入侵植物黄花草木樨的生殖构件性状及种子萌发特性研究
2022-02-24兰世超杨佳琴刘国伟胡华林
钟 娟, 兰世超, 杨佳琴, 刘国伟, 胡华林, 周 兵
(1.贵州师范学院, 贵阳 550018; 2.井冈山大学, 江西 吉安 343009;3.贵州省中国科学院天然产物化学重点实验室, 贵阳 550014)
外来植物的入侵常遵循“十数定律”,最终只有1‰的外来植物成为入侵植物。成功的入侵植物常具有较强的适应性、繁殖力和散布力[1]。繁殖力的大小常与入侵植物的入侵性成正比。种子是植物体重要的生殖构件之一,决定了大部分靠种子繁衍后代的植物的繁殖力大小[2]。大量种子的产生及较高的种子萌发率为入侵植物获得更多的后代增加机会,从而有利于入侵植物的扩散和定殖[3]。因此,探讨入侵植物生殖构件和种子萌发特性对揭示入侵植物的入侵机制有着重要意义。
黄花草木樨(Melilotusofficinalis)为豆科(Leguminosae)草木樨属(Melilotus)一年生或二年生草本植物,原产西亚至南欧一带,现广泛入我国,主要生长在山坡、河岸、路旁、沙质草地及林缘等区域,对入侵地生物多样性造成严重威胁[4]。黄花草木樨最初作为牧草在桔园、草场等地广泛种植[5],并具有较高的药用价值[6-7]。有研究表明,黄花草木樨对其他植物具有较强的化感抑制作用[8-9],对盐碱土具有较好的改善作用[10],环境适应能力较强[11]。这些特性增强了黄花草木樨的竞争力,为其成功入侵创造了便利条件。而关于黄花草木樨生殖构件特性、对入侵地生物多样性的影响、种子休眠及对沙生环境的适应性等报道较少。为此,从不同株密度下生殖构件性状、种子休眠特性、不同贮藏温度和沙埋深度下种子萌发特性等方面对黄花草木樨适应性进行了研究,以期为揭示黄花草木樨的入侵机制及其防治提供重要理论依据。
1 材料与方法
1.1 生殖构件特性测定
1.1.1样地概况
样地设在江西省吉安市吉州区吉州大道路边荒地(27°06′55″N,114°58′05″E),该地属赣江中游,罗霄山脉中段。气候温和、光照充足、雨量充沛,年平均气温为16~17.1 ℃,年平均降水量为1 487 mm。样地中黄花草木樨共生植物主要包括狗牙根(Cynodondactylon)、鸡眼草(Kummerowiastriata)、续断菊(Sonchusasper)等。样地土壤理化性质为:pH值为7.84(水土比=2.5∶1),有机质为8.74 g/kg,总氮为0.060%,碱解氮为47.40 mg/kg,速效磷为12.46 mg/kg,速效钾为92.90 mg/kg。
1.1.2生殖构件性状测量
于黄花草木樨种子成熟期,在全面踏查的基础上,选择人为干扰较小且不同密度(高48株/m2、中24.89株/m2、低13.56株/m2)的黄花草木樨种群分布地段,各设立3个样方,共9个样方,样方面积为1 m×1 m。调查每个样方内黄花草木樨数量,并在每个样方内选择生长较一致的完整植株10株,9个样方共90株。测定不同密度下黄花草木樨单株分枝数、单枝种子量、总种子量、百粒重等性状指标。
