农田景观格局对南疆苹果园梨小食心虫成虫种群动态的影响
2022-02-22宋博文杨龙潘云飞李海强李浩冯宏祖陆宴辉
宋博文,杨龙,潘云飞,李海强,李浩,冯宏祖,陆宴辉
农田景观格局对南疆苹果园梨小食心虫成虫种群动态的影响
宋博文1,2,杨龙2,潘云飞2,李海强2,李浩1,2,冯宏祖1,陆宴辉2
1塔里木大学农学院,新疆阿拉尔 843300;2中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室,北京 100193
梨小食心虫()是南疆苹果园一种重要害虫,严重影响苹果的产量和品质。论文旨在明确农田景观配置与组成对苹果园梨小食心虫种群数量的影响,为解析南疆地区种植业结构调整下梨小食心虫的发生趋势及灾变机制并制定种群区域治理对策提供科学依据。2017—2020年,在阿克苏地区共选择50个苹果园作为试验点,利用性诱剂诱捕监测梨小食心虫成虫种群动态,并调查试验点周围2.0 km半径范围的景观格局。在此基础上,拟合0.5、1.0、1.5、2.0 km尺度下景观格局的香农多样性指数(SHDI)、周长面积比(PARA)、边缘密度(ED)以及寄主作物、其他(非寄主)作物、非作物生境在景观中的面积占比与第1、2、3代梨小食心虫成虫数量的线性混合模型,结合赤池信息准则和模型平均,分析不同景观参数对各代成虫数量的影响。研究区域中,梨小食心虫寄主作物面积占比最高(45.7%—55.0%),其次为其他作物(18.2%—21.0%)、非作物生境(13.5%—19.7%)。模型拟合的结果显示,第1代梨小食心虫成虫种群丰富度与2.0 km景观尺度下其他作物比例负相关(=0.062)。第2代成虫种群丰富度与4个尺度下其他作物比例均呈负相关关系(0.5 km,<0.001;1.0 km,<0.001;1.5 km,=0.028;2.0 km,=0.043),与1.0、1.5 km尺度下寄主作物面积占比负相关(1.0 km,=0.026;1.5 km,=0.048),与0.5、1.0 km尺度下非作物生境比例负相关(0.5 km,=0.023;1.0 km,=0.019),与0.5、1.0 km尺度香农多样性指数(SHDI)正相关(0.5 km,<0.001;1.0 km,=0.005)。第3代成虫种群丰富度与0.5 km尺度下非作物生境比例负相关(0.5 km,<0.001)。农田景观系统中寄主作物、其他作物以及非作物生境比例的增加减少了苹果园梨小食心虫的发生量,多样性的景观格局促进了梨小食心虫在苹果园中的发生。南疆地区农业种植结构的调整显著地影响了梨小食心虫的发生,提高景观范围内其他作物、非作物生境面积占比并减少寄主作物的混栽将有利于苹果园梨小食心虫的防控。
农田景观格局;植食性昆虫;寄主植物;梨小食心虫;苹果园
0 引言
【研究意义】农田景观系统中,不同生境的组成和配置可以通过食物资源和微环境介导的“自下而上(bottom-up)”效应直接作用于害虫,亦可通过天敌昆虫“自上而下(top-down)”的控制作用间接调控害虫种群发生[1]。明确特定景观系统中不同生境的组成与配置对害虫种群发生的影响,是从景观尺度上明确农业害虫发生趋势和灾变机制,调整作物布局实现害虫区域性生态调控的关键。【前人研究进展】资源集中假说(resource concentration hypothesis)认为,在寄主作物生境比例较高的农田景观系统中害虫更容易获取连续性的食物资源,有利于害虫种群的增长[2]。例如,松褐天牛()的种群密度会随着林地面积的增大而上升[3]。