射干种子萌发的影响因素及其对NaCl胁迫的响应
2022-02-24耿晓东吉爱伟汪成忠钱剑林
耿晓东, 吉爱伟, 周 英, 汪成忠, 钱剑林
(苏州农业职业技术学院, 江苏 苏州 215008)

图1 种子外部形态及解剖形态
射干(BelamcandachinensisL.)为鸢尾科射干属多年生草本植物。射干属植物全世界仅有2种,其中的射干产于我国东北、华北、华中、西南等22个省、市、自治区,多生长于海拔较低的林缘或山坡草地[1]。射干花朵艳丽,精致有趣,在少花的夏季盛开,是优秀的观赏资源和育种资源。射干适应性广泛,抗寒和抗旱性均强,是水土保持、防风固沙的优选材料,在我国大部分区域的城市园林内都适宜栽培。由于其根状茎具有药用价值,很多学者对其研究较多的是射干的药理成分和药效方面[2-3],少部分研究涉及射干的生物学特性、系统分类等方面[4-6],关于射干种子的相关研究较为少见。
种子萌发是植物发育的起点和适应外界环境的初始环节,也是植物生活史中最为关键和脆弱的阶段[7-8]。前期研究发现,射干在自然条件下发芽缓慢、发芽率低、出苗不整齐,这成为限制射干规模化生产、影响其种苗繁育和杂交育种的重要因素。系统探究射干种子的萌发能力和促进发芽的便捷手段,对提高射干在城市园林中的有效应用显得十分必要。射干具有很强的抗性,在坝地、风沙滩地、盐碱地都可以生长[9],但目前鲜见相关研究。由于萌发阶段的试验数据可从多角度揭示植物的逆境胁迫机制,加深对植物响应环境胁迫的理解,本研究在明晰射干种子萌发条件基础上,利用萌发期对逆境胁迫极为敏感的特性,摸清射干种子的抗盐幅度范围,以期为射干的后续研究和生产实践提供基础数据,为耐盐植物的筛选和应用扩大种质资源库。
1 材料与方法
选取栽植于苏州农业职业技术学院相城科技园基地的三年生射干植株及采收的种子作为研究对象,2017—2019年分阶段进行各项试验。
1.1 种子产量与构成因素测定
随机选取30株射干植株。统计每花序的单花数、坐果率、胚珠数、种子质量[10-11]。
1.2 种子生活力、含水量及吸水速率的测定
种子生活力采用四唑染色法进行测定[12];种子含水量采用烘干法进行测定[14];种子吸水速率的测定:选取完整的射干种子和去皮种子各30粒,3次重复。称量后放入烧杯在25 ℃恒温箱中进行吸胀,每隔1 h进行一次称重。重复此操作直到质量恒定。
种子吸水率(%)=[(吸水后质量-吸水前质量)/吸水前质量]×100%。
1.3 抑制物萌发试验
粗提物的抑制活性测定[12]:分别取射干的两个不同部位(种皮和胚乳)的粗提液进行白菜种子萌发试验。28 ℃黑暗条件下培养,24 h后测发芽率,48 h后测幼根长度,每处理重复3次,蒸馏水作为对照(ck)。白菜种子千粒重为3.41 g,含水量约70 mg/g。
1.4 种子发芽试验
大田萌发:射干种子在采收后于翌年4月播种于大田。观察90 d的发芽情况。
温度处理: 1) 不同恒温条件的处理:观察5 ℃、10 ℃、15 ℃、20 ℃、25 ℃、30 ℃恒温条件下种子发芽情况; 2) 昼夜变温处理:白天(08:00—16:00时共8 h)用高温处理,夜间(16:00—08:00时共16 h)用低温处理。经预试验后,筛选出两种变温处理方式:30 ℃(8 h)/20 ℃(16 h)和25 ℃(8 h)/15 ℃(16 h)。
