TPM1 基因相关多态性与中国北方汉族人群非综合征性唇腭裂的相关研究
2022-03-07姜宇欣刘婷婷时旭恒
回 翔, 姜宇欣, 刘婷婷, 时旭恒, 王 琪, 宋 涛
(哈尔滨医科大学附属第一医院口腔颌面外科,黑龙江 哈尔滨 150001)
唇腭裂是人类常见的头面部出生缺陷,全世界活产儿中,唇腭裂发病率约为1‰~2‰[1]。 亚裔和美洲原住民发病率最高,欧洲族裔次之,非洲人群最低[2]。 根据是否伴有其他疾病,唇腭裂又分为综合征性唇腭裂(syndromic orofacial clefts, SOC)和非综合征性唇腭裂(nonsyndromic orofacial clefts, NSOC)两大类。 在NSOC 中,根据其所波及的解剖学结构,可分为非综合征性单纯唇裂 (nonsyndromic cleft lip only,NSCLO)、非综合征性唇腭裂(nonsyndromic cleft lip with or without cleft palate, NSCLP)及非综合征性单纯腭裂 (nonsyndromic cleft palate only, NSCPO)。NSOC 的病因包括多基因的相互作用[3]和环境的易感性[4]。
单核苷酸多态性(SNPs)在人类全基因组中有广泛的分布[5],同一SNPs 或单体型在不同的环境因素和不同的种族中具有不同的致病效力[6]。 比如,rs4444903 与巴西人群NSOC 不相关,但与中国北方人群相关;rs1799929 与阿根廷人群NSOC 相关,但与中国北方人群不相关[7]。
Ludwig 等[8]在2012 年报道了对欧洲人群大样本全基因组关联分析 (genome-wide association study,GWAS)的荟萃分析及巴尔的摩欧洲人群亲本三元样本独立研究,结果表明,rs1873147 无论在荟萃分析(P=2.81×10-8)中还是独立研究中(P=2.37×10-3)都对NSOC 的发病具有显著影响,但其在亚洲人群中没有显著性关联。 同时, 该研究认为,rs1873147 位于TPM1 基因的上游约20 kb,且该区域的变异映射到包含增强子和启动子及多个转录因子结合位点的调节区。 该研究同时发现,rs7179658(P=2.13×10-8)和rs4775599(P=6.79×10-9)在欧洲人群中具有全基因组意义,同时3 个位点完全连锁(r2=1.0)。Jia 等[9]在欧美人群三元样本和欧洲人样本的分析研究中验证了rs7179658 具有全基因组显著性,同时认为rs7179658 与rs1873147 位于同一连锁不平衡区块内,但是在单个群体或合并分析中并没有重复出rs7179658 与NSOC 的关联。
Pan 等[10]在中国江苏汉族人群中验证了rs1873147与NSCPO 相关, 其变异等位基因T 对单纯腭裂(cleft palate only,CPO)有保护作用。该团队于2015 年在TPM1 与中国华东地区汉族人群NSOC 的研究[11]中, 通过标签SNP 策略进一步论证了TPM1 与中国汉族人群NSOC 的关联。
综上,为了能够确保TPM1 上游SNPs 分析设计的成功,本研究分析了过往研究中TPM1 上游调控区域完全连锁的3 个显著遗传变异(rs7179658、rs1873147 和rs4775599)与中国北方汉族人群NSOC样本的关联, 并且进一步分析了与这些位点可能相关的基因变异在NSOC 组和其亚组风险的相关性。
