芥菜型油菜细胞质雄性不育系WJS01A的比较鉴定
2022-03-09胡文竞吴江生
孙 植,杨 谦,胡文竞,王 芳,吴江生,刘 超
(华中农业大学 国家油菜工程技术研究中心/农业农村部油菜遗传育种重点实验室,武汉 430070)
利用细胞质雄性不育(cytoplasmic male sterility, CMS)生产杂交种是作物杂种优势利用的主要途径之一,可显著提高作物的产量[1-2]。植物CMS一般是由线粒体基因组重排产生异常嵌合基因导致的,嵌合基因编码的多肽进一步影响了花粉的正常发育[3]。油菜(BrassicanapusL.)是中国乃至世界上最重要的油料作物之一,加强油菜杂种优势利用将有利于中国油菜产业的发展,进而保障中国食用油供给安全[4-5]。目前,在油菜中已被发现的细胞质雄性不育类型有10多种,譬如ogu、pol、nap、shan2A、Nca、Nsa、hau和inap等[6]。尽管它们中部分已实现三系配套[7-13],但是仅有pol CMS和ogu CMS在油菜生产上大规模应用[14]。目前中国油菜杂交种配制仍然以pol CMS为主,ogu CMS在欧洲和北美等地的油菜杂交种生产中得到广泛应用[15],但是因专利问题,导致其在中国的应用受到限制。目前中国在油菜杂种优势上已经形成了细胞质单一化的局面,油菜品种的遗传基础越来越狭窄,对油菜的安全生产存在潜在威胁[16]。因此创建新型优良胞质不育类型,并获得其配套的保持系和恢复系,对丰富油菜遗传基础,选育高产优质油菜新品种具有重要的现实意义。
WJS01A是利用本课题组在芥菜型油菜(Brassicajuncea)农家品种‘玉溪高棵’中发现的天然雄性不育株育成的细胞质雄性不育系[17]。进一步通过回交转育获得了甘蓝型油菜遗传背景的不育系WNJ01A,同时在WNJ01A与白菜(Brassicarapa)远缘杂交后代中选育获得了恢复系‘Hui01’,该恢复系可以完全恢复WNJ01A的育性。本研究对WJS01A进行细胞学、形态学、遗传学和分子生物学鉴定,研究结果将有助于揭示其败育机制,为该不育系在油菜育种中的应用提供理论基础。
1 材料和方法
1.1 试验材料
本研究所用实验材料均列于表1,其中NRO4270A是我们在萝卜甘蓝(Raphanusbrassica, RRCC)与甘蓝型油菜远缘杂交后代中发现的一种CMS类型[18]。实验材料种植于华中农业大学试验田,以及湖北省长阳县、巴东县和甘肃省和政县等油菜夏季繁殖基地。
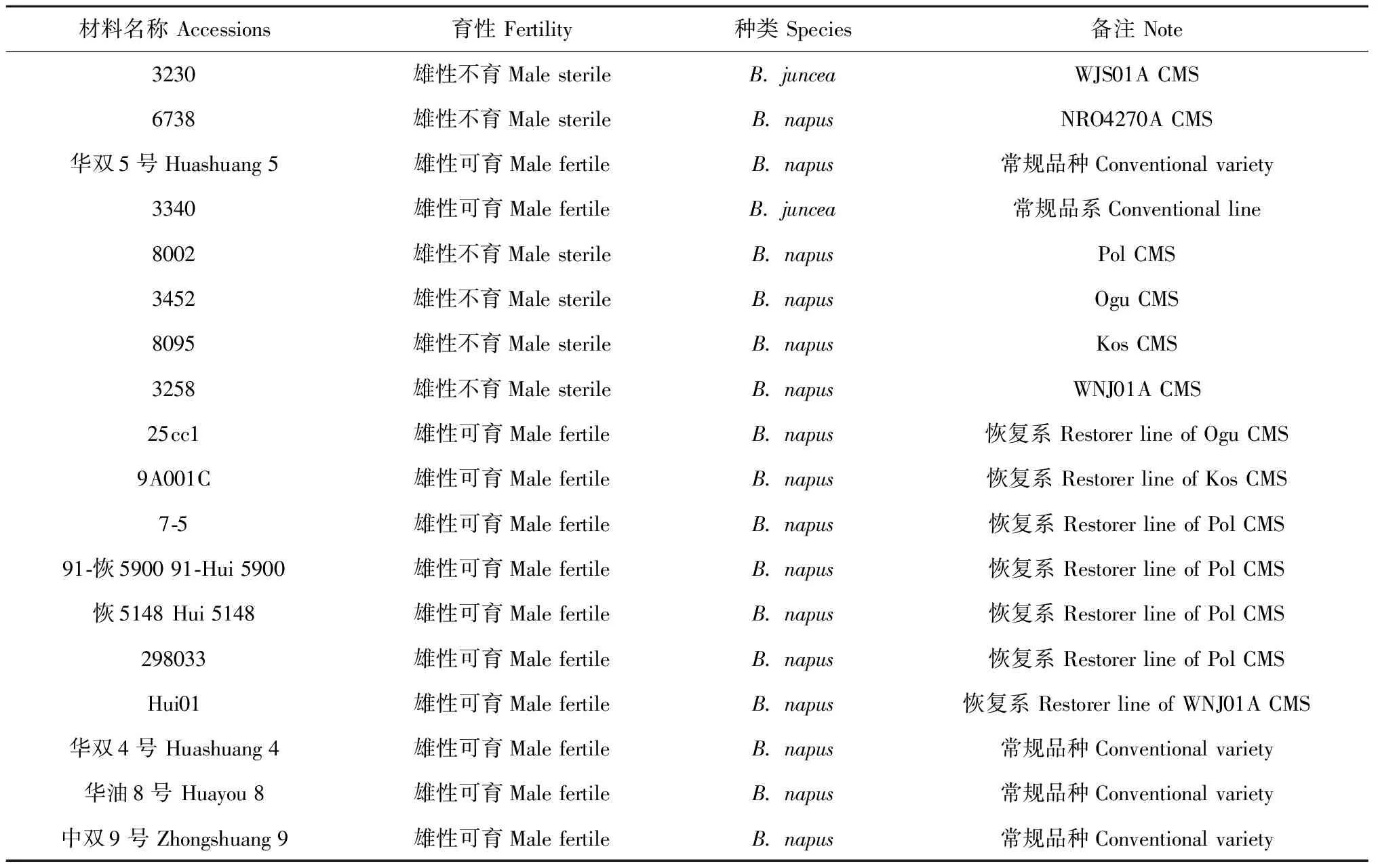
表1 实验用到的材料
1.2 形态观察和育性鉴定
