1010C高压灭菌锅,重庆雅马拓科技有限公司;SPX-250B-Z型生化培养箱,上海博讯实业有限医疗设备厂;QTRAP 6500+质谱仪,美国AB SCIEX公司;UPLC 30A色谱仪,日本SHIMADZU公司;5427 R台式高速冷冻离心机离心机,德国艾本德Eppendorf股份公司;easySpiral Pro全自动螺旋接种仪,法国Interscience有限公司。
1.3 试验方法
1.3.1 菌液准备 取冻存于-80℃的肠炎沙门氏菌野生株和突变株,室温溶解后,在XLD平板上划线,于37℃培养24 h。挑取单菌落于TSB培养基,于37℃培养24 h后,吸取菌悬液并接种至新鲜的TSB培养基,于37℃培养18 h,制成初始浓度为109CFU·mL-1的菌悬液,作为工作菌液。
1.3.2 粘附响应规律 将标准尺寸(75 mm×25 mm×1 mm)的不锈钢、玻璃、聚丙烯3种材质的接触表面薄片(以下简称“薄片”)均匀地插入塑料支架,使薄片直立地分布于可密封玻璃容器中,高压灭菌(121℃、15 min)后备用。使用TSB培养基将工作菌液稀释至102CFU·mL-1,随后注入上述玻璃容器中,菌悬液的用量能够使薄片一半浸没于液体,一半暴露于空气中。密封后,于15、20、37℃分别培养24、48、72、96、120 h,培养过程中避免液面晃动。培养至规定时间后,使用0.85%生理盐水洗去薄片表面的浮游菌体,控干液体后将薄片投入装有40 mL 生理盐水的均质袋中,使用拍击式均质器以每分钟360下的速度连续拍击1 min。接着使用生理盐水对均质液进行10倍梯度稀释,并选取2~3个稀释度,用全自动螺旋接种仪吸取100 μL稀释液并均匀地涂布至TSA平板,于37℃培养20 h后计数。每组试验重复4次。
1.3.3 胞外代谢组学分析
1.3.3.1 胞外多聚物的采集 使用TSB培养基将工作菌液稀释至102CFU·mL-1,并接种于48孔细胞板,每块孔板接入同一株菌,每孔接入800 μL稀释液。同时,接种无菌TSB作为阴性对照。将细胞板置于37℃培养48 h。轻轻倒出48孔板中的培养基,用0.85%生理盐水清洗3次后吸除每孔中残留的液体。向任意一孔加入200 μL生理盐水,使用无菌棉签蘸取液体后,依次对每一孔壁进行擦拭,将擦得的粘附体收集到2 mL无菌离心管中。再次向任意一孔加入400 μL生理盐水,用移液枪反复吹打收集残留的菌膜,随后将液体转移至下一微孔,重复上述操作,直至每孔中的菌膜被完全收集,将孔板和棉签上的菌液全部转移至上述2 mL离心管。离心(5 000×g,10 min,4℃)后,上清液即为胞外多聚物溶液(extracellular polymeric substances,EPS)[14]。每组试验重复6次。
1.3.3.2 液相色谱-质谱联用分析 取50 μL的EPS,向其中加入150 μL预冷的纯甲醇提取剂,涡旋1 min后,放入液氮中速冻3 min,取出,冰上解冻3 min,涡旋2 min,循环3次。离心(12 000 r·min-1,10 min,4℃)后,取上清液用于液相色谱-串联质谱联用(liquid chromatography-tandem mass spectrometry, LC-MS-MS)分析。