1 材料和方法
1.1 样本采集
收集就诊于哈尔滨医科大学附属第一医院和哈尔滨医科大学第二附属医院口腔科的性别、年龄、经济状况、受教育程度无显著差异的335 例汉族NSOC 患者(病例组)和572 例汉族健康对照者(对照组)。 本研究按照哈尔滨医科大学院校评审委员会的规定进行, 并征得所有参与者或其法定监护人的书面知情同意。 对照组来自同一地区基因无相关的中国汉族人群,均无任何发育异常、唇腭裂家族史或其他先天出生缺陷家族史, 由专业口腔外科医生仔细检查。 取每位入组者外周静脉血约2 mL,进行基因分型分析。 将病例组样本分为3 个亚组,单纯唇裂(cleft lip only,CLO)102 例、唇腭裂(cleft lip with palate,CLP)93 例、单纯腭裂(CPO)140 例。
1.2 DNA 提取和基因分型
我们通过DNA 纯化试剂盒 (Axygene 公司,美国)从静脉血中提取人类基因组DNA。 基因分型用于聚合酶链反应-连接检测反应 (polymerase chain reaction-ligation detection reaction, PCR-LDR)。 采用多重PCR 方法扩增出目的DNA 序列。 每个样本总体积为20 μL,包括DNA 1 μL(50 ng)、1×Buffer 2 μL、Mg2+0.6 μL (3 mmol/L)、dNTP 2 Taq DNA连 接 酶(1 U)、ddH2O 12.2 μL、引 物Mix 2 μL(0.5 pmol/L)。 扩增周期分别为95 ℃,2 min;94 ℃,30 s;62 ℃,90 s;72 ℃,1 min;72 ℃,10 min。 最终1 个体积为10 μL 的样本,包括1×Buffer 1 μL、探针混合物1 μL (2 pmol/μL)、TaqDNA 连接酶0.05 μL(2 U)、ddH2O 4 μL、多重聚合酶链反应产物4 μL。采用95 ℃,2 min;94 ℃,15 s;50 ℃,25 s, 进行40 个循环。 用3730DNA 测序仪(不列颠哥伦比亚省温哥华生命技术公司,加拿大)对所得产品进行鉴定。 为了进行质量控制,随机抽取了10%的DNA 样本,并对其进行基因分型,结果完全一致。
1.3 方法及标准设定
用SHEsis 软件(http://analysis.bio-x.cn/)对病例组和对照组所有的SNPs 进行哈迪温伯格平衡(Hardy-Weinberg equilibrium, HWE)检验,病例组和对照组等位基因频率分布差异的χ2检验,连锁分析和单体型分析。 以D’和R2值作为评估连锁不平衡(linkage disequilibrium, LD)的标准,并认定当D’和R2值同时大于0.8 时,位点之间存在明显的连锁不平衡;当D’和R2值同时等于1 时,表明完全连锁,SNPs 的相关信息可以相互替代, 则不计算单体型。通过SPSS 22.0 软件计算基因型频率分布差异的χ2检验(T>5 取Pearson 检验P值;1<T<5 取连续校正的P值),以比值比(odds ratios, ORs)和95%置信区间(confidence interval, CI)来检验SNPs 与NSOC 或其亚组风险的相关性。P<0.05 被认为是具有统计学意义上的相关性。 所有的数据测试都是双侧的。 查询PubMed 生物信息数据库(https://www.ncbi.nlm.nih.gov/pubmed)检索本研究SNPs 的相关信息。
