楸树CbuATX1,CbuATX1-like和CbuATX2基因克隆及生物信息学分析
2022-03-02赵佳明樊二勤王军辉曲冠证
赵佳明 樊二勤,2 刘 轶 王 智 王军辉 曲冠证
(1. 东北林业大学林木遗传育种国家重点实验室,哈尔滨 150040;2. 中国林业科学研究院林业研究所,林木遗传育种国家重点实验室,国家林业和草原局林木培育重点实验室,楸树国家创新联盟,北京 100091;3. 中国检验检疫科学研究院,北京 100176)
开花是由内部遗传因素和外界环境因素共同调节的复杂生命过程,是植物进入生殖生长具备繁殖能力和遗传能力的象征。在模式植物拟南芥()中,有近百个开花相关基因被鉴定出来,例如,,,,等。这些基因主要通过光周期途径、春化途径、自主途径和赤霉素途径等多种途径发挥功能,同时与,,等开花整合因子相互关联共同调控植物开花。目前开花机制与基因调控网络在拟南芥中研究较为深入,而在其他物种中研究较少,尤其是在木本植物中。
表观遗传学(epigenetics)通常被理解为DNA序列没有发生改变的情况下基因功能或细胞类型的可遗传变化。介导表观遗传的分子机制通常有3 种形式,即DNA 甲基化(DNA methylation)、组蛋白翻译后修饰(histone post-translational modifications,hPTM)和非编码RNA(non-coding RNA,ncRNA)。通常不同类型以及不同位点的甲基化修饰具有不同的效应,如H3K4me3、H3K36me3 和H3K79me3 与基因的转录激活和常染色质有关,H3K27me3、H3K9me2∕me3多与基因沉默和异染色质相关。组蛋白H3K4的甲基化是细胞修饰启动子和增强子的1 种方法,组蛋白H3K4 的三甲基化(H3K4me3)主要聚集在转录活跃的启动子区,而一∕二甲基化(H3K4me1,H3K4me2)则主要集中在增强子区域。拟南芥同源基因()是一种已知的花发育调节因子,它是一种H3K4组蛋白甲基转移酶,主要通过调节细胞周期的持续时间、细胞产量以及从根尖分生组织中的细胞增殖到细胞伸长的转变来控制根的生长。研究发现拟南芥基因是根发育过程中细胞产生、构型和形态发生所必需的。在维管植物中,厚壁组织细胞的次生壁增厚受到一个复杂的转录因子网络的严格控制。通过对拟南芥全基因组分析显示,在茎发育过程中,参与次生壁形成的基因上调在很大程度上是通过增加H3K4三甲基化水平来调控的,在拟南芥H3K4me3 的所有组蛋白甲基转移酶中,ATX1在花序茎发育过程中显著增加,结果表明ATX1在拟南芥花序茎发育过程中对束间纤维次生壁生物合成的调控起着关键作用。
β 族乙酰氨基葡萄糖(β-GlcNAc)对蛋白质的翻译后修饰是由O-GlcNAc 糖基转移酶(OGlcNAc transferases,OGTs)催化完成,这种糖基化修饰参与调控细胞内多种重要的生物学过程。O-GlcNAc 转移酶SEC(SECRET AGENT)的功能的缺失会导致拟南芥提前开花表型,且突变体中开花时间的负调节因子FLC 的转录受到抑制,同时FLC 染色质区组蛋白H3K4me3 修饰水平显著降低,表明植物体内糖基转移酶SEC 参与表观遗传介导的开花时间调节过程。FLC 位点组蛋白的H3K3me3 修饰由组蛋白甲基转移酶ATX1 催化完成,该研究发现SEC 可以直接催化ATX1使其获得O-GlcNAc修饰,在体外及体内条件下,SEC均表现出通过O-GlcNAc 修饰而激活ATX1 的组蛋白甲基转移酶活性。而且,遗传分析表明ATX1的功能依赖于SEC,所以表明拟南芥O-GlcNAc 转移酶SEC激活组蛋白甲基转移酶ATX 1 可以调节开花。利用全基因组亚硫酸氢盐测序、RNA-seq 和RTPCR 联合分析表明,SDG25 和ATX1 的突变降低了组蛋白H3K4me3 的水平,增加了DNA 胞嘧啶甲基化,并抑制了拟南芥在逆境恢复过程中热胁迫应答基因的表达,ChIP-qPCR 结果证实ATX1 与这些靶基因相关的染色质结合,结果证明SDG25 和ATX1 是拟南芥耐热胁迫所必需的。铜是生物体必需的微量元素,体内铜平衡机制是蛋白质介导的,它将细胞内的铜传递给靶蛋白,这一途径是通过铜伴侣蛋白实现的。特定的蛋白质,即所谓的铜伴侣,介导有害金属的运输。研究发现ATX 参与铜向受体转移的转运蛋白,并发现ATX1家族的细胞质伴侣蛋白和膜结合型ATPase 参与了铜的转运。
