核桃JrNPR1基因的克隆与抗病响应潜力
2022-03-02马凯恒何佳凝谢牧洪黄倩雪杨桂燕
马凯恒 何佳凝 谢牧洪 任 斌 黄倩雪 杨桂燕*
(1. 西北农林科技大学林学院陕西省核桃工程技术研究中心,杨凌 712100;2. 西北农林科技大学林学院陕西省经济植物资源开发利用重点实验室,杨凌 712100)
植物抗病基因在识别病原菌和抗病信号传导的病害防御过程中起重要作用。近年,利用转座子标签技术、定位克隆和抗病基因同源序列法等从植物中克隆到大量的抗病基因。这些基因的克隆、鉴定和功能分析对于深入研究植物抗病机制、通过基因工程提高植物抗病性具有十分重要的意义。(nonexpressor of pathogenesis-related genes 1)基因是植物防御信号转导途径的重要调控因子,是植物水杨酸(Salicylic acid,SA)信号通路的重要组成部分,参与了SA 与茉莉酸(Jasmonic acid,JA)∕乙烯(Ethylene,ET)信号途径的相互作用;因此是植物响应病原菌侵染的核心因子。典型的基因含有BTB和ANK-r结构域,这与NPR1 同其他转录因子互作密切相关。如,与bZIP 转录因子中的TGA 家族之间的互作,通过BTB 结构域与TGA2 的抑制区结合使TGA2 抑制病程相关基因(Pathogenesis-related gene,PR)表达的功能缺失。目前,很多植物中的基因被陆续克隆,并对其功能进行了研究,发现来自不同植物的同源基因或相同植株的不同基因,其功能具有一定差异。如,烟草()具有对SA 敏感的反式激活结构域,在水稻()中过表达基因会自发激活防御基因的表达,在水稻和草莓()中防御反应的激活可能会对植物生长发育产生负面影响,影响作物生产相关的重要过程,但在拟南芥()和许多其它植物中不存在这种现象。在大量植物中,过表达及其同源基因被证实能够增强植物的抗病性,说明了1在抗病中的保守作用。可见,对未被报道的基因进行研究,一方面可以探究基因应对植株病害的功能,另一方面又可能挖掘基因的新功能。
核桃()属胡桃科(Juglandaceae)胡桃属()多年生落叶乔木,享有“世界四大坚果”之一的美誉,也是我国主要经济树种之一。随着核桃栽培面积不断扩大,核桃产业链逐渐延长,为区域经济发展做出了重要贡献。但近年,核桃病虫害问题日益突出,对核桃生长及产量产生了严重的制约,尤其是核桃炭疽病、枝枯病和黑斑病对核桃植株的危害较大,严重时可能导致绝收,甚至树木死亡。当前,防治核桃此类病害主要依靠喷洒化学杀菌剂。化学农药存在残留问题,对食品安全、环境保护等都会造成不良影响,这急切需要掌握核桃的病害调控机制,进而为寻求高效安全的病害防治措施提供依据。因此,本研究从核桃中鉴定获得1条基因(命名),通过基本生物信息及病害响应表达分析其生物学功能,为揭示核桃抗病响应机制提供候选基因。
1 材料与方法
1.1 材料及处理
植物材料:2年生‘香玲’核桃嫁接苗。
病原:①胶孢炭疽菌()(简写为CgTJ):引起核桃炭疽病的病原菌。②矩圆黑盘孢菌()(简写为MoZK):引起核桃枝枯病的病原菌。③黄单胞菌()(简写为XcHB):引起核桃细菌性黑斑病的病原菌。
处理:分别制备浓度为1×10个·L的胶孢炭疽 菌 分 生 孢 子 菌 液,配 置100 μmol·L水 杨 酸(SA)溶液。对植株分别进行SA、CgTJ、CgTJ+SA、MoZK、MoZK+SA、XcHB、XcHB+SA 喷洒处理。以喷水处理为对照。在处理后第0、1、6、9、12 d 等时间分别取叶和茎,用液氮速冻后保存于-80 ℃冰箱备用。每个处理包含6棵植株。
1.2 JrNPR1基因的克隆与分析