2 结果
2.1 SNPs 与TPM1 基因位置关系
如表1所示,3 个SNPs 位于TPM1 上游22~24 kb,在Ludwig 等[8]的研究中,该区域存在多个连锁的SNP,且该区域与调控元件相关。
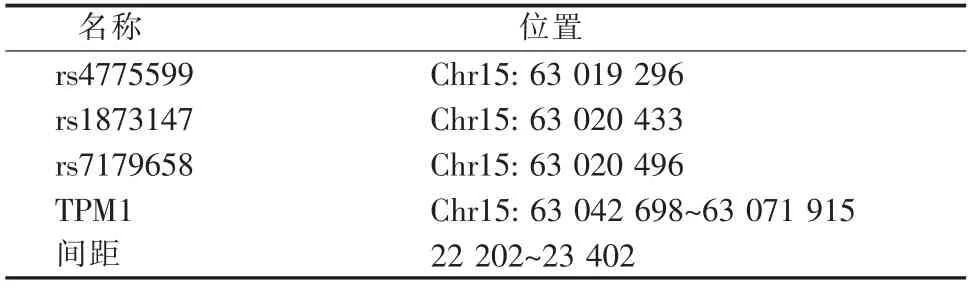
表1 SNPs 和TPM1 基因Table 1 SNPs and TPM1
2.2 不同人群等位基因的比较和HWE 检验
由表2 可知,该位点在不同种族中存在等位基因频率的差异,本组对照组人群参考等位基因频率与东亚人群较接近,分别相差0.019、0.016 和0.016,同时该结果远高于世界范围水平。用SHEsis 软件对各组样本进行HWE 检验,等位基因无显著性差异(P>0.05),各组人群遗传平衡(表3)。

表2 不同人群等位基因的差异Table 2 Differences of alleles in different populations

表3 哈迪温伯格平衡检验Table 3 Hardy-Weinberg equilibrium test
2.3 连锁分析和单体型分析
如图1 所示,rs7179658、rs1873147 和rs4775599的3 个SNPs 在本研究样本的各组中都处于强连锁不平衡状态(图1),D’=R2=1。 故3 个SNPs 的相关信息可以相互替代,同时不计算单体型。
图1 TPM1 相关3 个SNPs 连锁分析Figure 1 Linkage analysis of three SNPs related to TPM1
2.4 SNPs 基因型和等位基因的统计分析
选取rs1873147 为代表的以最小等位基因频率(minor allele frequency, MAF)基因为关注基因(等位基因T),将等位基因频率分布及基因型频率分布的统计检验P值、OR 值、95%CI 整理于表4 中。 以病例组相对于对照组等位基因频率较高者为风险等位基因(表5),把风险等位基因设计为关注基因,将rs7179658、rs1873147、rs4775599 基 因 型 频 率 及等位基因频率分布的统计检验P值、OR 值、95%CI作对比整理分别整理于表6~表9, 其中CPO 组的风险等位基因分别为等位基因C、等位基因T、等位基因G,其余各组分别为等位基因T、等位基因C、等位基因A。
2.4.1 rs1873147 等位基因频率的分析 如表4 所示,rs1873147 等位基因的分布在NSOC 组中没有统计学意义,进一步的分层研究显示,其等位基因频率在CLP 组和对照组之间的差异有统计学意义(P=0.035),在其他亚组中没有统计学意义。该结果不仅显示了等位基因T 对NSOC 亚型的影响存在差异,同时也提示分层研究可能具有更高的效能,rs1873147 等位基因T 可能对NSOC 的发病起保护作用。 rs1873147 遗传模型在各组中都没有显著性差异(P>0.05)。