对高度保守重复基因的拟南芥和进行了比较研究,揭示了部分冗余和功能分化的特征。尽管它们在结构上相似、同源物高度相似但编码的蛋白具有不同的生化功能以及调控序列。和很大程度上影响了互不重叠的基因集的表达,研究发现只甲基化了一小部分核小体。即使在共同调节共享靶标时和也可能采用不同的机制。两种蛋白质都使组蛋白H3K4甲基化,但当将其三甲基化时,将其二甲基化。有研究报道了基因、三胸基因家族(Trithorax group genes)基因、多梳基因家族(polycomb group genes)基、以及ULTRAPETALA 1 基因在早期花发育过程中的相互作用,和在表观遗传调控基因表达方面具有拮抗作用。TrxG 蛋白家族作为最重要的表观遗传调控元件之一,参与依赖于染色质修饰的转录调控过程,TrxG 蛋白家族在生物进化过程中极其保守,并且已经被证实参与多类生物发育调控相关基因的激活表达。在拟南芥TrxG家族中发现()和()功能缺失突变体对ABA 更加高敏,实验证明在ABA 诱导下,和结合非磷酸化的RNA 聚合酶II,通过H3K4me3 修饰激活的表达,从而参与ABA和干旱胁迫响应过程。
植物开花是十分复杂的过程,作为植物重要的性状之一,对于木本植物的经济价值和环境适应性具有极其重要的意义,然而由于木本植物童期较长,基因组复杂等,使得木本植物开花分子机制的研究比较缓慢。楸树(C.A.Mey.)隶属紫葳科(Bignoniaceae)梓树属()多年生木本植物,是我国特有的珍贵用材和具园林观赏价值的古老树种,多种植于皇家园林。楸树童期时间较长,种植五年以上开花,花期为10~15 d。百日花楸树是以优良杂种无性系“洛楸4 号”和滇楸为亲本杂交培育的新品种,种植当年即可开花,花期约为100 d。与普通楸树相比,百日花楸树的发现为研究楸树开花分子机制提供了十分宝贵的材料。通过本课题组前期转录组数据发现,基因在百日花楸与普通楸树中表达具有显著差异,对其进行研究以期能够进一步阐述百日花楸树开花的性状,为楸树开花机制的研究和楸树的定向遗传改良提供理论支持。
1 材料与方法
1.1 实验材料与主要试剂
以嫁接的一年生普通楸树()无性系9-1和突变株系—百日花楸树为实验材料,样品由中国林业科学研究院本课题组提供,在河南省洛阳市完成样品采集。
DNA 胶回收试剂盒,大肠杆菌质粒提取试剂盒购自杭州博日科技(Bioer)有限公司;大肠杆菌()Trans1-T1 感 受 态,pEASY-T1 Clonging Vector 购自北京全式金生物技术(Trans-Gen Biotech)有限公司;RNA 提取试剂盒购自北京百泰克生物技术(BioTeke)有限公司;反转录试剂盒,PrimeScripRT reagent Kit,TB GreenPremix ExⅡ,酶,dNTP Mixture 等均购自Ta-KaRa 公司(大连);其他试剂为进口或国产分析纯;所用引物合成、测序服务由擎科生物技术有限公司完成(哈尔滨)。
1.2 方法
1.2.1,和基因克隆
分别采集普通楸树9-1 系和百日花楸树处于营养生长期、生殖生长期的混合芽,放入无RNA酶冻存管中,用液氮快速冷冻后,储存于超低温冰箱中备用。参照百泰克生物公司RNA提取试剂盒说明书,提取样品的Total RNA,利用Nanodrop 2000C分光光度计检测RNA 的纯度和浓度,取0.5 μg RNA 为材料,利用PrimeScriptRT reagent Kit 试剂盒反转录合成第一链cDNA。根据楸树在营养生长期和生殖生长期的发育时期转录组数据进行,和差 异 基 因 筛选,并根据未公布的楸树基因组数据设计楸树,和基因特异性引物,扩增目的基因片段所用引物见表1。PCR 扩增反应体系为模板2 μL,引物Forward 和Reverse 各1 μL、10×ExBuffer 2.5 μL、Ex0.25 μL、dNTP 2 μL,用ddHO 补 足至25 μL,反 应条 件94 ℃5 min,94 ℃30 s,58 ℃30 s,72 ℃1 kB·min,40 个循环;最后72 ℃7 min;反应结束后取25 μL PCR 产物进行1%琼脂糖凝胶电泳,选取条带大小正确的进行胶回收。回收产物与pEASY-T 连接载体,转化大肠杆菌感受态细胞(Trans1-T1),涂于LB 固体培养基(含50 mg·mL卡那霉素),37 ℃培养12 h后,随机挑取单克隆并进行菌液PCR 检测,将条带位置正确的阳性单克隆菌液送往哈尔滨擎科生物公司测序。