以“nonexpressor of pathogenesis related genes 1”为关键词在‘香玲’核桃转录组中筛选基因,通过BLAST 分析对比后,选择其中1 条基因(命名为)进行后续分析。用ORF finder(http:∕∕www.ncbi.nlm.nih.gov∕gorf∕gorf.Html)确 定基因开放读码框(ORF),通过ExPASy(http:∕∕web.expasy.org∕protparam∕)分 析基因的序列特征。采用CD-Search(https:∕∕www.ncbi.nlm.nih.gov∕Structure∕cdd∕wrpsb.cgi)对的保守结构域进行分析。利用BLASTP(http:∕∕blast.ncbi.nlm.nih.gov∕Blast)对同源性蛋白序列进行搜索。利用MEGA7 构建系统进化树。使用NEW PLACE(https:∕∕www.dna.affrc.go.jp∕PLACE∕)分析启动子顺式作用元件。
1.3 JrNPR1基因的表达分析
样品总RNA 采用十六烷基三甲基溴化铵(CTAB)方法提取。RNA 反转录cDNA 使用Prime-ScriptRT Reagent Kit(CWBIO,康为世纪,中国)试剂盒进行。cDNA 稀释10 倍用作实时荧光定量PCR(qRT-PCR)模板。qRT-PCR 反应体系参照SYBR Green Real time PCR Master mix(CWBIO)制备。定量反应仪器为Step OneReal-Time PCR System(Applied Biosystems 生产)。反应程序为:94 ℃预变性30 s;94 ℃变性12 s,60 ℃退火45 s,72 ℃延伸45 s,45个循环;81 ℃读板1 s,重复3次。基因定量引物为:5′-CCGATGTATGTTGGCTGT-3′∕5′-GGTCTACATGCATCATGG-3′。 内参基因为18S rRNA(HE574850)基因,引物为:5′-GGTCAATCTTCTCGTTCCCTT-3′∕5′-TCGCATTTCGCTACGTTCTT-3′。相对表达结果用2法进行分析。数据使用SPSS 软件包(SPSS,Chicago,Illinois,USA)处理。样品变异性用标准偏差表示。不同时间点与0 d 之间的表达差异用检验分析(<0.05)。
2 结果与分析
2.1 JrNPR1全长cDNA序列基本信息
通过查找核桃转录组数据获得1 条基因,经分析发现该基因ORF 长1 782 bp(GeneBank登录号:MW202261),拟推导的蛋白含有593 个氨基酸(见图1),理论等电点为6.40。保守结构域分析发现含有NPR1-like-C 结构域(见图1),多序列比对发现该蛋白与其它物种的NPR1 蛋白相似性很高(见图2),表明属于NPR1-like基因家族成员。构建进化树分析发现JrNPR1 蛋白与杨梅()、欧洲栓皮栎()以及加州白栎()等的进化关系较近(见图3)。

图1 JrNPR1蛋白的NPR结构域分布Fig.1 The NPR domain distribution of JrNPR1 protein

图2 JrNPR1蛋白与其同源蛋白的NPR结构域比对Jr.核桃;Mr.杨梅;Rc.蓖麻;Pd.美洲黑杨;Ss.密花豆Fig.2 Multiple sequence alignment of NPR domains from JrNPR1 and its homologous proteinsJr.Juglans regia;Mr.Morella rubra;Rc.Ricinus communis;Pd.Populus deltoids;Ss.Spatholobus suberectus
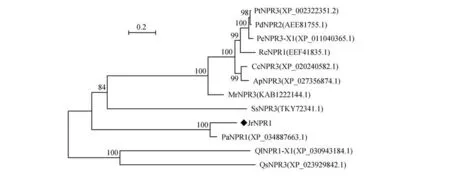
图3 JrNPR1蛋白的进化分析Jr.核桃;Mr.杨梅;Rc.蓖麻;Pd.美洲黑杨;Ss.密花豆;Ql.加州白栎;Qs.欧洲栓皮栎;Cc.木豆;Ap.非洲相思子;Pe.胡杨;Pa.银白杨;Pt.毛果杨Fig.3 Phylogenetic tree analysis of JrNPR1 proteinJr.Juglans regia;Mr.Morella rubra;Rc.Ricinus communis;Pd.Populus deltoids;Ss.Spatholobus suberectus;Ql.Quercus lobata;Qs.Quercus suber;Cc.Cajanus cajan;Ap.Abrus precatorius;Pe.Populus euphratica;Pa.Populus alba;Pt.Populus trichocarpa