在盛花期对不育系WJS01A及其衍生不育系WNJ01A的花粉育性开展调查,育性调查的分级标准采用杨光圣、傅廷栋[19]制定的方法,并观察其是否存在死蕾现象。
1.3 恢保关系测定
在开花期选取7个恢复系以及3个甘蓝型油菜常规品种,分别与WNJ01A、Pol、Ogu 和Kos进行测交,收获F1杂种并于下一种植季种植于田间。在开花期对每个单株至少5朵花进行不少于3次的育性调查,采用育性分级标准鉴定[19],以保证调查结果的准确性。
1.4 花药石蜡切片
取不育系WJS01A的花蕾制作石蜡切片,具体步骤参照植物显微技术的方法操作[20]:(1)取材和固定:在油菜开花期,取大小不同花蕾浸没于FAA固定液(70%酒精90 mL,福尔马林5 mL,冰醋酸5 mL)中,固定24 h;(2)染色和清洗:将固定后花蕾中的花药剥离,加入爱式苏木精染料进行整染4 d,用蒸馏水漂洗3~5次,每次0.5~1 h;(3)乙醇脱水:按照30%、50%、70%、85%和95%不同浓度乙醇的顺序分别对材料进行逐级脱水处理1 h,再用无水乙醇脱水2次,每次1 h;(4)氯仿透明:按照20%、40%、60%和80%不同浓度氯仿分别对材料进行逐级透明处理1 h,再用氯仿处理2次,每次1 h;(5)浸蜡和包埋:加入碎蜡后在37 ℃恒温箱中温育2 d,随后在金属盒中进行逐级浸蜡,50%石蜡(48 ℃)温育2 h,75%石蜡(53 ℃)温育2 h;纯蜡A(56 ℃),纯蜡B(56 ℃),纯蜡C(56 ℃),每级温育0.5~1 h;(6)切片:将包埋好的蜡块在轮转式切片机(KD-2508)上切片,随后用蛋清甘油粘片和展片,并放置37 ℃烘片3 d以上;(7)脱蜡和封片:二甲苯脱蜡处理0.5~1 h后用加拿大树胶封片,放置37 ℃烘箱烘片3 d以上。利用Nikon Ds-Ri1全自动显微镜对切片进行观察并照相。
1.5 多重PCR鉴定
参考多重PCR区分油菜不同细胞质类型的鉴定方法[21],以WJS01A、NRO4270A、Kos、Ogu和Pol等5种雄性不育材料以及衍生不育系WNJ01A DNA为模板,用等量混合的3对引物进行多重PCR,分别鉴定线粒体基因orf138、orf222/nad5c和orf224[21]。多重PCR采用的反应体系为:50 ng模板DNA,2 μL 10×PCR 缓冲液,1.2 μL MgCl2(25 mmol/L),0.4 μL dNTP(10 mmol/L),0.5 U Taq DNA聚合酶,6 μL混合引物,添加ddH2O至20 μL。PCR反应程序为:94 ℃预变性2 min,94 ℃变性30 s,60 ℃退火50 s,72 ℃延伸70 s,35个循环,72 ℃终延伸7 min。反应结束后,取5 μL PCR产物加5 μL上样缓冲液,用2%的琼脂糖凝胶电泳。100 V电压下电泳35 min左右,用凝胶成像系统观察、照相。