液相条件:色谱柱:Waters ACQUITY UPLC HSS T3 C18 1.8 μm,2.1 mm ×100 mm; 流动相:A相为超纯水(0.04%乙酸),B相为乙腈(0.04%乙酸);洗脱梯度: 0 min 水/乙腈(95∶5,v/v),11.0 min 为 5∶95 v/v,12.0 min为 5∶ 95 v/v,12.1 min 为 95∶5 v/v,14.0 min 为 95∶5 v/v;流速 0.4 mL·min-1;柱温 40℃;进样量 2 μL。
质谱条件:电喷雾离子源(electrospray ionization, ESI)温度500℃,质谱电压 5 500 V(positive),-4 500 V(negative),离子源气体Ⅰ(ion source gas Ⅰ, GS Ⅰ)55 psi,气体Ⅱ(Gas Ⅱ, GS Ⅱ)60 psi,气帘气(curtain gas, CUR)25 psi,碰撞诱导电离(collision-activated dissociation, CAD)参数设置为高。在三重四极杆中,每个离子对根据优化的去簇电压(declustering potential, DP)和碰撞能(collision energy, CE)进行扫描检测。
1.3.3.3 代谢物定性和定量 基于靶向标品数据库(metware database,MWDB)对信息进行定性分析。代谢物定量利用三重四极杆质谱的多反应监测模式(MRM)分析完成。差异代谢物的筛选标准为:变异系数(fold change,FC)≥ 2 和 FC ≤ 0.5;变量重要性投影(variable importance in the projection,VIP)≥ 1。
1.3.4 数据处理 使用SPSS 23.0对数据进行统计分析,采用独立样本t检验进行显著性分析,以P<0.05表示差异显著,P<0.01表示差异极显著。使用Prism 8.0软件绘图。
2 结果与分析
2.1 粘附响应规律
由图1可知,在本研究中,3种材质的接触表面在37℃下沙门氏菌菌体粘附数量最高,20℃次之,而15℃最低。由曲线的走势可以发现,肠炎沙门氏菌的粘附数量随着时间的推移总体呈现出先增长后趋于平稳并有缓慢下降的趋势,这与生物菌膜形成过程中所经历的粘附、成熟、瓦解等关键阶段吻合,生物菌膜生长规律在其中有所体现。值得关注的是,在37℃下,ΔSaaS的菌膜形成能力显著大于WT,该现象在3种接触面上均有发生,并在聚丙烯表面尤为明显;而在20和15℃下,除聚丙烯表面外并未发现类似现象。

注:A:不锈钢表面;B:玻璃表面;C:聚丙烯表面。内嵌图为37℃下粘附曲线的局部放大图。 **代表WT与ΔSaaS在0.01水平有显著性差异。Note:A: Stainless steel surface. B: Glass surface. C: Polypropylene surface. The inline images are a zoom of the adhesion curve at 37℃. ** indicates significant differences at 0.01 level between WT and ΔSaaS.图1 粘附响应规律Fig.1 Adhesion law in response to environmental conditions