1.2 种子休眠特性测定
在室温储藏条件下,连续5年,每个月选取50粒籽粒饱满、大小一致的种子,用1%的NaClO消毒10 min,后用蒸馏水冲洗干净,将种子置于铺有一张滤纸的直径为9 cm的玻璃培养皿中,用移液管移取8 mL蒸馏水到培养皿中,然后置于人工光照培养箱内培养,培养条件为(25±1)℃恒温,光照12 h,黑暗12 h,湿度70%, 之后每天观察记录种子发芽数,以胚根突破种皮1 mm作为种子发芽的标志。设置4个重复,每皿为1个重复。
1.3 不同贮藏温度下种子萌发特性测定
将采集的种子分别进行常温、冷藏(4 ℃)、冷冻(-20 ℃)储藏,于翌年春季进行种子萌发试验。每个处理50粒种子,4个重复。种子萌发试验同1.2。
1.4 不同沙埋深度下种子萌发特性测定
1.4.1河沙处理
将河沙分别过孔径0.25 mm和1.00 mm筛子,取粒径为0.25~1.00 mm的沙子,用自来水洗净之后再用蒸馏水润洗,室温下晾干备用。
1.4.2种子萌发的测定
挑选籽粒饱满、大小一致的种子若干,用1%的NaClO消毒10 min之后,用蒸馏水冲洗干净,均匀撒播于装有河沙(厚度2 cm)的具孔塑料方盆(20 cm×10 cm×5 cm)中,每盆50粒种子。再分别以0(无覆盖)、0.50、1.00、1.50 cm厚的河沙覆盖。播种后浇水,浇水量以播种方盆底部渗出水为标准,并用塑料薄膜将盆口封好,防止水分蒸发。然后置于人工光照培养箱内培养,培养条件同1.2。之后每天统计种子萌发出土情况。设置4个重复,每1方盘视为1重复。
1.5 数据统计分析
采用Excel软件和SPSS 19.0软件进行相关数据统计分析,采用Duncan检验法进行差异显著性分析。采用Pearson指数法分析生殖构件性状间的相关性。
2 结果与分析
2.1 不同密度下黄花草木樨生殖构件性状
由表1可见,密度对黄花草木樨的生殖构件性状产生较大影响,基本表现为生殖构件性状随着密度升高而下降的趋势。在中密度和低密度下,黄花草木樨单株的分枝数、单枝种子量、总种子量以及百粒重间没有显著差异,但显著高于高密度下的相应指标(p<0.05)。尤其是在单株种子量方面,低密度下单株种子量是高密度下的1.72倍,然而其植株密度只有后者的1/2,高密度样方将产生更多的种子。相关性分析表明,除单株种子数量与百粒重呈显著正相关外(p<0.05),其他指标间均为极显著正相关(p<0.01)。
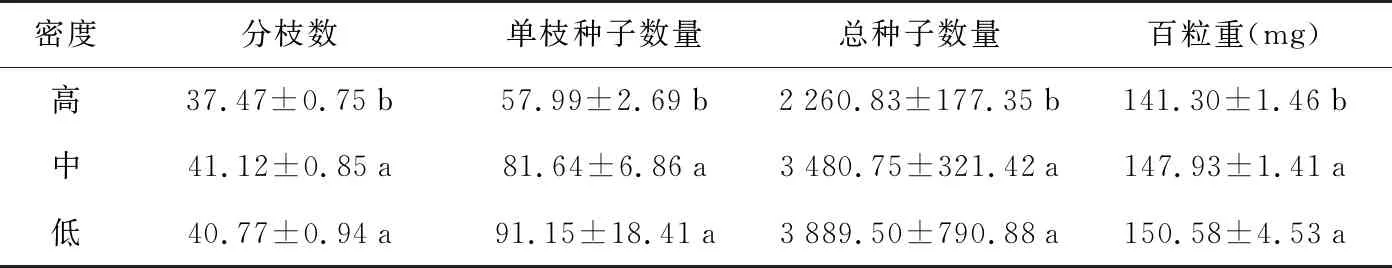
表1 不同密度下黄花草木樨生殖构件性状
由表2可以看出,单株分枝数与单枝种子数量、总种子数量、百粒重呈极显著正相关(p<0.01),且与总种子数量的相关系数最大,为0.599。单株种子数量与总种子数量、单株分枝数呈极显著正相关(p<0.01),且与总种子数量的相关系数最大,为0.946;与百粒重呈显著正相关(p<0.05)。总种子数量与其他生殖构件性状间也呈极显著正相关(p<0.01),其中总种子数量与百粒重间相关系数最小,为0.195。