1.6 不同胞质类型的RFLP分析
1.6.1 基因组DNA的提取和酶切取WJS01A、Pol、Ogu、Kos和NRO4270A的新鲜叶片5g,采用CTAB方法[22]提取总DNA。用EcoRI和BamHI(Fermentas)进行总DNA酶切,酶切体系为:15 μg模板DNA,5 μL 10× buffer,3 μL限制性内切酶(10 U/μL),添加ddH2O至50 μL。在37 ℃条件下过夜酶切,然后加入1/10体积的上样缓冲液终止酶切反应。
1.6.2 电泳与转膜将酶切产物用琼脂糖凝胶电泳(电压35 V)20 h 后,转移凝胶至0.25 mol/L HCl中处理10 min,直至溴酚蓝变为黄色(脱嘌呤);蒸馏水漂洗后用0.4 mol/L的NaOH处理10 min,重复1次(30 min)后再用蒸馏水漂洗;然后用0.4 mol/L NaOH转膜缓冲液将凝胶DNA转移到Hybond N+尼龙膜上;最后用2×SSC溶液漂洗尼龙膜20 min,于80 ℃烘箱中烘烤2 h后置于4 ℃备用。
1.6.3 探针的制备与标记根据已知的油菜线粒体全基因组序列[23],利用软件Primer 3.0设计atp1、atp6、atp9和cox1等4对线粒体引物(表2)。通过PCR扩增得到探针后用DNA凝胶回收试剂盒(AxyPrep)回收探针DNA。标记探针采用随机引物法,先将2.0 μL探针添加11.0 μL ddH2O,沸水处理5 min;冰浴5 min后,依次加入2.5 μL 10×buffer,2.5 μL dNTPs(-dCTP),1.0 μL Klenow Fragment,2.0 μL随机引物,2.0 μL[α-32P]dCTP(10Ci/μL),置于37 ℃水浴2~4 h;沸水变性10 min后,置于冰上备用。
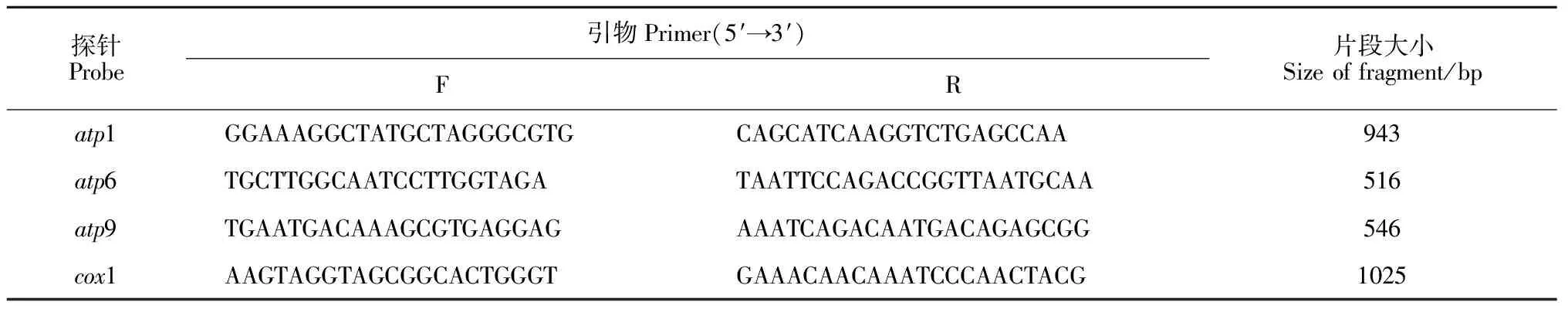
表2 用于RFLP的线粒体探针引物
1.6.4 探针杂交与放射自显影将已标记的探针加入65 ℃预热的300 μL预杂交液,放入杂交炉中,65 ℃条件下与尼龙膜杂交12~16 h;取出尼龙膜,用30 mL洗膜液A(2×SSC、0.1% SDS),洗膜8~10 min,重复2次;再用洗膜液B(0.5×SSC、0.1% SDS、65 ℃预热)洗膜30 min,重复2次。将杂交尼龙膜压在磷屏(Fujifilm BAS-MS 2025)上处理6~8 h,然后用多功能成像系统(Fujifilm FLA-9000)扫描磷屏检测杂交结果。