表1 本实验中所用的引物Table 1 Primers used in this study
1.2.2,和基因生物信息学分析
利用BioEdit 软件进行氨基酸序列比对分析;利 用 在 线 软 件(https:∕∕www.ebi.ac.uk∕interpro∕search∕sequence-search)进行蛋白的保守结构域分析;利 用 在 线 网 站(http:∕∕web.expasy.org∕protparam∕)进行蛋白的理化性质分析;利用在线网站(https:∕∕npsa-pra-bi. ibcp. fr∕cgi-bin∕npsa automat.plpage=npsa_sopma.html)预测蛋白质二级结构;利用在线网站(https:∕∕swissmodel.expasy.org∕interactive)预测蛋白的三级结构及预测基因编码蛋白的功 能;利 用 在 线 网 站(http:∕∕web. Expasy. org∕protscale∕)进行亲∕疏水性进行分析;利用在线网站(https:∕∕embnet. vital-it. ch∕cgi-bin∕tmpred_form_parser)进行蛋白序列跨膜区的预测;利用在线网站(http:∕∕www.cbs.dtu.dk∕services∕SignalP-3.0∕)进行蛋白质信号肽分析;利用在线网站(http:∕∕www.cbs.dtu.dk∕services∕NetPhos)进行磷酸化位点预测;利用在线网站(http:∕∕bioinformatics.psb.ugent.be∕webtools∕plantcare∕html)进行启动子顺式作用元件分析;利用MEGA7.0软件构建系统进化树。
1.2.3,和基因在楸树混合芽不同发育时期表达量分析
利用Instegrated DNA Technologies 在线网站设计定量引物。参考TB GreenPremix ExⅡ荧光定量试剂盒反应体系,以基因为内参基因,以营养生长期和生殖生长期的普通楸树和百日花楸树为样品反转录得到的cDNA为模板,重复3 次,进行qRT-PCR 检测。数据分析 采 用2法,得到,和基因在普通楸树和百日花楸树中不同发育时期的相对表达量。
2 结果与分析
2.1 CbuATX1,CbuATX1-like 和CbuATX2 基因CDS全长克隆及其编码的氨基酸序列分析
通过对处于不同发育时期的混合芽样品进行转录组测序分析,我们从差异基因集中筛选得到差异显著的关键基因家族。根据楸树基因参考序列设计特异性引物,进行PCR 扩增,克隆得到了楸树,和基因,条带位置与预期相同(见图1)。连接pEASY-T 载体转化大肠杆菌后,菌液PCR 并送测序(见图2)。测序结果与转录组数据一致。基因CDS 全长726 bp,编码一个由241个氨基酸残基组成的蛋白序列;基因CDS全长801 bp,编码一个由266个氨基酸残基组成的蛋白序列;基因CDS全长1 554 bp,编码一个由517个氨基酸残基组成的蛋白序列(见图3)。
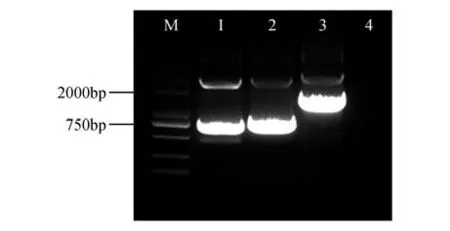
图1 CbuATX1、CbuATX1-like、CbuATX2基因PCR产物M.DNA marker DL5000;1.CbuATX1 基因的PCR 产物;2.CbuATX1-like 基因的PCR 产物;3.CbuATX2 基因的PCR 产物;4:ddH2O 作为阴性对照Fig.1 CbuATX1,CbuATX1-like,CbuATX2 gene PCR productsM.DNA marker DL5000;1.CbuATX1 Gene PCR products;2.CbuATX1-like Gene PCR products;3.CbuATX2 Gene PCR products;4:ddH2O as a negative control