机械及层积处理: 1) 新采收的种子经碾压后去除外种皮,20 ℃条件下发芽试验; 2) 射干种子采收后散置于湿度为60%的湿沙箱内,在4 ℃冰箱内保存30 d。常规发芽试验。
发芽试验方法:40 ℃温水浸种。选择沉水种子每处理30粒,3次重复。统计各项发芽指标。
发芽率(%)=(种子发芽数/供试种子总数)×100%;
发芽势(%)=(发芽高峰期发芽的种子数/供试种子总数)×100%;
相对发芽率(%)=(胁迫处理的发芽率/对照的发芽率)×100%;
发芽指数=∑Gt/Dt。
式中,Gt为第t日的发芽数,Dt为相应的发芽日数。
1.5 种子萌发对NaCl胁迫的耐受性试验
NaCl胁迫试验中设置5个NaCl浓度水平,依次为:100 mmol/L、200 mmol/L、300 mmol/L、400 mmol/L、500 mmol/L。将等量的NaCl溶液分别加入培养皿中,保证皿内滤纸全部浸湿且达到饱和状态,随机放置30粒种子。25 ℃(8 h)/15 ℃(16 h)条件下培养,3次重复。以等量的蒸馏水作为对照(ck)。
1.6 数据处理
数据统计及作图使用Excel 2016软件,差异显著性分析使用SPSS 22.0软件。
2 结果与分析
2.1 射干种子潜在产量
植物结实的最终目的是获得最大的种子产量。种子产量的高低,直接影响单株植物的繁殖效率,说明其扩大再生产的能力。观察统计30株射干植株的开花和坐果情况,如表1所示。称重后计算平均值,种子单粒重为22.91 mg,由所有数据计算得出射干的潜在种子产量为21.67 g,实际种子产量占潜在种子产量的77.8%。说明射干的实际种子产量很高,可以满足规模化育苗的需要。

表1 射干的开花数和坐果率
2.2 射干种子生活力和含水量
射干种子的含水量随着贮藏时间的延长出现大幅度下降且差异极显著(p<0.05)。新鲜种子及采收4个月、8个月、12个月后,种子含水量分别为14.6%、10.7%、9.1%及8.9%。贮藏4个月的种子含水量下降26.7%,贮藏8个月的种子含水量下降37.7%。因此采收8个月内是种子失水的高峰期。
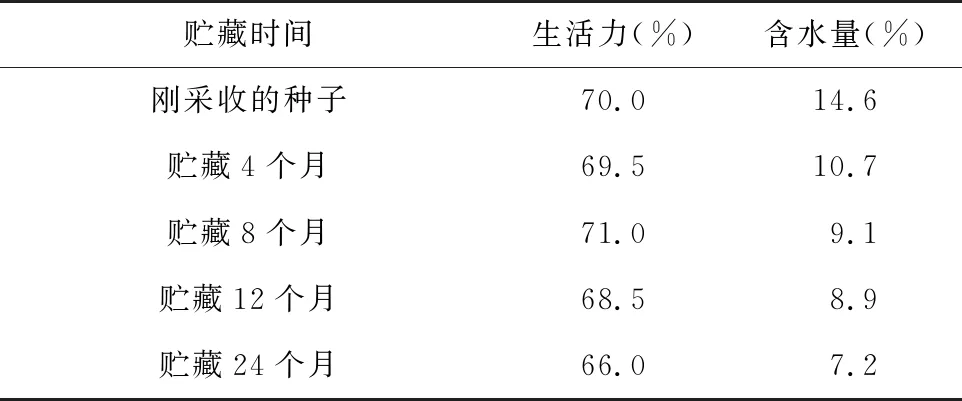
表2 不同贮藏时间的射干种子生活力和含水量
2.3 射干种子的吸水速率
从吸水试验可以看出(表3),射干的去皮种子和带皮种子吸水速率均较高,在吸胀2 h时吸水率即达到58.96%和35.22%,吸胀60 h后趋于饱和。带皮种子在吸胀第1天的吸水率达96.71%,说明射干种皮的透水性较好。

表3 射干种子的吸水率
2.4 射干抑制物萌发试验结果
抑制物萌发试验中,白菜种子在射干种皮浸提液中的发芽率降低21.48%,胚根长度在萌发48 h和72 h后仅为ck的50%左右。种皮浸提液对白菜种子的发芽和胚根生长影响显著(p<0.05)(表4)。说明射干种皮虽有较好的透水性,但其含有的某些内源物质仍抑制了种子萌发。

表4 抑制物萌芽试验的发芽情况
2.5 射干种子发芽试验结果
2.5.1大田发芽率
观察发现,射干种子在大田播种后的12~14 d陆续发芽,至30 d左右时发芽率为30.27%。其后发芽率极为缓慢,在80 d时统计大田发芽率为43.84%。说明射干种子在自然条件下的发芽率较低。