2 结果与分析
2.1 WJS01A花器官形态观察及育性鉴定
对不育系WJS01A及正常芥菜型油菜3340花序和花器官进行观察发现,它们的花序结构差异不大,然而对于花器官,WJS01A的花蕾相较于3340略显瘦小,且它的花朵张开度、花瓣长度和宽度略小于3340(图1, A、B)。WJS01A的雌蕊发育正常,柱头高于雄蕊,花丝缩短,花药变小、白化且表面无花粉(图1, C)。而3340的柱头稍低于花药,且花丝粗壮,花药饱满,表面布满大量黄色花粉(图1, D)。此外,我们研究发现WJS01A来源的甘蓝型油菜背景不育系WNJ01A花器官的花丝短小,且花药呈现不同程度的白化并缩小为三角状[17]。
为鉴定WJS01A不育表型对环境的敏感性,连续多年多代(2010—2021)对其了进行育性调查。结果表明WJS01A的不育性稳定,在湖北省武汉市、长阳县、巴东县和甘肃省和政县种植的不育系WJS01A育性标准为0级,不受环境条件的影响,不育率和不育度均为100%,且无死蕾现象。
2.2 恢保关系的分析
在开花期,将7个恢复系(25cc1、9A001C、7-5、91-恢5900、‘恢5148’、298033和Hui01)以及3个甘蓝型油菜品种(‘华双4号’、‘华油8号’和‘中双9号’),分别与WNJ01A、Pol、Ogu、Kos进行测交,鉴定F1的育性。结果表明:91-恢5900、7-5、‘恢5148’和298033只能恢复Pol的育性,表明Pol与WNJ01A、Ogu、Kos有不同的恢保关系;恢复系25cc1、9A001C均能恢复Ogu和Kos的育性,却不能恢复WNJ01A和Pol的育性,说明Ogu和Kos恢保关系相同[18],但与WNJ01A和Pol有不同的恢保关系;同样地,Hui01只能恢复WNJ01A的育性,却不能恢复Pol、Ogu和Kos的育性。此外,‘华双4号’等3个甘蓝型油菜品种均不能恢复这4种不育系的育性。以上结果说明WJS01A具有与Pol、Ogu和Kos明显不同的恢保关系(表3)。
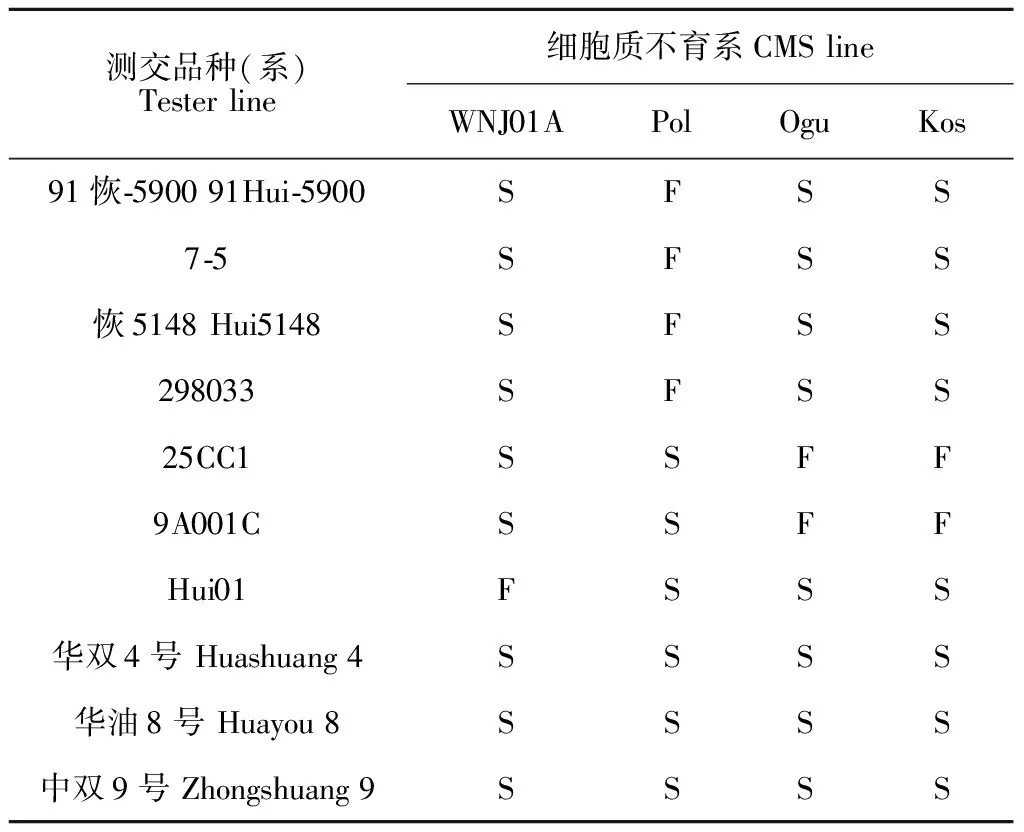
表3 不同胞质类型恢保关系鉴定结果
2.3 芥菜型油菜不育系WJS01A的细胞学分析