2.2 JrNPR1启动子元件特征
经BLASTX 比对发现与GeneBank 中的XP_018838913.1 序列一致,以此鉴定其上游1 233 bp 的DNA 序列设计引物,以‘香玲’核桃DNA 为模板扩增获得该基因启动子序列进行生物信息学分析,NEW PLACE 预测显示,启动子中含有多种与抗病性相关顺式作用元件,如WRKY 转录因子识别的WRKY71OS 元件,RAV1转录因子识别的RAV1AAT元件,组织特异表达的CAATBOX1 元件(见表1)。推测其具有调控参与抗病的能力。

表1 JrNPR1启动子正向包含的主要顺式作用元件Table 1 The cis-acting elements contained in JrNPR1 promoter
2.3 JrNPR1基因响应不同病害的表达
对试材分别进行SA、CgTJ、CgTJ+SA、MoZK、MoZK+SA、XcHB、XcHB+SA 等处理,分析处理后第0、1、6、9、12 d 时叶和茎中基因的表达(见图4~5)。结果发现,在叶中炭疽病胁迫下,胁迫1 d 时的表达达到最大值,为对照的9.96 倍,之后的表达逐渐降低,与SA 处理下的变化趋势一致。在进行CgTJ+SA处理后,基因随胁迫后时间延长的变化趋势与单独进行SA和CgTJ处理的变化趋势一致,但其表达水平高于单一处理下的表达,在0~12 d,分别为单一SA和CgTJ 处 理 下 的1.15~3.78、1.31~7.66 倍。在MoZK、MoZK+SA 胁迫下,基因的表达变化与SA、CgTJ、CgTJ+SA 相似,在1 d 的表达值最大,分别为对照的7.00、12.41 倍。XcHB、XcHB+SA 处理下的表达最大值出现在6 d,为对照的7.91、17.86 倍。在SA 存在情况下,基因响应XcHB 表 达 是 单 独XcHB 处 理 的2.21~178.89 倍(见图4)。
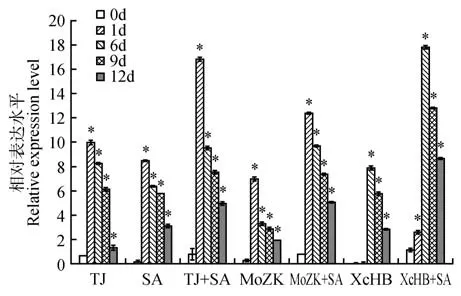
图4 不同处理下JrNPR1基因在叶中的表达*表示相同处理下不同时间点与0 d之间的差异显著(P<0.05),下同Fig.4 The expression of JrNPR1 gene in leaf under different treatments*Indicates that the difference between different time points and 0 d under the same treatment is significant(P<0.05),the same as below
在茎中,基因在SA、CgTJ、CgTJ+SA、XcHB、XcHB+SA 处理下随时间的变化趋势相似,都是在6 d 达到最大值,分别为对照的12.06、13.62、20.19、8.10、17.06 倍。在MoZK、MoZK+SA处理下的最大表达值在9 d,分别为对照的9.40、12.41 倍。SA 存在情况下,基因响应CgTJ、MoZK、XcHB 的转录水平均高于对应的单独处理下的表达,即,CgTJ+SA、MoZK+SA、XcHB+SA处理下的表达分别为CgTJ、MoZK、XcHB 处理下转录水平的1.67~12.70、1.32~2.07、1.53~3.59倍(见图5)。
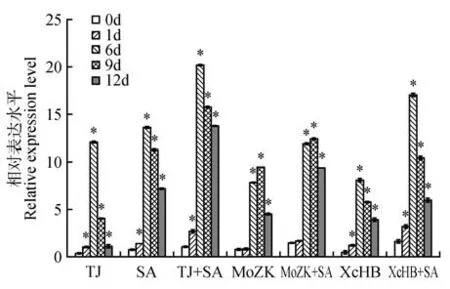
图5 不同处理下JrNPR1基因在茎中的表达Fig.5 The expression of JrNPR1 gene in stem under different treatments