2.5.2温度对种子萌发的影响
如表5所示,射干种子在恒温条件下的发芽情况十分不理想。在较为适宜的温度条件下(25 ℃、20 ℃)发芽率也仅为28.76%和40.51%。而在两种变温条件下,种子发芽率得到显著提高(p<0.05)。发芽率分别达到84.04%和88.68%,较恒温条件的发芽率提高了2倍以上,说明变温是提高射干种子发芽率的重要手段。25 ℃/15 ℃与30 ℃/20 ℃相比,发芽率稍高,但两者之间差异不显著(p>0.05)。
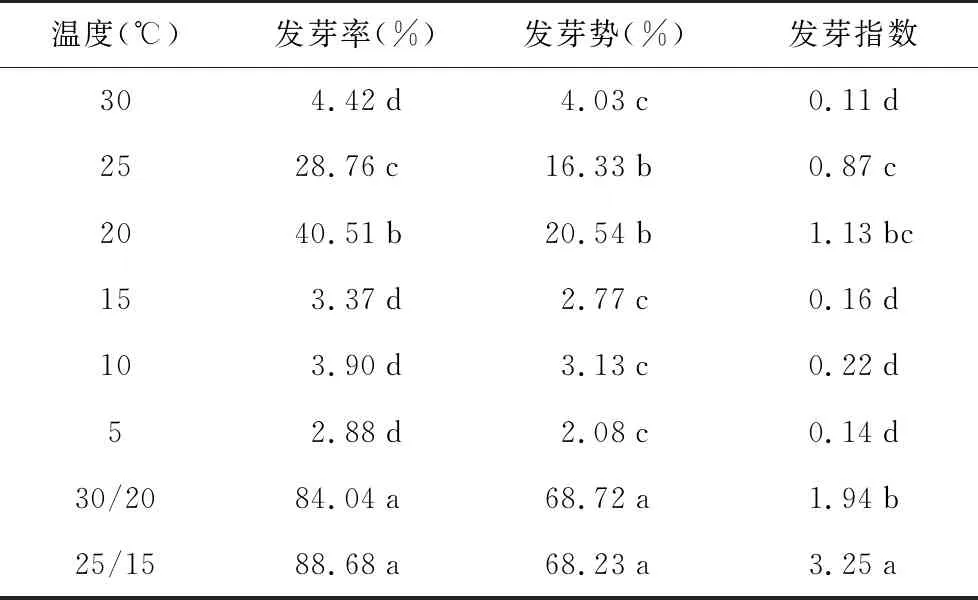
表5 不同恒温及变温条件下射干种子的发芽情况
2.5.3机械去皮及层积处理对种子萌发的影响
我不知怎么竟然摸到了一根竹竿。这根竹竿是前几天同屋里的一位同事从积木场上顺回来晾晒他的内衣内裤的,那上边还有花花答答的裤衩子。现在我要派上用场了。
如表6所示,将射干去除种皮后在20 ℃恒温条件下培养,发芽率达到50.04%,与完整种子同温处理40.51%的发芽率差异显著(p<0.05),进一步说明射干种皮抑制发芽。
经过2个月的低温沙藏,观察发现有少量射干种子已经在湿沙中发芽,经统计发芽率为1.64%,说明种子在长期的低温层积过程中已经打破休眠。低温层积后的3种温度条件发芽试验结果表明(表6),仍是25 ℃/15 ℃变温处理下发芽率最高,达到96.47%,同时这一处理下的发芽势也较非层积处理后的发芽势高,说明低温层积不但能够打破休眠,提高种子发芽率,而且对提高发芽整齐度具有重要促进作用。

表6 机械去皮及低温层积处理后射干种子的发芽情况
2.6 NaCl胁迫对射干种子萌发的影响
如表7所示,100 mmol/L NaCl胁迫下射干种子的发芽率、发芽势、相对发芽率和发芽指数均稍高于ck,但差异不显著(p>0.05)。此时的发芽率可高达90%以上,说明低浓度的NaCl胁迫可以小幅促进种子的萌发;200 mmol/L的NaCl胁迫下各项指标与ck持平,两者之间差异不显著(p>0.05);当NaCl胁迫浓度高于300 mmol/L时,各项萌发指标开始发生显著降低(p<0.05);400 mmol/L NaCl胁迫浓度时,种子发芽率仅为ck的36.4%,发芽势和发芽指数也分别为ck的38.4%和52.6%;至500 mmol/L时,种子发芽率已降低至10%左右,仅为ck的14.5%,且发芽不整齐,此时种子活力下降幅度大,严重影响了种苗的质量。

表7 不同浓度NaCl处理下射干种子的发芽情况
将种子发芽率下降到50%时所对应的NaCl胁迫浓度作为种子萌发的临界值。将种子发芽率下降到25%时所对应的NaCl胁迫浓度作为种子萌发的极限值[12]。据此发现NaCl溶液浓度和射干种子发芽率之间呈显著的负相关关系。一元线性回归方程进行拟合后效果较好,回归方程为y=-0.207 8x+120.02(R2=0.942 9)。据此预测射干种子NaCl胁迫的临界值为337 mmol/L,极限值为457 mmol/L。
3 讨 论
3.1 射干种子的发芽能力及影响因素