2.2 胞外代谢组学
2.2.1 代谢物定量结果 肠炎沙门氏菌生物菌膜胞外代谢物的定量结果如表1所示。本研究检测到ΔSaaS和WT的代谢物总数为275;差异代谢物总数为54,其中53个显著上调。
2.2.2 主要差异代谢物 本研究根据差异倍数大小,对检测到的差异代谢物进行了排名,并将排名前20的代谢物展示如下(图2)。在显著上调的差异代谢物中,琥珀酸和氨基丙二酸的差异倍数最大;二乙醇胺为唯一显著下调的差异代谢物。
2.2.3 差异代谢物KEGG功能注释及富集分析
2.2.3.1 差异代谢物KEGG分类 本研究对差异显著代谢物的KEGG注释结果按照通路类型进行了分类,分类结果如图3所示,纵坐标为KEGG代谢通路的名称,横坐标为注释到该通路的代谢物个数及其个数占被注释的代谢物总数的比例。本试验差异代谢物的注释结果主要集中于代谢途径(92.5%)、ABC转运蛋白(35%)、二级代谢物的生物合成(25%)、不同环境中的微生物代谢(20%)、嘧啶代谢(18%)、嘌呤代谢(15%)、氨基酸的生物合成(15%)和精氨酸和脯氨酸代谢(15%)等通路。

表1 差异代谢物数量统计Table 1 Statistics of differential metabolites

图2 差异代谢物条形图Fig.2 Bar graph of differential metabolites
2.2.3.2 差异代谢物KEGG富集 本研究对差异代谢物进行了KEGG通路富集(图4)。本研究中的差异代谢物较为显著地富集于代谢途径、ABC转运蛋白、嘧啶代谢、嘌呤代谢等通路。
3 讨论
接触表面材质可能因为粗糙度、表面疏水性等特性的不同而影响生物菌膜的形成[15-17]。本研究以304不锈钢、玻璃和聚丙烯塑料作为载体,并于不同环境温度下对生物菌膜进行培养,其中37℃为沙门氏菌生长的最适温度,20℃贴近大部分肉类加工车间的环境温度,15℃用于模拟低温。本研究发现,肠炎沙门氏菌在不同材质接触表面的粘附量并无明显差异。而De Oliveira等[18]研究禽肉源沙门氏菌时发现,菌膜在玻璃、聚氯乙烯和不锈钢上的长势不同,且玻璃最不利于其形成。该结果与本研究结果不一致,可能是由于菌株的来源或种类不同。值得注意的是,在37℃下,ΔSaaS与WT的粘附差异在初始粘附和微菌落形成阶段(24 h)相对较小,在菌膜成熟阶段(72和96 h)明显扩大。说明sRNA SaaS能够在37℃下抑制生物菌膜的形成,且这种调控作用可能最早发生于粘附期,并在成熟期得到强化。基于以上发现,本研究进一步对37℃下野生株与突变株生物菌膜的胞外代谢物进行了分析。

图3 差异代谢物KEGG分类图Fig.3 KEGGClassification of differential metabolites

图4 差异代谢物KEGG pathway富集结果Fig.4 Enrichment of KEGG pathways of differential metabolites