表2 黄花草木樨生殖构件性状的相关性
2.2 黄花草木樨种子休眠特性
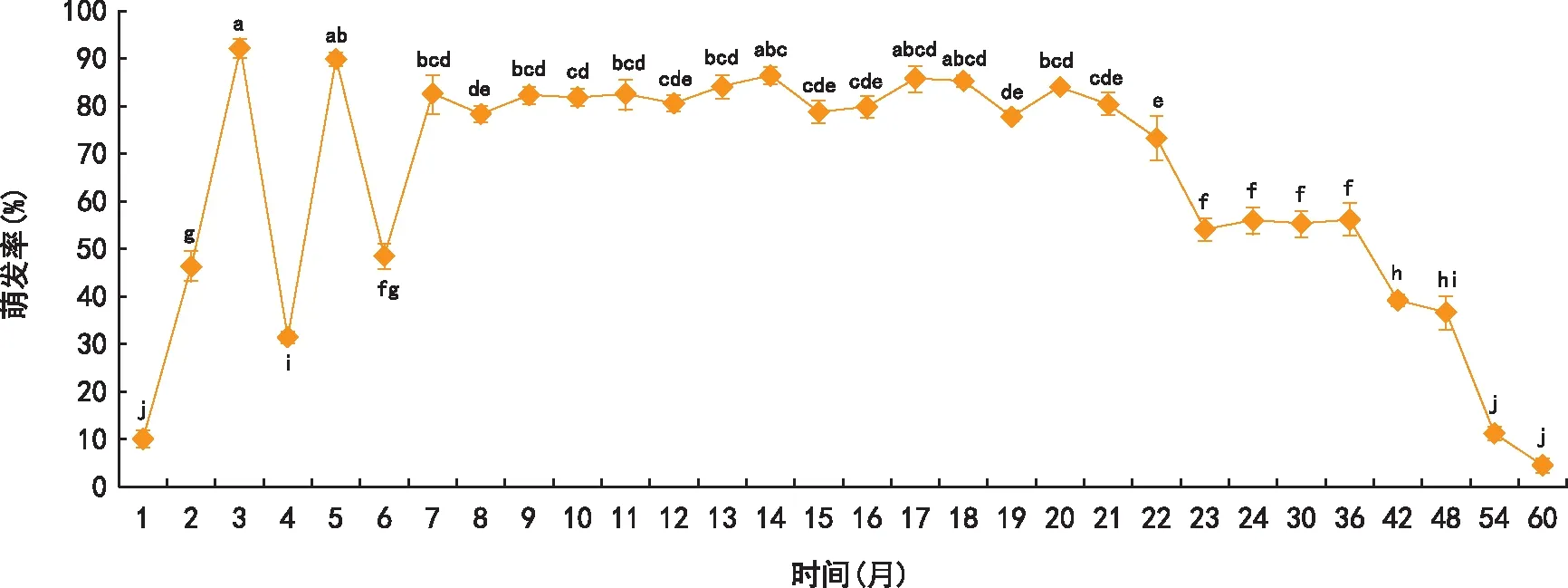
由图1可见,在黄花草木樨种子收获后的前6个月内其种子的萌发率很不稳定,而第7个月后,保持较高且较稳定的萌发高峰,其萌发率保持在80%左右直至第21个月。之后种子萌发率逐渐下降,第48个月时,其萌发率为36.75%,到第60个月时,其萌发率仅为4.50%。
注:不同小写字母表示差异显著(p<0.05)。下同。
2.3 不同储藏温度对黄花草木樨种子萌发的影响
不同储藏温度对黄花草木樨种子萌发产生了一定的影响(图2)。与室温储藏相比,冷藏可促进黄花草木樨的种子萌发,其最终萌发率为室温的1.13倍,而冷冻则对其种子萌发产生抑制作用,其最终萌发率为室温的70.39%。
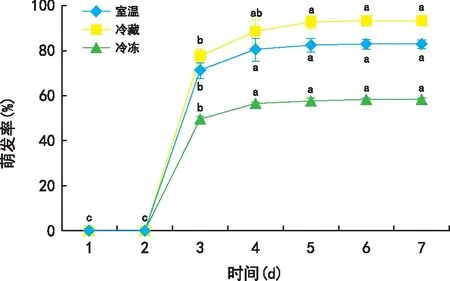
图2 不同贮藏温度对黄花草木樨种子萌发的影响
2.4 不同沙埋深度对黄花草木樨种子萌发出苗的影响
沙埋对黄花草木樨种子的萌发出苗产生一定的影响(图3)。沙埋降低了黄花草木樨的萌发出苗率,随着播种深度的增加,其最终出苗率有所降低,但各处理间没有显著差异(p>0.05)。另一方面,沙埋延缓了黄花草木樨种子的出苗速度,在播种深度为0 cm时,第4天即达到出苗高峰,而不同沙埋处理下第6天才出苗,且出苗不整齐。

图3 不同沙埋深度对黄花草木樨种子萌发出苗的影响
3 讨 论
3.1 种群密度对黄花草木犀生殖构件性状的影响
植物之间的竞争是植物获得生存空间和资源的主要方式,植物能够通过形成最佳的资源分配格局,调整自身的繁殖对策适应周围生存环境的变化,提高植物的适合度,最终保持物种在不同生境中能够得以持续生存和繁衍[12]。一年生植物通过营养生长与繁殖生长的快速转化实现最大的繁殖产量。繁殖分配是指植物在生长过程中用于繁殖结构的资源比例, 它不仅与环境有关,还受到种群密度的影响[13]。种群密度的变化影响植物对资源的竞争能力,植物通过调整生殖构件性状适应环境的变化来完成其繁衍。种群密度对黄花草木犀生殖构件性状的影响存在差异,黄花草木樨的单枝种子数量,总种子数量和百粒重与种群密度成反比,而单株分枝数在中密度下达到最大,高密度和低密度的分枝数均比中密度下的小。在中密度和低密度下,黄花草木樨单枝种子数量、总种子数量以及百粒重间没有显著差异,但显著高于高密度下的相应指标(p<0.05)。可见,中密度更适宜黄花草木犀的繁殖入侵。
据报道,许多入侵植物均能产生大量有效的种子[14-15],比如美洲商陆(Phytolaccaamericana)也具有极高的种子产量,每花序结种子200~500粒,每株结实量达1 000~10 000粒[16];苋科入侵植物反枝苋(Amaranthusretroflexus)、鲍氏苋(Amaranthuspowellii)及北美苋(Amaranthusblitoides)每株能产种子10 000~20 000粒[17]。