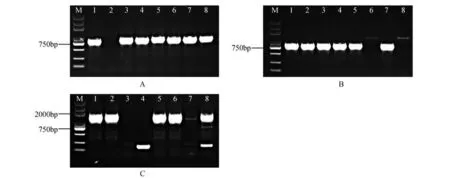
图2 大肠杆菌菌液PCR分析M.DNA marker DL5000;A.1~8,pEASY-ATX1菌液PCR检测结果;B.pEASY-ATX1-like菌液PCR检测结果;C.pEASY-ATX2菌液PCR检测结果Fig.2 PCR analysis of Escherichia coliM.DNA marker DL5000;A.1-8,pEASY-ATX1 PCR results of bacterial liquid;B.pEASY-ATX1-like PCR results of bacterial liquid;C.pEASYATX2 PCR results of bacterial liquid

图3 CbuATX1,CbuATX1-like和CbuATX2基因的cDNA序列及其编码的氨基酸序列A.CbuATX1基因的cDNA序列及其编码的氨基酸序列;B.CbuATX1-like基因的cDNA序列及其编码的氨基酸序列;C.CbuATX2基因的cDNA序列及其编码的氨基酸序列Fig.3 The cDNA sequence and amino acid sequence of CbuATX1,CbuATX1-like and CbuATX2 geneA.The cDNA sequence and amino acid sequence of CbuATX1 gene;B.The cDNA sequence and amino acid sequence of CbuATX1-like gene;C.The cDNA sequence and amino acid sequence of CbuATX2 gene
对楸树基因编码的蛋白质的理化性质进行预测分析(见表2),结果显示:蛋白的氨基酸中共含有18 个带负电荷氨基酸残基(Asp+Glu),37 个带正电荷氨基酸残基(Arg+Lys),不稳定系数为52.52,该值大于40,为不稳定蛋白,蛋白疏水性平均值为-0.550,为亲水性蛋白;蛋白的氨酸中共含有25 个带负电荷氨基酸残基(Asp+Glu),41个带正电荷氨基酸残基(Asp+Glu),不稳定系数为59.46,为不稳定蛋白,蛋白疏水性平均值为-0.627,为亲水性蛋白;蛋白的氨基酸中共含有78个带负电荷氨基酸残基(Asp+Glu),78 个带正电荷氨基酸残基(Arg+Lys),不稳定系数为41.71,为不稳定蛋白,脂肪族指数为78.43,蛋白疏水性平均值为-0.586,为亲水性蛋白。
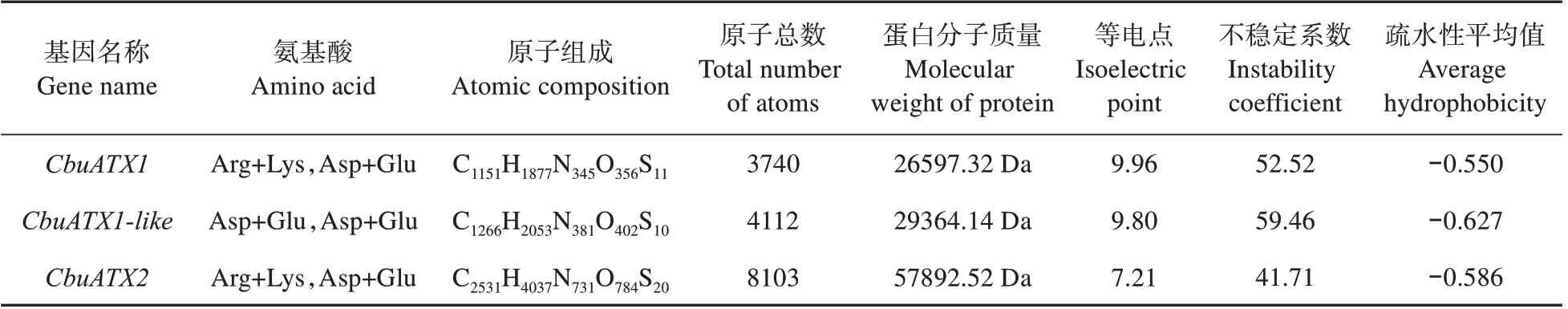
表2 蛋白的基本理化性质Table 2 Basic physical and chemical properties of protein
2.2 楸树CbuATX1,CbuATX1-like和CbuATX2蛋白质二、三级结构的预测分析
利用SOPMA 在线软件对蛋白质的二级结构进行预测和分析。结果表明:CbuATX1 和CbuATX1-like 蛋白的二级结构的含量中:无规则卷曲>延伸链>α 螺旋>β 转角;CbuATX2 蛋白的二级结构的含量中:无规则卷曲>α 螺旋>延伸链>β转角(见图4)。因此可推断,无规则卷曲和延伸链是CbuATX1 和CbuATX1-like 蛋白的主要组成成分,无规则卷曲和α 螺旋是CbuATX2 蛋白的主要组成成分。
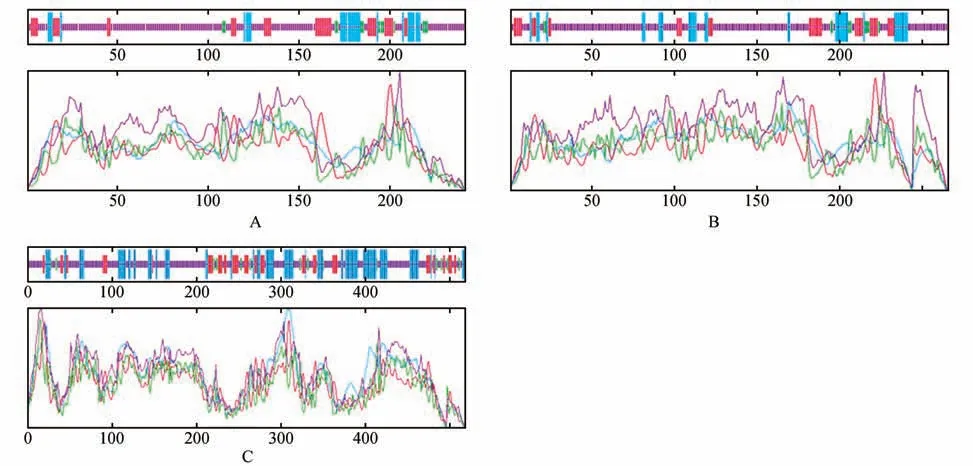
图4 蛋白的二级结构预测与分析A.CbuATX1蛋白的二级结构预测与分析;B.CbuATX1-like蛋白的二级结构预测与分析;C.CbuATX2蛋白的二级结构预测与分析Fig.4 Prediction and analysis of the secondary structure of proteinA.Prediction and analysis of the secondary structure of CbuATX1 protein;B.Prediction and analysis of the secondary structure of CbuATX1-like protein;C.Prediction and analysis of the secondary structure of CbuATX2 protein