2.4.2 风险等位基因分析 由表4 可知, 由于所关注的MAF 基因并不是所有分组的风险等位基因(如NSOC 组、CLO 组、CLP 组),在表5 中我们对各组的风险等位基因进行了确认, 并计算了OR 值和95%CI。 由表5 可知, 在CLP 组中,rs1873147 的等位基因C 对于对照组存在显著性差异,结果具有统计学意义(P<0.05),且等位基因C 在CLP 组中增加了发病的风险[1.723 (1.033~2.875)], 这说明对于CLP 的患者而言,rs1873147 的突变是有益的。
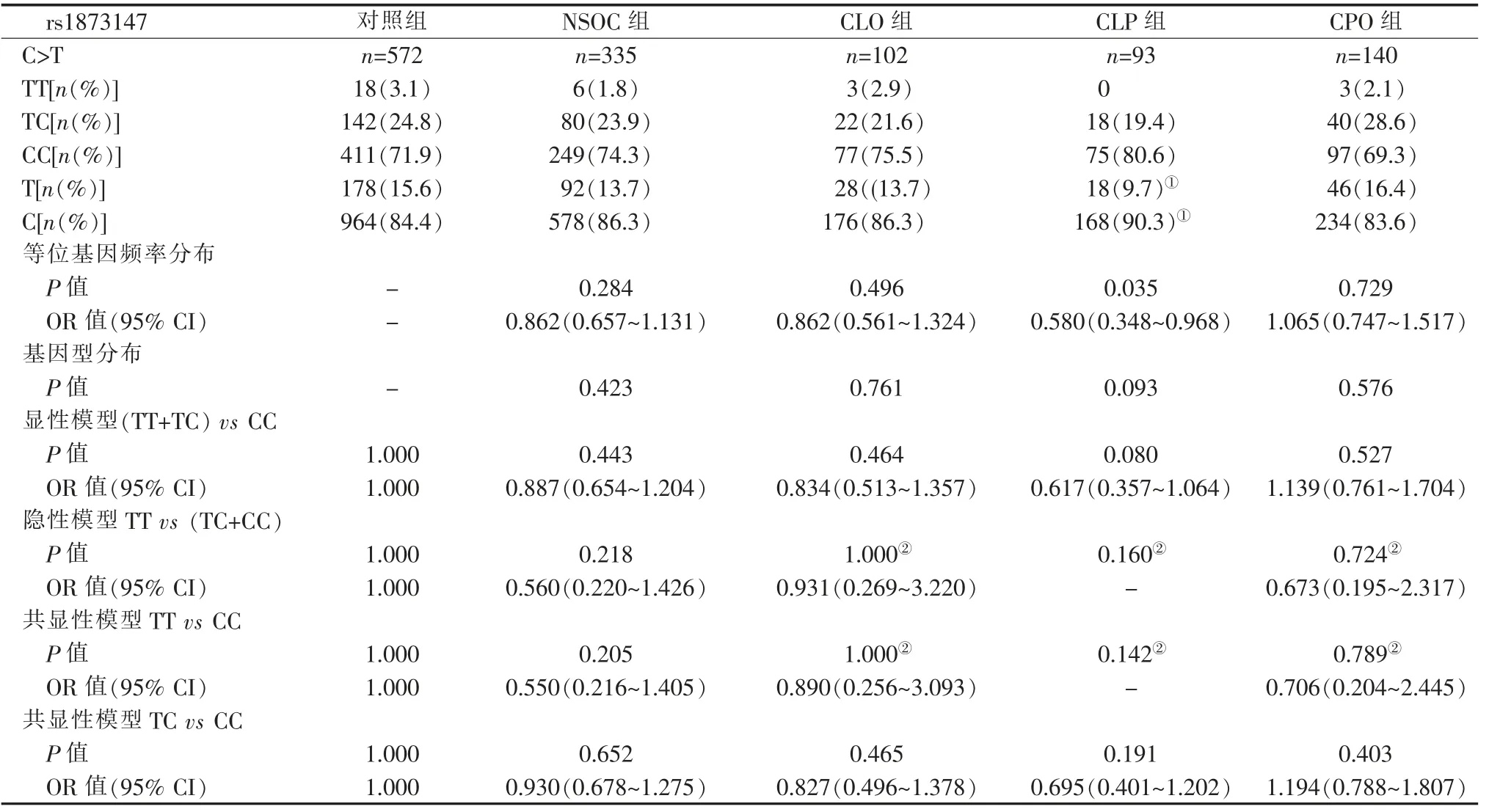
表4 rs1873147 等位基因、基因型分布Table 4 Distribution of alleles, genotypes of rs1873147
2.4.3 遗传模型的分析 根据表5 所确定的风险等位基因, 我们对所筛选的3 个SNPs 进行了遗传模型的分析,并进行了纵向的比较(表6~表9)。 这种完全连锁确实导致了rs7179658、rs1873147 和rs4775599 遗传信息的一致性。 对各种遗传模型的分析没有发现显著性的差异。

表5 风险等位基因频率分布对照Table 5 Comparison of risk allele frequency distribution

表6 NSOC 组基因型分布Table 6 Genotypes distribution of NSOC group
3 讨论
本研究筛选了在其他人群中被证明与NSOC发病存在关联的3 个SNPs,实验结果进一步表明这些位点与中国北方汉族人群NSOC 发病存在关联,差异具有统计学意义,同时这些位点高度连锁。
Qian 等[11]认为,基于NSOC 易感位点rs1873147与TPM1 可能存在调控关系, 同时TPM1 变异可增加NSOC 的易感性, 原因可能为TPM1 提供了肌动蛋白细丝的结构稳定性和调节细胞骨架的功能,TPM1 变异对果蝇头部发育产生影响, 并可致胚胎小鼠死亡,以及其在颌面部非肿瘤中的炎性作用可以明显下调牙髓TPM1 的表达。
在哺乳动物细胞中,TPM 家族由4 个独立的基因表达(TPM1、TPM2、TPM3、TPM4),并且通过每个基因的启动子选择和剪接的交替进行,可以产生多达40 个差异性的mRNA 变体, 其中大多数已经被证明在不同的组织中以蛋白质的形式表达[12]。 原肌球蛋白(由TPM 编码)与肌钙蛋白(troponin, TN)复合,通过该复合物对横纹肌收缩进行调节[13]。虽然它在颅面发育中的确切作用尚不清楚,但已经发现它与孤立的马蹄内翻有关,后者是一种相对常见的出生缺陷[10],说明其可在胚胎发育期产生效应。

表7 CLO 组遗传模型Table 7 Genetic model of CLO group

表8 CLP 组遗传模型Table 8 Genetic model of CLP group

表9 CPO 组遗传模型Table 9 Genetic model of CPO group
TPM1 是肥厚型心肌病的相关基因[14]。 Liu 等[15]在对小型猪次级腭发育相关的mRNA 表达分析研究中发现了一些与心脏发育相关的基因, 其中,由TPM1 等蛋白组成的肌钙蛋白-原肌球蛋白复合体,在肌肉收缩的调节中发挥中心作用[16]。在胚胎的腭架上可以识别出发育中的骨骼肌细胞,某些与腭裂相关基因的功能丧失与次级腭架内一些肌肉相关基因表达改变存在关联[17]。 这些都从侧面暗示了TPM1 可能参与了NSOC 的发生, 本研究的差异性表达也出现于包含腭裂的CLP 亚组中,与过往的研究结果相一致。