但大面积的寄主作物也可能会稀释田间害虫的种群密度,Yang等在我国华北地区通过景观尺度上的研究发现小麦大面积种植的农田景观中麦田蚜虫的迁入量较低[4]。天敌假说(enemies hypothesis)认为,复杂景观系统中大量的非作物生境能为天敌昆虫提供可替代猎物或寄主、花粉、花蜜等营养物质以及庇护场所,能促进农田生态系统中天敌昆虫的发生与控害,进而减轻田间害虫的发生[5]。如华北农田景观系统中非作物生境林地和休耕地比例的增加有利于促进麦田、玉米田中捕食性天敌瓢虫种群的发生,能有效抑制害虫的种群增长[6-7]。此外,非作物生境在景观中的配置同样作用于害虫的发生,Ardanuy等发现景观边缘密度(edge density,ED)的增加更有利于天敌昆虫从田埂等非作物生境中获取资源,提高了玉米田中天敌昆虫小花蝽(spp.)的种群密度,有利于控制田间害虫的发生[8]。然而,景观中的非作物生境有时能成为害虫的庇护所,为迁入作物田中的害虫提供虫源。Santoiemma等发现,樱桃园周围的森林生境能为斑翅果蝇()提供适宜的微环境和可替代性寄主,随着周围森林面积的增加果蝇在樱桃园的种群密度和危害程度加重[9]。【本研究切入点】种植结构调整改变着害虫生存的生态环境、食物资源以及有益天敌对害虫的控制功能,常显著影响田间害虫的种群发生。南疆地区特色林果业进入了转型升级阶段[10],果树的种类和种植面积日益增加,产业结构的调整将给南疆地区果树害虫发生和种群消长带来重要影响。苹果是阿克苏地区重要的经济作物,2018年阿克苏地区苹果栽种面积达2.59万公顷,总产量63.04万吨[11]。梨小食心虫()属鳞翅目,小卷蛾科,是一种世界性的果树害虫,也是危害该地区苹果产业的重要害虫之一[12]。阿克苏地区梨小食心虫一年发生5代,越冬代4月上旬开始羽化,4月中旬达到羽化高峰,第1代6月上旬达到羽化高峰期,第2代7月中旬达到羽化高峰期,第3代8月上旬达到羽化高峰期,第4代成虫于9月上旬达到羽化高峰[13]。其中第3代幼虫对苹果危害最为严重,蛀果率可高达70%以上[14]。但产业结构调整下,大尺度范围内梨小食心虫种群发生和消长规律以及影响因素还缺乏研究。【拟解决的关键问题】以南疆阿克苏地区特有的荒漠-绿洲生态系统中的苹果园为研究对象,评估农田景观格局对苹果园中梨小食心虫成虫种群动态的影响,从景观尺度上明确梨小食心虫发生趋势和灾变机制,为通过调整作物布局实现害虫区域性生态调控提供理论依据。
1 材料与方法
1.1 梨小食心虫成虫种群动态调查
在阿克苏地区阿克苏市和温宿县,2017—2020年连续4年共选取了50个苹果园(图1),采用性诱剂诱捕的方法调查梨小食心虫成虫种群发生动态。其中,2017、2018、2019和2020年所选的果园数量分别为12、16、11和11个,诱捕的时间分别为6月19日至8月19日、7月18日至8月21日、5月10日至9月2日、6月6日至7月25日,所选果园树龄均为5—10年。每年在所选果园中随机选择5棵果树悬挂诱桶,每棵树彼此间距离不得少于20 m,且距离果园边缘至少15 m。将装有中捷四方生物科技股份有限公司提供的梨小食心虫性信息素诱芯的诱桶悬挂于距地1.5 m高度的苹果枝干上,确保诱桶四围空气流动性良好。每周收集诱捕到的梨小食心虫成虫并带回实验室统计数量,并在每周收集成虫后更换诱芯。
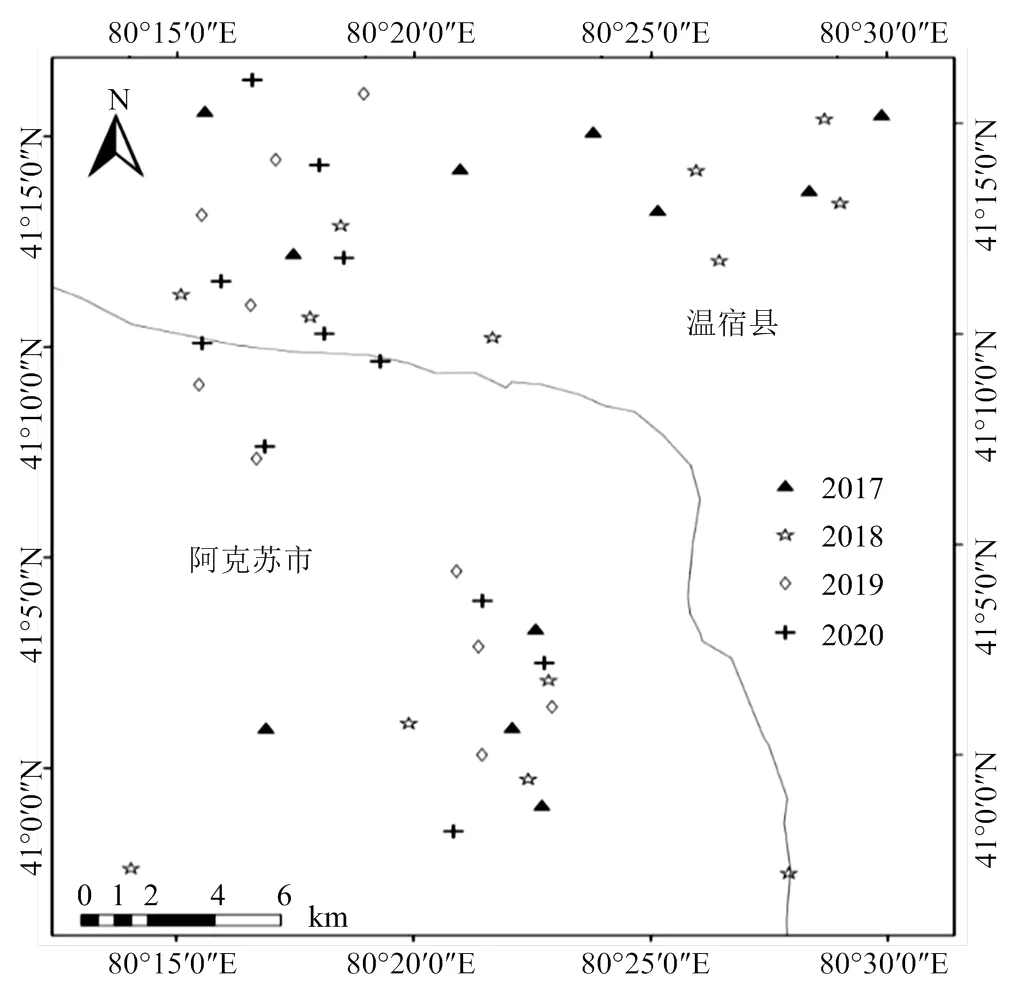
图1 2017—2020 年苹果园调查试验点
1.2 农田景观格局调查