蛋白质的多肽链在二级结构的基础上进行盘曲或折叠形成具有规律的三维空间结构。利用在线软件SWISS-MODEL 对蛋白质三级结构进行预测 分 析,获 得 了CbuATX1,CbuATX1-like 和CbuATX2 蛋白的三维结构模型,可知CbuATX1,CbuATX1-like 都以NBS-LRR class disease resistance protein 作为目标建模,均含有2 个α 螺旋,4个β 折 叠,组 分 间 均 以β 转 角(loop)相 连;CbuATX2 以Hepatoma-derived growth factor 作为 目标建模,含有1 个α 螺旋,4 个β 折叠,组分间以β转角(loop)相连;这3 种蛋白未形成复杂的三级结构(见图5)。
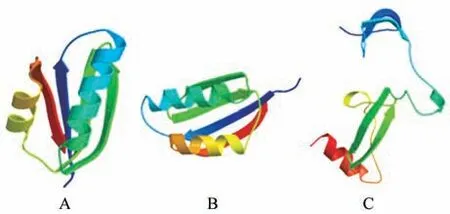
图5 蛋白三级结构预测A.CbuATX1 编码蛋白三级结构;B.CbuATX1-like 编码蛋白三级结构;C.CbuATX2编码蛋白三级结构Fig.5 Prediction of protein structureA.CbuATX1 coding protein tertiary structure;B.CbuATX1-like coding protein tertiary structure;C.CbuATX2 coding protein tertiary structure
2.3 楸树CbuATX1,CbuATX1-like和CbuATX2蛋白保守结构域分析
利用NCBI 的Conserved Domain Search(https:∕∕www.ncbi.nlm.nih.gov∕Structure∕cdd∕wrpsb.cgi)在线软件,对CbuATX1,CbuATX1-like 和CbuATX2 的氨基酸序列进行保守结构域的分析,结果显示:CbuATX1 和CbuATX1-like 蛋白有1 个相同的保守结构域,相同基序是LRVSLHC,KMRKHISRMEGVTSFNID,KKVTLTG 和VLSSIS。具有相同的保守结构域,均属于Heavy-metal-associated domain(HMA)superfamily,HMA 结构域包含两个半胱氨酸残基,它们在铜、镉、钴和锌等金属离子的结合和转移中起重要作用。CbuATX2 蛋白有2 个保守结构域,属于Pro-Trp-Trp-Pro(PWWP)superfamily,该结构域与赖氨酸、H4K20me 上甲基化的组蛋白结合,揭示它是甲基赖氨酸识别基序;也包含F∕Y-rich N-terminus(FYRN),它通常类似于trithorax∕ALL1 family proteins。
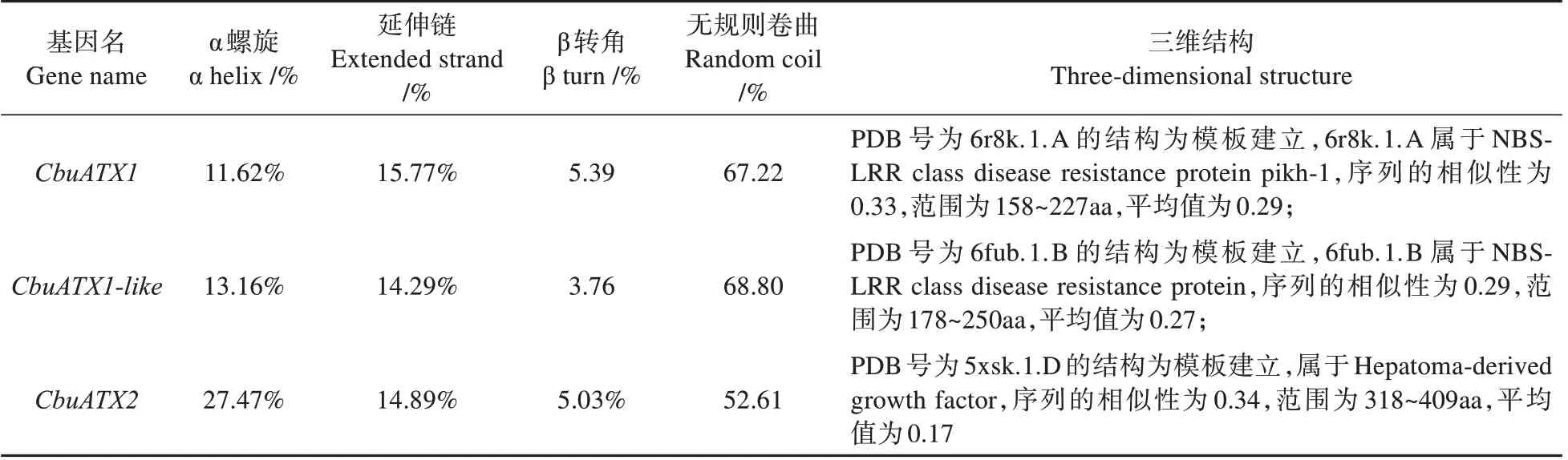
表3 蛋白二/三级结构组成Table 3 Protein secondary/tertiary structure composition
2.4 楸树CbuATX1,CbuATX1-like 和CbuATX2基因启动子顺式作用元件分析
根据实验室已测序的楸树基因组序列,选取基因上游1 500 bp 序列作为启动子序列,利用P1antCare 在线网站对楸树,和启动子顺式调控元件进行预测分析(见表4~6)。结果表明,基因启动子含有多个真核生物启动子的基本元件,如CAAT-box和TATA-box 等,还含有与光响应作用元件、生长发育作用元件(玉米醇溶蛋白代谢调控元件、分生组织诱导)、激素诱导元件(脱落酸诱导、赤霉素诱导、生长素响应元件)、胁迫元件(低温诱导、厌氧诱导、干旱诱导、防御和应激响应元件),此外还包括启动子和增强子区常见的顺式作用元件等,说明基因很可能与光响应密切相关并参与外界环境胁迫响应等过程。