另一方面,很多入侵植物常具有60%以上种子萌发率[18-20]。大量种子的产生及较高种子萌发率为入侵植物获得更多的后代增加机会,从而有利于入侵植物的扩散和定殖。黄花草木犀单株可产生种子2 000~4 000粒,随着储藏时间的延长,其种子萌发率于7个月后趋于稳定,保持在80%左右,21个月后其萌发率开始下降,第3、4、5年,这3年之间种子萌发率差异显著。从第4年到第5年,萌发率曲线呈直线下降趋势,直至48个月后种子萌发率极低。滇重楼(parispolyphylla)、非洲狗尾草(Setariaanceps)也具有相类似的种子萌发和休眠规律[21-22]。
3.2 环境条件对黄花草木樨种子萌发特性的影响
除遗传因子和采摘成熟度外,种子贮藏温度也是影响其活力和萌发的最重要的因素之一。唐洪辉等[23]研究表明,低温干燥的储藏条件会延缓种子衰老,可在很长时间内维持较高的生活力,有助于种子长期保存和萌发。本研究表明,冷藏对黄花草木樨种子的萌发有促进作用,萌发率提高了10%左右,而冷冻对黄花草木樨种子的萌发有抑制作用,萌发率降低了30%左右。本研究结果与唐洪辉等[23]对金合欢(Acaciafarnesiana)、黄黎芳等[24]对火龙果(Hylocereusundulatus)、张渊等[25]对杏(Armeniacavulgaris)的结果保持一致,低温储藏下比常温储藏下种子的萌发率更高,其原因可能是低温可以减弱种子呼吸和缓减种胚内多种活化酶转化,减少了种子内营养物质的消耗,从而延长了种子寿命,提高萌芽率。
沙埋是许多沙生环境中植物需要适应的环境条件之一,沙埋对植物种子萌发、幼苗出土、生长和繁殖等方面产生影响[26-27]。Grundy等[28]认为,所有植物最大的种子萌发出苗率应该在土壤表面,其种子萌发出苗率随着沙埋深度增加而降低。Benvenuti等[29]对钝叶酸模(Rumexobtusifolius)、周兵等[30]对三叶鬼针草(Bidenspilosa)的研究结果支持了这一结论。但Forcella等[31]认为,一些植物的种子萌发出苗率与沙埋深度呈现抛物线型关系,一定深度的沙埋将有利于其出苗。杨慧玲等[32]对柠条锦鸡儿(Caraganakorshinskii)、Ren等[33]对蓼科(Polygonaceae)沙拐枣属(Calligonumsp.)植物等研究结果与之一致。然而,无论是哪种观点,由于过度沙埋极大的增加了覆盖物对植物幼苗的压力,严重降低了叶片的光合作用效率,最终导致植物种子萌发出苗率下降和幼苗生长受到抑制[26]。本研究结果表明,随着沙埋深度的增加,黄花草木樨种子萌发出苗率逐渐下降,这一结果从一定程度上支持了Grundy等[28]的结论,而另一方面,各处理的最终萌发出苗率差异显著(p>0.05),这也表明黄花草木樨对沙埋环境具有一定的适应能力。
可见,黄花草木樨具有较高的种子产量、保持长时间较高的种子萌发率以及对沙埋环境一定的适应能力,这些特性将有利于其在新生境中的定殖与扩散,有助于其入侵。