通过对不育系WJS01A花蕾横切石蜡切片观察发现,它在花药发育的早期阶段未出现孢原细胞和造孢细胞的形成和分化,整个花药在表皮以内都由性质均一的细胞组成。而且4个长雄蕊和2个短雄蕊的发育形态不完全一样,长雄蕊花药呈长梭形,短雄蕊花药则呈近似方形(图2, A、C、D)。同时,观察到花药的药隔维管束,没有孢原细胞和壁细胞的形成。表皮内部充满薄壁组织,且发生液泡化,没有形成典型的蝶形花药结构(图2, B)。当花药继续发育时,其外形和体积均有所增大,但除药隔维管束外,表皮以内仍保持无任何分化的薄壁细胞状态,而且维管束的韧皮部和木质部不发达(图2, C、D)。此外也发现少数材料的花药在孢原细胞早期停止发育,但表型仍表现为完全败育。由此认为,不育系WJS01A属无花粉囊型不育系,主要特点是不能形成正常的孢原细胞进而分化出花粉囊等花药结构,具体的败育时期应为花药原基到孢原细胞时期。
2.4 多重PCR结果分析
利用多重PCR对WJS01A、NRO4270A、Kos、Ogu和Pol 等雄性不育材料以及WJS01A来源不育系WNJ01A进行鉴定和区分。结果显示(图3),常规‘华双5号’仅在1 000 bp左右处存在特异条带。而Ogu和Kos胞质则在500 bp和1 000 bp左右存在特异条带的组合;NRO4270A胞质仅在500 bp左右处有一条特异条带,且与Ogu和Kos胞质的特异条带之一相同。Pol胞质在750 bp左右有一条特异条带,在500 bp左右有一条较淡的条带。
正常的芥菜型油菜3340在500 bp左右和1 000 bp左右处有2条较淡的条带。WJS01A和WNJ01A胞质的扩增结果与3340相同。从以上结论可以看出多重PCR标记可以将WJS01A与其他不育胞质类型的材料进行区分,但不能与正常的芥菜型油菜进行区分。
2.5 WJS01A线粒体基因组RFLP分析
利用atp1、atp6、atp9、cox1与限制性内切酶EcoR I和BamH I组成的8种探针/酶组合,对不同油菜不育胞质类型的线粒体基因组进行RFLP分析,结果显示(图4),在所有探针/酶组合中WJS01A与Ogu、Pol、Kos、NRO4270A不育胞质类型存在着显著不同数目或大小的条带。其中,在atp9/BamH I探针/酶组合中,尽管WJS01A与Kos、Ogu和Pol存在2条相似条带,但WJS01A仍存在另外1条特异条带(图4, F)。因此,这些细胞质类型材料的线粒体基因组间存在着显著的差异,而以上的探针/酶组合可以明显地区分WJS01A与其他不育类型,说明WJS01A是一种不同于Ogu、Pol、Kos、NRO4270A的胞质类型。
3 讨 论
本研究中细胞质雄性不育系WJS01A来源于芥菜型油菜农家品种中的天然不育株,该不育类型在不同的生态坏境下均表现为彻底败育,不受光照和温度的影响,且无死蕾现象。由此表明将WJS01A在育种上利用可以解决部分油菜不育胞质类型在杂交种制种中存在的微粉问题。