表4 CbuATX1基因启动子顺式作用元件分析Table 4 Analysis of homeostasis elements in CbuATX1 gene promoter
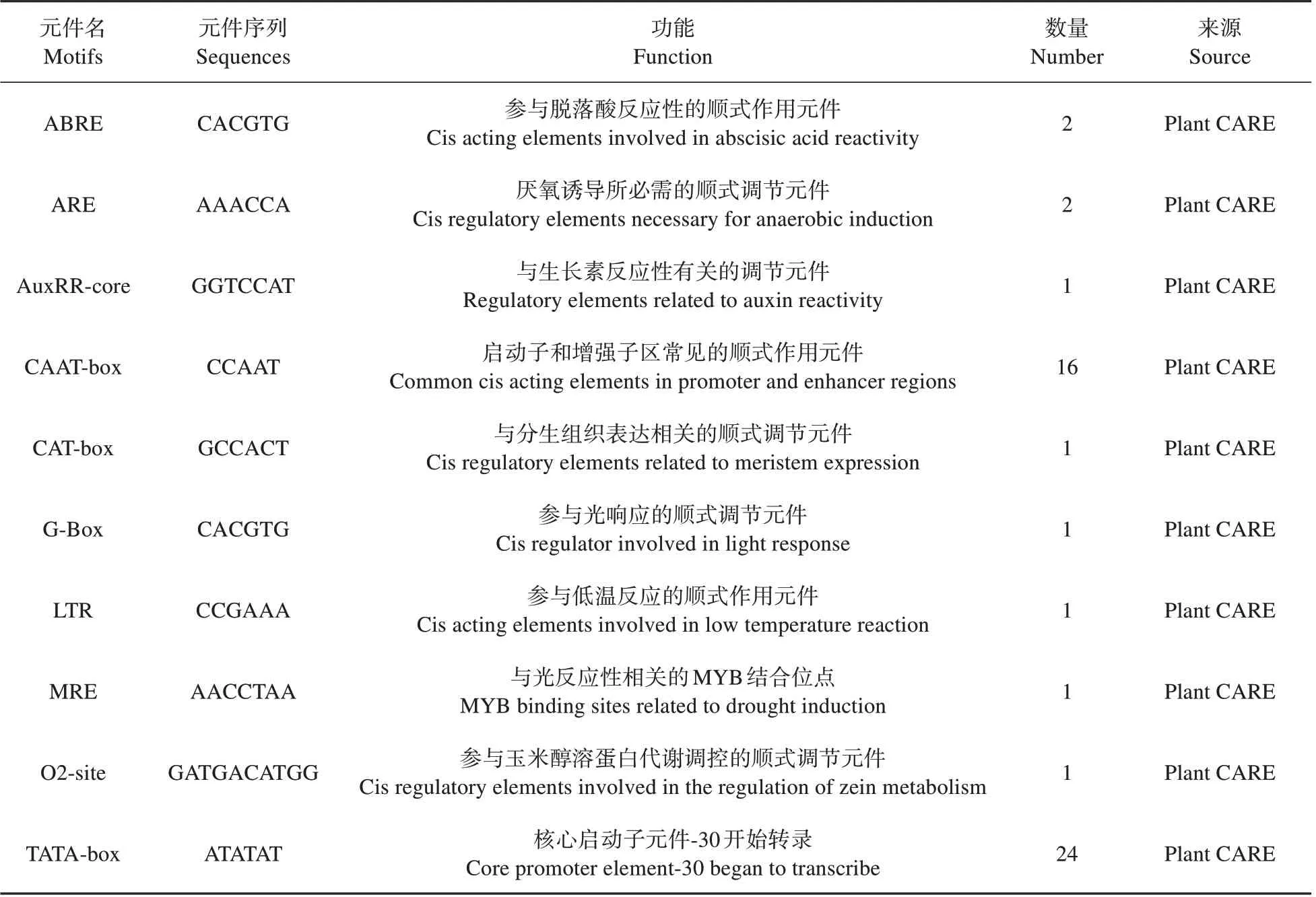
表5 CbuATX1-like基因启动子顺式作用元件分析Table 5 Analysis of homeostasis elements in CbuATX1-like gene promoter
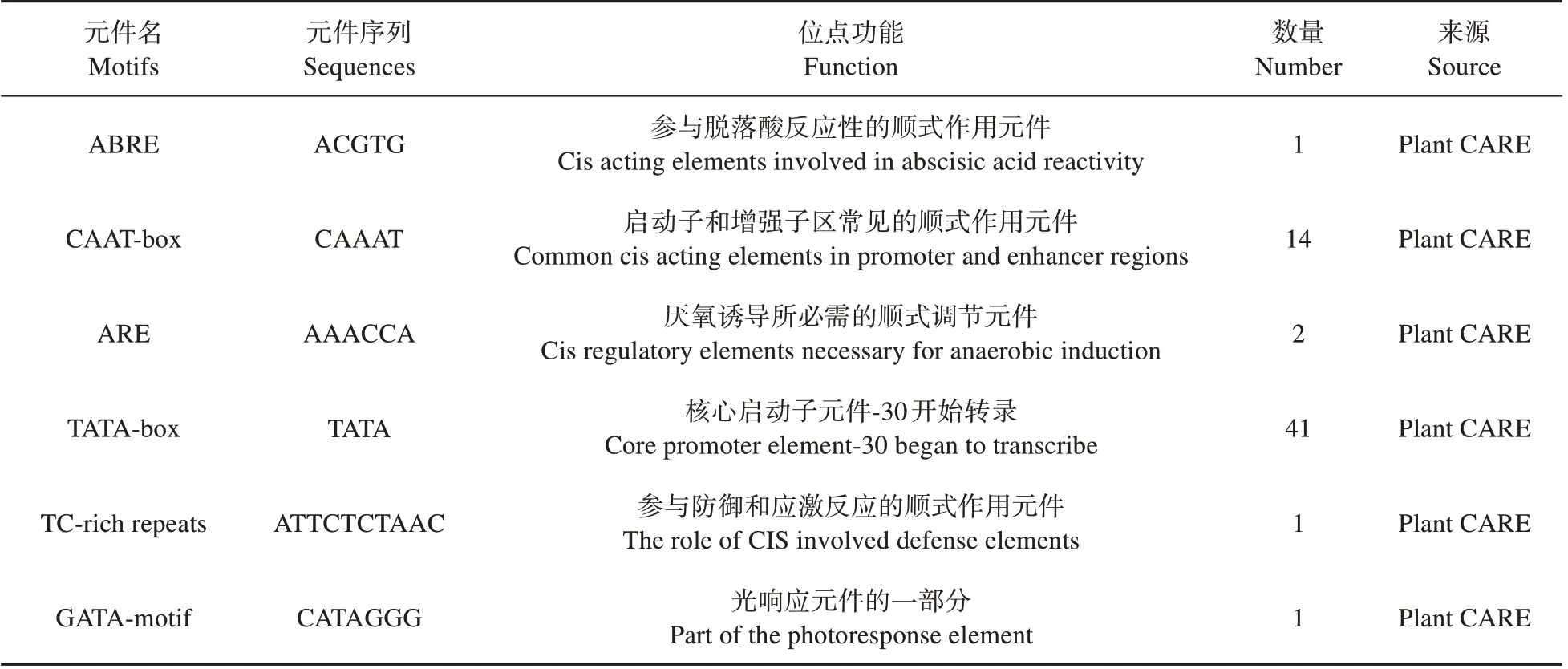
表6 CbuATX2基因启动子顺式作用元件分析Table 6 Analysis of homeostasis elements in CbuATX2 gene promoter
2.5 楸树CbuATX1,CbuATX1-like和CbuATX2氨基酸同源性分析
利用NCBI 数据库中的blastp 程序,以楸树CbuATX1,CbuATX1-like 和CbuATX2 氨基酸序列为检索对象,对不同物种进行比对,筛选得到与这3 个氨基酸序列同源性较高且具有代表性物种的同源序列,利用BioEdit 软件中的ClustalW 工具对上述不同物种的同源序列进行多重序列比对分析,结果发现:CbuATX1和CbuATX1-like蛋白在不同物种中具有高度保守性,与芝麻()相似性较高,CbuATX1 和CbuATX1-like 蛋白与其他物种相同的基序有QVV,LRVSL,RKH,MEGVTSFNI,KKV 等(见图6A)。CbuATX2 蛋白的同源序列较少,只检索到与其他属的马尾草()和拟南芥()相似性较高,CbuATX2 蛋白与其他物种相同基序有KLGVD,QCKVYW,WPLDA,QFFGTHDFAR,SFLKGLL,EEAKMYL等(见图6B)。
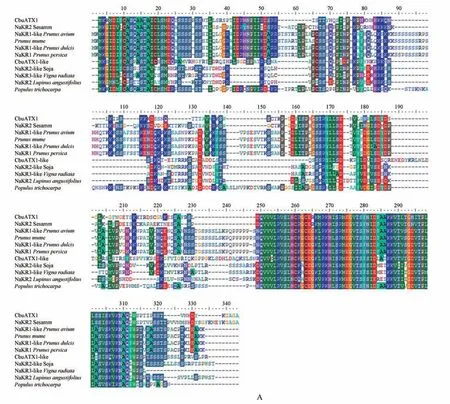
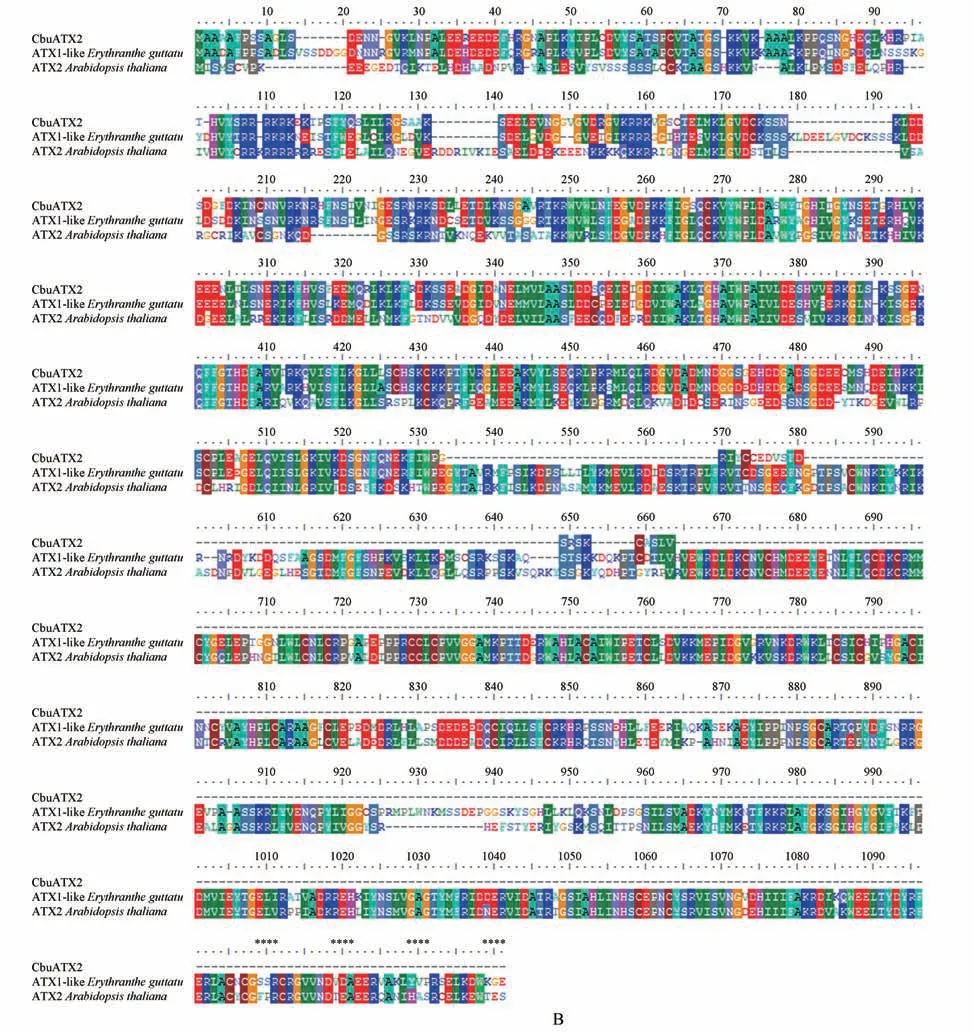
图6 CbuATX1,CbuATX1-like和CbuATX2与其它物种同源氨基酸序列比对A.CbuATX1 and CbuATX1-like与其它物种同源氨基酸序列比对;B.CbuATX2与其它物种同源氨基酸序列比对Fig.6 CbuATX1,CbuATX1-like and CbuATX2 amino acid sequence alignment with other speciesA.CbuATX1 and CbuATX1-like amino acid sequence alignment with other species;B.CbuATX2 amino acid sequence alignment with other species
2.6 CbuATX基因系统进化树构建
在NCBI 数据库中搜索,和同源基因,利用生物学软件MEGA7.0 对 楸 树CbuATX1,CbuATX1-like 和CbuATX2 氨基酸序列进行系统进化树的构建,采用遗传距离建树法的相邻连接法,并进行1000 次的Bootstrap 校正。结果显示,CbuATX1 与芝麻()、甜菜(),CbuATX1-like 与胡萝卜()、欧洲橄榄()、三叶通菜(),CbuATX2 与马尾草()的同源序列处于同一个进化分支上,表明它们的亲缘关系最近(见图7)。
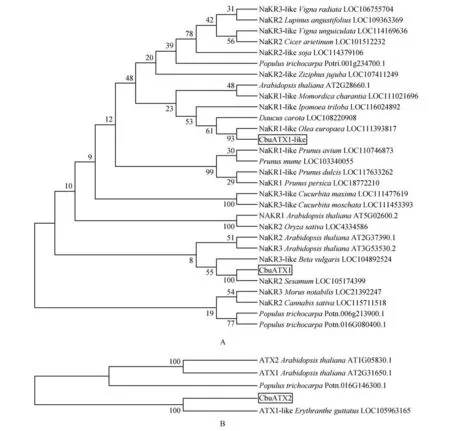
图7 CbuATX1,CbuATX1-like和CbuATX2与其它物种同源蛋白的系统进化树分析A.CbuATX1 and CbuATX1-like与其它物种同源蛋白的系统进化树分析;B.CbuATX2与其它物种同源蛋白的系统进化树分析Fig.7 CbuATX1,CbuATX1-like and CbuATX2 Phylogenetic tree analysis of proteins homologous to other speciesA.CbuATX1 and CbuATX1-like Phylogenetic tree analysis of proteins homologous to other species;B.CbuATX2 Phylogenetic tree analysis of proteins homologous to other species