差异代谢物大量富集于ABC转运蛋白途径,ABC转运蛋白属于膜结合蛋白,对生物菌膜的形成起到重要作用[19-20]。Vanderlinde等[20]发现,ABC转运蛋白的缺失会造成胞外多糖分泌减少,并使生物菌膜的形成受到严重损害。在本研究中,该途经下的无机与有机离子转运系统中的亚精胺、腐胺、甘氨酸甜菜碱等代谢物在突变株组发生了上调。sRNASaaS的缺失可能通过促进亚精胺/腐胺结合周质蛋白(spermidine/putrescine-binding periplasmic protein, PotD) 的合成,从而改变多胺的合成速率,并刺激生物菌膜的形成[21];与此同时,甘氨酸甜菜碱也具有恢复和促进生物菌膜形成的作用[22-23]。
嘌呤/嘧啶代谢途径也富集了大量差异代谢物,腺嘌呤、尿嘧啶、肌苷等代谢物在突变株组发生上调。研究表明,核苷酸生物合成途径与卷曲菌毛和纤维素的产生紧密相关,且嘧啶核苷酸的可用性能够强烈影响卷曲蛋白操纵子的转录[24]。卷曲菌毛和纤维素是生物菌膜胞外多聚物的重要组分。前者与细菌在非生物表面的粘附和菌膜形成相关[25],后者有助于成熟菌膜三维结构的形成并对菌体细胞起保护作用[26]。而嘌呤生物合成途径能刺激多糖的合成,外源尿嘧啶能触发纤维素的合成,外源添加肌苷能使生物菌膜的形成量显著增加[27-28]。sRNA SaaS的缺失可能通过刺激核苷酸的合成,加强EPS的分泌,从而促进细胞间的粘附和生物菌膜的成熟。另一方面,嘌呤/嘧啶代谢涉及ATP、DNA与RNA的合成。胞外DNA(eDNA)作为胞外多聚物,起到维持菌膜三维立体结构、为细胞提供底物和增强遗传物质交流的作用[29]。sRNA SaaS的缺失也可能通过促进核甘酸的合成代谢,增加eDNA的合成与分泌,进而促进成熟菌膜的形成。
氨基酸代谢途径也有较多差异代谢物富集,且N-乙酰天冬氨酸、S-甲基-L-半胱氨酸、丙氨酸、谷氨酸等氨基酸在突变组发生上调。Rajendran等[30]在针对白色念珠菌(Candidaalbicans)的研究中也观察到类似现象,发现氨基酸代谢在生物菌膜形成能力较强的分离株组中上调。氨基酸代谢是细菌正常生长的重要条件,在生物菌膜形成早期,氨基酸水平的增加有助于菌膜成熟阶段细菌生物量的增加[31]。此外,与能量代谢相关的琥珀酸[32]在本试验突变组也发生大幅上调,为变化最显著的代谢物。这说明sRNA SaaS缺失突变株可能具有更强的碳代谢率,能够更快地进行增殖,进而能够在生物量的维度上更快地形成生物菌膜。
综上,sRNA SaaS可能通过抑制氨基酸代谢和碳代谢减缓细菌的增殖,同时通过抑制嘌呤/嘧啶代谢,减少卷曲菌毛、纤维素、多糖等胞外多聚物的合成与分泌,分别从菌体细胞的数量和胞外多聚物的产量两方面抑制肠炎沙门氏菌生物菌膜的形成,并阻碍菌膜三维立体结构的维持(图5)。
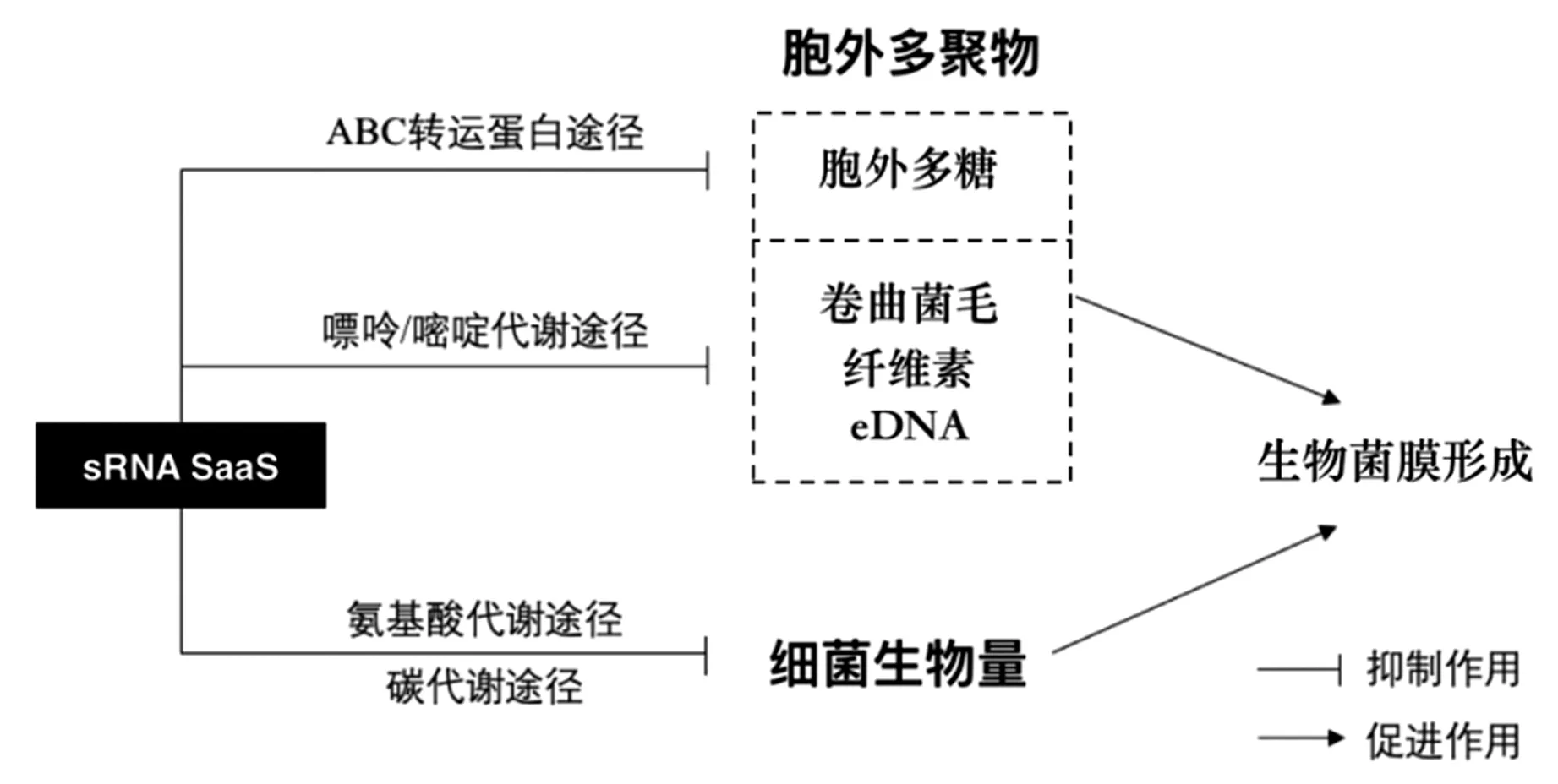
图5 sRNA SaaS对生物菌膜形成的作用机制Fig.5 Mechanism of sRNA SaaS on biofilm formation
4 结论
本研究发现sRNA SaaS对肠炎沙门氏菌粘附能力的影响具有温度依赖性。在37℃下,SaaS 能够抑制该菌在304不锈钢、玻璃和聚丙烯塑料表面形成生物菌膜,且抑制作用在聚丙烯表面尤为明显。进一步分析发现,sRNA SaaS可能一方面通过减缓细菌的增殖,另一方面通过减少卷曲菌毛、纤维素、多糖等胞外多聚物的合成与分泌,实现对生物菌膜形成的抑制作用。本研究成果有助于完善生物菌膜的理论体系,为揭示sRNA SaaS对生物菌膜形成的分子作用机制提供线索,为研发生物菌膜新型控制技术提供新思路。