利用细胞生物学方法探究雄性不育花药败育过程中重要的结构变化,有助于对不育基因调控花粉败育发生机制的解析[24]。WJS01A花药发育的细胞学研究结果显示,在花药发育早期阶段无孢原细胞和造孢细胞的形成与分化,同时,少数材料的花药在孢原细胞早期出现异常发育。因此,WJS01A败育时期为花药原基到孢原细胞时期。Pol花药败育时期为孢原细胞分化期[25],而Ogu败育大多受阻于小孢子四分体至单核花粉时期[26]。与Pol和Ogu相比,WJS01A在花药发育上有明显的不同,因此它是不同于Pol和Ogu的一种不育胞质类型。此外,研究发现WJS01A与另外3种同属芥菜型油菜来源的不育系hau、欧新A和orf220-type在花器官形态和花药败育特征均有差异[27-29]。其中,hau的不育性由不育基因orf288控制[30],它的雄蕊退化为花瓣状,且其败育关键时期是雄蕊原基分化期[27];欧新A的不育性由不育胞质和核内1对隐性基因控制,它的花蕾表现为“突柱段”的柱头外露[31];orf220类型不育系受线粒体基因orf220控制,它的雄蕊退化为瓣状和心皮状等不同变异类型[29]。同时,我们的研究观察发现WJS01A的花药维管束不发达,这与前人在大量雄性不育材料中发现维管束组织异常的现象相一致[32]。
植物细胞质雄性不育通常是由线粒体基因组重排产生的异常嵌合基因导致的,不同败育胞质类型往往具有其特异的异常嵌合基因,因而不同胞质类型间的败育机理存在着较大的差异[3]。本研究利用线粒体基因多重PCR方法对WJS01A、NRO4270A、Kos、Ogu和Pol等雄性不育材料以及WJS01A甘蓝型油菜背景不育系WNJ01A进行鉴定和区分。结果表明多重PCR标记可以将WJS01A与其他不育胞质类型区分开,但却无法和正常的芥菜型油菜进行区分,证实了该WJS01A与orf138、orf222/nad5c和orf224不育基因并无关联,但他与正常芥菜型油菜线粒体基因组之间依然存在高度同源,因此需进一步筛选特定多重PCR标记方可区分两者之间的差异。此外,利用RFLP对WJS01A及Pol、Ogu、Kos和NRO4270A等不育材料的线粒体基因组进行分析,检测结果显示在所有的探针/酶组合中,WJS01A在条带数量和大小上均显示出与其他4种不育胞质类型的显著差异。以上结果说明不育系WJS01A与其他不育胞质类型的线粒体基因组存在显著变异,是一种不同的油菜细胞质雄性不育胞质类型。WJS01A细胞质不育类型的发现不仅丰富了中国油菜细胞质雄性不育类型,而且将有利于拓宽目前油菜杂种优势利用的遗传基础。