2.7 楸树CbuATX1,CbuATX1-like和CbuATX2功能预测
为 了 研 究 楸 树CbuATX1,CbuATX1-like 和CbuATX2 蛋白的功能,分别利用ProtScale 在线软件预测蛋白疏水性;利用TMpred 在线软件预测蛋白跨膜结构域;利用SignalP-3.0 在线软件预测蛋白质信号肽;利用NetPhos 在线软件预测蛋白磷酸化位点。结果显示:
CbuATX1蛋白其多肽链中疏水性最强的在16区域,高达1.844,疏水性最小的在70 区域,低至-3.533,与理化性质分析的基本一致,推测是一种亲水性蛋白;CbuATX1 蛋白从内到外共有1 个跨膜区域在6~24 aa,推测是一种跨膜蛋白;CbuATX1 蛋白信号肽S 值在1~15 位氨基酸之间,其中最大值在第7个氨基酸,预测值是0.532,平均分值为0.280(小于阈值0.5),说明该蛋白不含有信号肽,推测可能是一种非分泌性蛋白;CbuATX1蛋白磷酸化位点有41个,包括Ser(丝氯酸)30个、Thr(苏氨酸)10个、Tyr(酪氨酸)1个,蛋白磷酸化位点上不存在特定修饰激酶,说明氨基酸序列上的磷酸化位点含有多种修饰类型。
CbuATX1-like 蛋白其多肽链中疏水性最强的在232 区域,高达1.344,疏水性最小的在112 区域,低至-3.522 与理化性质分析的基本一致,推测是一种亲水性蛋白;CbuATX1-like 蛋白从内到外共有1 个跨膜区域在2~18 aa,从外到内有1 个跨膜区域,在1~18 aa,可能是一种跨膜蛋白;CbuATX1-like 蛋白信号肽S 值在1~15 位氨基酸之间,其中最大值在第14个氨基酸,预测值是0.290,平均分值为0.173(小于阈值0.5),说明该蛋白不含有信号肽,推测可能是一种非分泌性蛋白;CbuATX1-like 蛋白磷酸化位点有60 个,包括Ser(丝氯酸)46 个、Thr(苏氨酸)8 个、Tyr(酪氨酸)6个,蛋白磷酸化位点上不存在特定修饰激酶,说明氨基酸序列上的磷酸化位点含有多种修饰类型。
CbuATX2 蛋白其多肽链中疏水性最强的在384 区域,高达1.978,疏水性最小的在99 区域,低至-3.778,与理化性质分析的基本一致,推测是一种亲水性蛋白;CbuATX2蛋白没有跨膜结构,可能是一个与非参与细胞信号传导的蛋白质;CbuATX2蛋白信号肽S值在1~4位氨基酸之间,其中最大值在第1个氨基酸,预测值是0.311,平均分值为0.145(小于阈值0.5),说明该蛋白不含有信号肽,推测可能是一种非分泌性蛋白;CbuATX2蛋白磷酸化位点有44 个,包括Ser(丝氯酸)35 个、Thr(苏氨酸)8 个、Tyr(酪氨酸)1 个,蛋白磷酸化位点上不存在特定修饰激酶,说明氨基酸序列上的磷酸化位点含有多种修饰类型。
2.8 楸树CbuATX1,CbuATX1-like 和CbuATX2基因不同发育时期样品中的表达量分析
通过实时荧光定量PCR,对,和基因在不同发育时期的相对表达量进行检测。发现这3 个基因在CK(普通楸树)、EF(百日花楸树)中营养生长期和生殖生长中的相对表达量与表达趋势存在差异。如图8A 所示,基因在普通楸树和百日花楸树中均呈现出从营养生长期转变到生殖生长期基因相对表达量下降的趋势。在百日花楸树中营养生长期表达量最高,与普通楸树营养生长期中相差9.3 倍,差异极显著;与百日花楸树生殖生长期相差5.8 倍,差异显著。在百日花楸树中生殖生长期表达量也要高于普通楸树生殖生长期,是其1.8 倍。如图8C 所示,基因相对表达量变化趋势与相同。但在百日花楸树中生殖生长期表达量要低于普通楸树生殖生长期,相差15%。
如图8B 所示,基因在普通楸树与百日花楸树中,其营养生长期转变为生殖生长期的相对表达量呈相反趋势。在普通楸树营养生长期转变为生殖生长期时,基因表达量上调为2倍;而在百日花楸树营养生长期转变为生殖生长期时基因表达量下调,但差异不大,仅为1 倍。综合以上结果我们发现,和3 个基因与普通楸树营养生长期相比,在百日花楸树的树营养生长期相表达量均上调,这暗示着基因在营养生长期表达的差异,很可能是造成楸树花期产生变化的重要原因之一。