3 讨论
核桃作为我国广大山区百姓脱贫致富、实现国家乡村振兴的重要木本油料树种,种植面积广泛,形成了可明显改善种植户经济收入、推动区域经济发展的重要产业链。但核桃产业的稳产、增收必须要有科学的综合管理,包括树体、营养、病虫危害防控等各个方面。特别是近年时常爆发的病虫害,值得关注。而这就要求必须掌握核桃主要病虫害发生规律及防御机制,选育抗病优良品种。NPR1 是植物病害响应中的重要因子,随着在较多草本植物中的功能解析被报道,基因得到了较多的关注,认为其表达可以增强植株的抗病能力,是植物抗病育种的重要候选基因。但在核桃中,基因尚未被鉴定,这对揭示核桃抗病机制不利。因此,本研究中我们克隆获得一条核桃基因(命名为),通过保守结构域、多序列比对、同源进化等分析发现与其它物种的NPR 蛋白具有较高的相似性和较近的亲缘关系(见图2~3),在此基础上,探究基因响应核桃主要病害的潜力。
由于启动子包含较多的可起转录调节并可预测基因潜在功能的顺式作用元件,首先,本研究鉴定获得了基因上游1 233 bp启动子进行顺式作用元件分析。结果发现,该启动子包含WRKY、ABRE、Dof 等转录因子识别的顺式作用元件,且有组织特异性表达、病害、干旱等逆境响应相关原件(见表1)。由于其他较多启动子含有相似元件,均作为预测目标基因潜在功能的重要依据,如,海蓬子()基因启动子含有MYB、WRKY、MYC等转录因子识别的元件,且涉及病害、激素响应,后被证实与基因抵抗渗透胁迫相关。核桃基因的启动子包含Dof、MYB、MYC、WRKY 等顺式作用元件和激素响应相关的ARFAT 等元件,且受NaCl、PEG及ABA 诱导,认为是重要的逆境调控候选基因。同时,考虑到基因是病害响应重要基因,可推测基因很可能参与核桃抗病响应过程。
由于大多数基因的表达水平与植物的抗性水平密切相关。如,拟南芥基因过表达对霜霉病菌()和丁香假单胞菌()的抗性增强;将基因转入番茄(),不仅能对番茄花叶病毒产生抗性,而且对多种真菌性和细菌性病害表现出广谱抗性;在水稻中组织特异性表达基因提高了对稻纹枯病的抗性。过表达苹果()基因可增强苹果对火伤病和真菌病害的抗性,并激活一系列下游抗病相关基因的表达。在葡萄()中超表达基因能够增强对白粉病的抗性。因此,为了进一步明确的病害响应功能潜力,对核桃进行了主要病害—炭疽病、枝枯病、细菌性黑斑病—胁迫处理,分析在接种后不同时间叶和茎中的表达。结果显示,基因可被胶孢炭疽菌、矩圆黑盘孢菌、黄单胞菌诱导表达,且体现了一定的病害响应特异性和组织差异性(见图4~5)。综上可得知,可作为抵抗核桃炭疽病、枝枯病、细菌性黑斑病的重要候选基因。
SA 在植物生长发育等过程中具有重要的作用。内源SA是诱导植物产生系统获得抗性(SAR)的信号物质,可调节植物体内多种生理代谢。外源SA 可诱导植株产生生物与非生物胁迫耐受性,其中病害抗性更为突出。如,外源喷施SA 和吸附SA 插签两种给药方式均能诱导烟草产生对烟草番茄斑萎病、黑胫病、普通花叶病、野火病和赤星病的抗性。在采收前对葡萄果穗喷施SA,可显著降低果实生长期间潜伏真菌率,提高红地球葡萄对潜伏侵染病害的抗性,抑制采后灰霉病效果明 显。因 此,对 核 桃 进 行 了SA、CgTJ+SA、MoZK+SA、XcHB+SA 处理,结果发现,基因的表达可被SA 显著诱导,且基因在CgTJ+SA、MoZK+SA、XcHB+SA 处理下的表达水平要明显高于单一的CgTJ、MoZK、XcHB 处理下的表达(见图4~5)。这与其他基因响应激素和逆境表达推测的功能潜力一致。如,核桃MYB 转录因子在干旱、SA 等处理下的表达认为可作为核桃逆境响应机制研究及抗逆育种的优良候选基因。番茄、、基因在SA 存在情况下,NaCl诱导的叶中表达被显著增强。可见,SA是介导基因响应核桃炭疽病、枝枯病、细菌性黑斑病的重要因子,基因是核桃抗病的重要候选基因。在后续研究中,将通过在木本和草本植株中过量表达来全面解析的抗病功能。