卡恩吸水理论[15]认为,种子吸水率能够达到38%及以上是保障其萌发的基本条件之一。射干种子的种皮质地较为疏松,吸水速率较高,在吸胀2 h的吸水率为58.69%,说明射干种皮的透水性满足其萌发需求。刚采收的新鲜射干种子的生活力为70.0%,且在较长的时间内保持稳定,说明生活力也不是限制射干种子萌发的主要因素。研究表明,多数休眠种子体内含有内源抑制物质[16-17],种子浸提液的生物测定可以有效检测抑制物是否存在[18]。本研究结果表明,射干的种皮较胚乳对白菜种子的萌发抑制作用显著。后续的20 ℃恒温条件萌发试验中,去除种皮后的种子发芽率比完整种子的发芽率提高了40.9%,表明去除种皮是提高种子发芽率的有效手段。胚乳浸提液虽对小白菜种子萌发有轻微抑制,但抑制作用不显著,说明抑制物主要为种皮,其导致射干种子的生理休眠。种皮内源抑制物质的有机成分和具体含量还需要进一步测定和验证。
温度作为种子萌发的先决条件,对植物的萌发结果影响显著[19]。本研究发现,恒温条件并不适合射干种子萌发,各梯度处理中20 ℃下最高,但萌发率也仅为35.51%。这一研究结果与曲艳等[20]得出的射干种子在20 ℃恒温条件下发芽率最高的结论有一定差异。是否因不同种源、不同年份种子造成了这种差异值得进一步探究。从另一方面来看,射干种子萌发的温度跨度很大,这与其南北均有分布的广大范围、温度差异大的生长环境是相适应的。经过10 ℃温差的昼夜变温处理后,射干种子的发芽率超过80%,较恒温条件提高了50%以上,说明温差是射干种子发芽极其重要的因素。很多研究表明,在种子发育过程中,低温能够打破新收获种子的休眠[21],昼夜变温能够促进种子萌发的速率和整齐度。有学者曾经测定85种植物种子的萌发条件,发现变温有利于68种植物种子的萌发[22]。很多相关研究均证实了这一点[23-24],本研究也与这些研究结果一致。
层积处理30 d后,射干种子在恒温条件下的发芽率得到较为显著的提升,说明层积改善了种子对温度的敏感度。同时观察发现,层积后的种子种皮表皮空隙变大,种胚体积明显增大,种胚弯曲幅度降低,种胚伸长,胚乳开始被消耗。说明层积解除了射干种子的形态休眠,这与其他一些植物的低温层积后种胚的发育阶段是一致的[25-26]。相关研究也表明,层积能够消除种子内源抑制物,诱导胚发育所需物质的形成。综合各种萌发处理方式的效果和难易程度,本研究提出适宜射干种子萌发的便捷方法为:低温层积处理30 d后置于25/15 ℃昼夜变温条件下萌发。如果进行大田播种,30 d左右的层积处理也有利于提高种子发芽势,使得出苗整齐,有利于规模化生产。
3.2 射干种子的NaCl耐受能力
射干抗性强,有一定的耐盐碱能力,本研究在种子阶段证明了这一点。结果表明,射干种子的各项萌发指标在0~200 mmol/L NaCl胁迫下变化不显著,其中100 mmol/L NaCl溶液浓度甚至轻微促进了种子萌发,这在一定程度上是Na+刺激作用的结果[27]。当胁迫浓度增加到300 mmol/L以上时,射干种子的发芽率、发芽势、发芽指数随着胁迫浓度的提高均呈显著降低趋势。当胁迫浓度增加到500 mmol/L时,射干种子的发芽能力、发芽数量已经严重下降,表现为岀苗不整齐、长势不整齐,部分胚根变质,严重影响后续的种苗正常发育。本研究根据相关方程预测射干种子NaCl胁迫的临界值和极限值分别为337 mmol/L、457 mmol/L,这一结果为射干的实生苗繁育、改良土壤、降低土壤的碱性提供数据参考。关于射干种子在不同萌发阶段和不同器官的NaCl耐受性和耐受机理还有待进一步的研究。