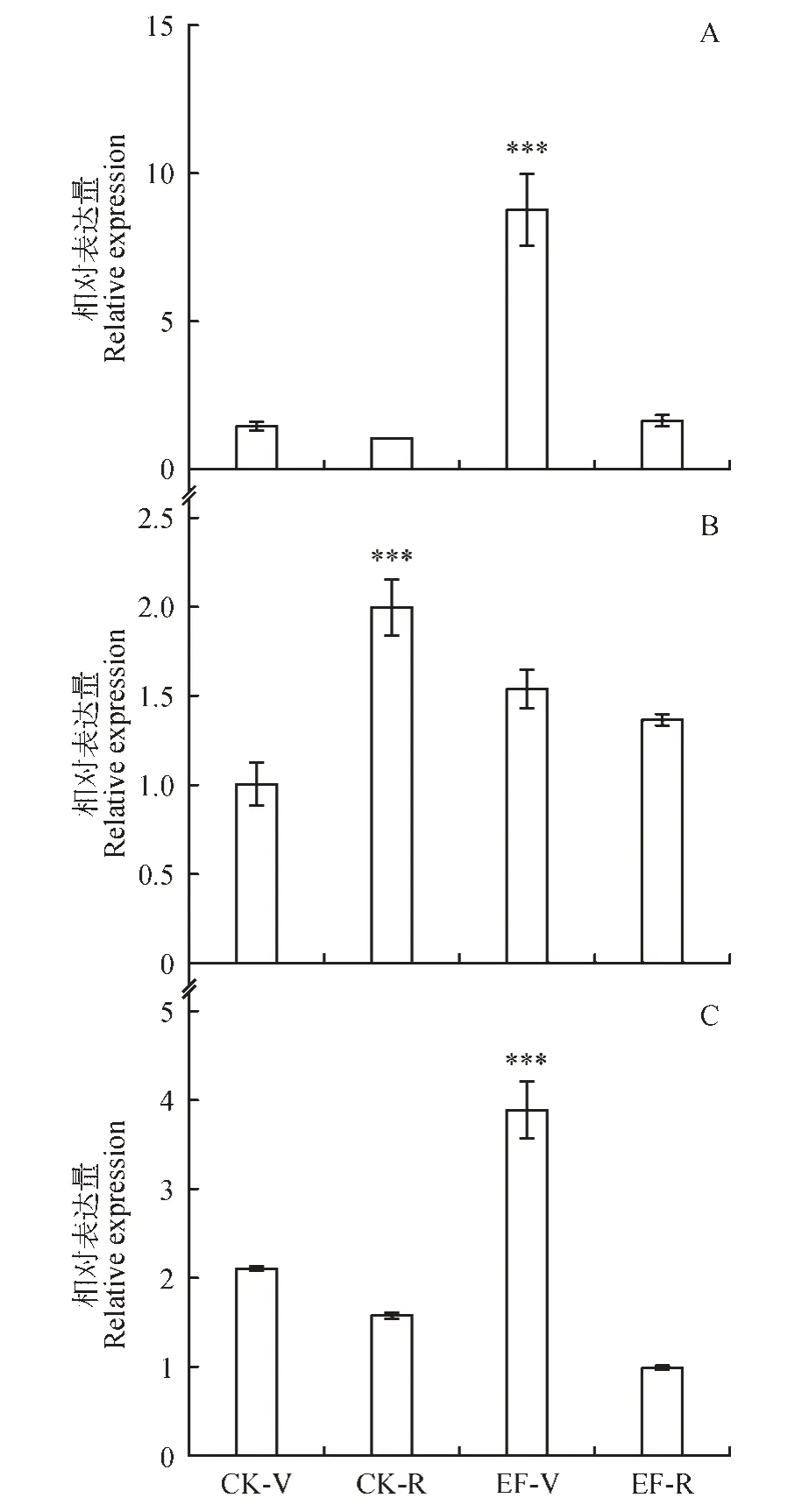
图8 不同时期不同品种A.CbuATX1 基因相对表达量;B.CbuATX1-like 基因相对表达量;C.CbuATX2 基因相对表达量;CK-V.普通楸树营养生长期;CK-R.普通楸树生殖生长期;EF-V.百日花楸树营养生长期;EF-R.百日花楸树生殖生长期,方差显著性分析;P>0.05表示差异不显著;0.01<P<0.05 表示差异性显著;P<0.01 表示差异极显著;*P<0.05,**P<0.01,***P<0.001Fig.8 Different species in different periodsA.Relative expression of CbuATX1 gene;B.Relative expression of CbuATX1-like gene;C.Relative expression of CbuATX2 gene;CK-V.Vegetative growth period of C.bungei;CK-R.Reproductive growth period of C. bungei;EF-V.Vegetative growth period of bairihua;EF-R.Reproductive growth period of bairihua;**P<0.01,**P<0.001 analysis of variance significance;P>0.05 means no significant difference;0.01<P<0.05 indicates significant difference;P<0.01 indicates extremely significant difference;*P<0.05,**P<0.01,***P<0.001
3 讨论
通过对处于不同发育时期的花芽样品进行转录组测序分析,我们从差异基因集中筛选得到差异显著的关键基因家族,通过荧光定量PCR筛选出3 个差异基因,和,并进行生物信息学分析,为探究百日花楸树开花周期奠定理论基础。本试验利用生物信息学方法对,和基因的理化性质、同源性、启动子顺式作用元件等进行分析比较。结果发现编码一个由241个氨基酸残基组成的蛋白序列,不存在信号肽,等电点为9.96,为碱性蛋白,可能具有一定的抗菌性,与芝麻、甜菜亲缘关系较近。编码一个由266个氨基酸残基组成的蛋白序列,不存在信号肽,等电点为9.80,为碱性蛋白,可能具有一定的抗菌性,与胡萝卜、欧洲橄榄亲缘关系较近。编码一个由517 个氨基酸残基组成的蛋白序列,不存在信号肽,等电点为7.21,为碱性蛋白,可能具有一定的抗菌性,与马尾草亲缘关系较近。这3 个基因含有与光响应、低温响应、生长素响应、参与防御和应激反应所必需的顺式调节元件等,说明基因很有可能与光响应密切相关并参与外界环境胁迫响应等过程。拟南芥Trithorax 1 同源基因是与根发育相关,而且动态调节拟南芥花序茎纤维次生细胞壁的生物合成的功能。本研究发现与基因相似度较高,但是否具有相同的功能还需要进一步探索。在真核生物中,PcG 和TrxG 在表观遗传调控基因表达方面具有拮抗作用,TrxG 蛋白在拟南芥中抵消了PcG 介导的花抑制,但对其在其他发育过程中的作用知之甚少,有研究报道了基因、基因和基因以及基因()在早期花发育过程中的相互作用,结果发现基因的突变未能挽救突变体的早花表型,而、和基因幼苗表现出一种新的根膨大表型。开花是相对复杂的网络调控过程,它利用许多信号通路完成,并且这些信号通路有几乎相同的下游基因,这些基因通过上游信号启动开花调控。目前比较常见的开花整合因子 主 要 有,和,究 竟,-和这3 个基因与哪个开花通路相关还有待进一步研究。这3个基因在不同发育时期表达具有一定的差异,和这两个基因具有相同的表达趋势,无论在普通楸树还是百日花楸树中从营养生长期到生殖生长期的转变都呈下降趋势,且与表达趋势相反,与基因相似度高,表达趋势却相反,3 个基因间是否存在协同与抑制还需要进一步的研究探讨。