TPM1 及其3'非翻译区与转录及翻译相关的因子结合可以抑制肿瘤的生长[18-19]。NSOC 可能与肿瘤存在共同的遗传背景, 如6p21.33、20q12 不仅与NSOC 相关,还分别与滤泡性淋巴瘤和皮肤鳞状细胞癌有关[20]。 NSOC 中TPM1 上游5'端的相关报道较少, 所有的文献最终的追溯都来自于Ludwig 等[8]的研究。Jia 等[9]认为,rs7179658 与rs1873147 来自于相同的连锁区,可以用来替代对后者的研究,但该团队并没有复制到显著性差异的结果。同样,rs4775599也来自于该连锁区域。 我们实验的结果也证明了在中国北方人群中,rs7179658、rs1873147 及rs4775599存在连锁不平衡。 基于现有证据, 我们不能判定rs7179658、rs1873147、rs4775599 的作用是其本身所造成的或是由于与真正致病的位点连锁而产生的。
本研究结果与以往关于欧洲裔的GWAS 和GWAS meta 分析的结果[8]存在差异,该研究认为这3 个位点存在连锁但与亚洲人群NSOC 无关, 而本研究认为TPM1 上游3 个完全连锁的SNPs 与中国北方汉族人群NSOC 相关, 野生型等位基因可能增加了NSOC 的发病风险,突变型等位基因可能降低了NSOC 的发病风险。
de Aquino 等[21]对巴西2 个地区人群进行了结构化的方法研究, 并确认rs1873147 是巴西非洲裔NSOC 的易感标记,巴西东北地区纯合子基因型CC的发病风险可达TT 基因型的2 倍以上[OR(95%CI)=2.47(1.29~4.74)],但对于该位点与TPM1 的关系,该团队仅仅复述过往研究的观点。 同时该巴西人群的风险等位基因为C 等位基因,与我们的实验结果一致。NSOC 发病除了与传统遗传学DNA 一级结构变化有关外, 也有可能与表观遗传学中DNA 及组蛋白的修饰有关[22],这也提示我们需要进一步的研究。 除了本研究中的3 个SNPs 外, 我们还检索了Ludwig 等[8]所提供的该区域其他8 个完全连锁的SNPs,遗憾的是并没有发现相关的研究。
本研究的结果是TPM1 上游SNPs 为中国北方汉族人群NSOC 的发病提供了依据。 但是,本次研究仍然有一些限制。 我们的样本规模相对较小,当我们将NSOC 分为4 组时,我们需要继续收集更多的样本。 本研究的SNPs 所反映的遗传信息比较单一, 应该超越该连锁群进一步扩大SNPs 的搜索范围。 另外,TPM1 及TPM1 上游在颅面发育中的确切作用尚无直接证据,需要进一步的研究。 此外,基因与环境之间的相互作用的多因素研究,表观遗传学的修饰对我们下一步的实验设计也很重要,这可能在NSOC 的病因学研究及遗传表型的差异化表达中也起到至关重要的作用。
综上所述,TPM1 相关rs7179658、rs1873147 和rs477559 与中国北方汉族人群NSOC 可能存在关联, 野生型等位基因增加了NSOC 的发病风险,突变型等位基因对NSOC 发病具有保护作用。rs7179658、rs1873147 和rs477559 在中国北方汉族NSOC 人群中完全连锁。