以所选苹果园作为圆心,实地调查2.0 km半径范围内所有土地使用信息,并将各个斑块的类型标记到预先打印的纸质卫星影像地图上。结合纸质影像图上记录的斑块信息在Google earth软件中利用多边形绘图工具绘制各种生境类型的矢量图,将绘制好的矢量地图导入到Arcgis软件进行几何修复,按尺度筛选图层并转为栅格数据导出。然后,将栅格数据导入到Fragstats 4.2,分别计算出0.5、1.0、1.5和2.0 km 4个尺度下相应的景观参数。
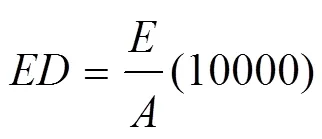

1.3 数据统计与模型拟合
根据梨小食心虫成虫种群动态及相关文献[15],将调查得到的数据划分成第1、2、3代,采用线性混合模型(linear mixed model)拟合第1—3代梨小食心虫成虫种群数量对各尺度上景观变量的响应。其中,不同代次梨小食心虫成虫数量作为因变量,各景观尺度上寄主作物、其他作物以及非作物生境面积占比、边缘密度、周长面积比、香农多样性指数作为固定效应因子,试验站点嵌套在年份里作为随机因子以排除年份和诱捕时间导致的差异。线性混合模型的全模型如下:
Full model=lmer (Grapholitha_molesta.xt~scale (Othercrops)+scale (Host crops)+scale ()+scale (Non-crop habitats) +scale ()+scale ()+(1|Year/Site))
其中,Grapholitha_molesta.xt表示某一世代梨小食心虫成虫种群数量,Non-crop habitats、Other crops、Host crops依次表示非作物生境、其他作物、寄主作物在某一尺度的面积占比,scale ()函数对自变量进行标准化。
用矫正后赤池信息准则(Bias-corrected Akaike’s information criterion,AICc)分别对不同尺度上的所有模型进行排序,AIC和AICc值计算公式如下:

式中,k为模型中参数个数,L为模型对数极大似然值,n为样本个数,RSS为误差平方和。然后,筛选出不同尺度上第1、2、3代AICc差值(AIC)小于2的竞争模型进行模型平均,解析农田景观组成与配置对不同代次梨小食心虫种群动态的影响。
数据分析使用R语言(R version 4.0.3)完成,采用bestNormalize ()函数对不同世代的梨小食心虫羽化高峰期成虫种群数量进行正态转换;用car程序包中的vif ()函数对模型进行共线性检验,发现所有模型中变量的方差膨胀因子VIF值均小于3.5,表明各变量间的共线性对模型拟合结果并无显著影响;用lme4程序包中的lmer ()函数进行线性混合模型的拟合,MuMIn程序包进行模型平均,并用DHARMa程序包对拟合后的模型残差进行正态性检验、方差齐次性检验和空间自相关检验,没有发现残差与拟合值以及各自变量间有明显的趋势,且不存在空间自相关问题。
2 结果
2.1 苹果园梨小食心虫种群动态
5月中旬到8月中旬共包含3个世代,其中5月中旬至6月中旬为第1代,6月中旬至7月中旬为第2代,7月中旬至8月中旬为第3代。苹果园梨小食心虫第1、2和3代成虫数量分别为32.76±3.74、20.27±1.71和19.02±1.85头/桶/周。其中,第1代梨小食心虫成虫发生量最高,其次是第2代成虫(图2)。
2.2 农田景观特征
景观组成在4个景观尺度表现为寄主作物面积占比最高(45.7%—55.0%),其次为其他作物(18.2%—21.0%)、非作物生境(13.5%—19.7%)(表1)。

表1 阿克苏地区农田景观特征
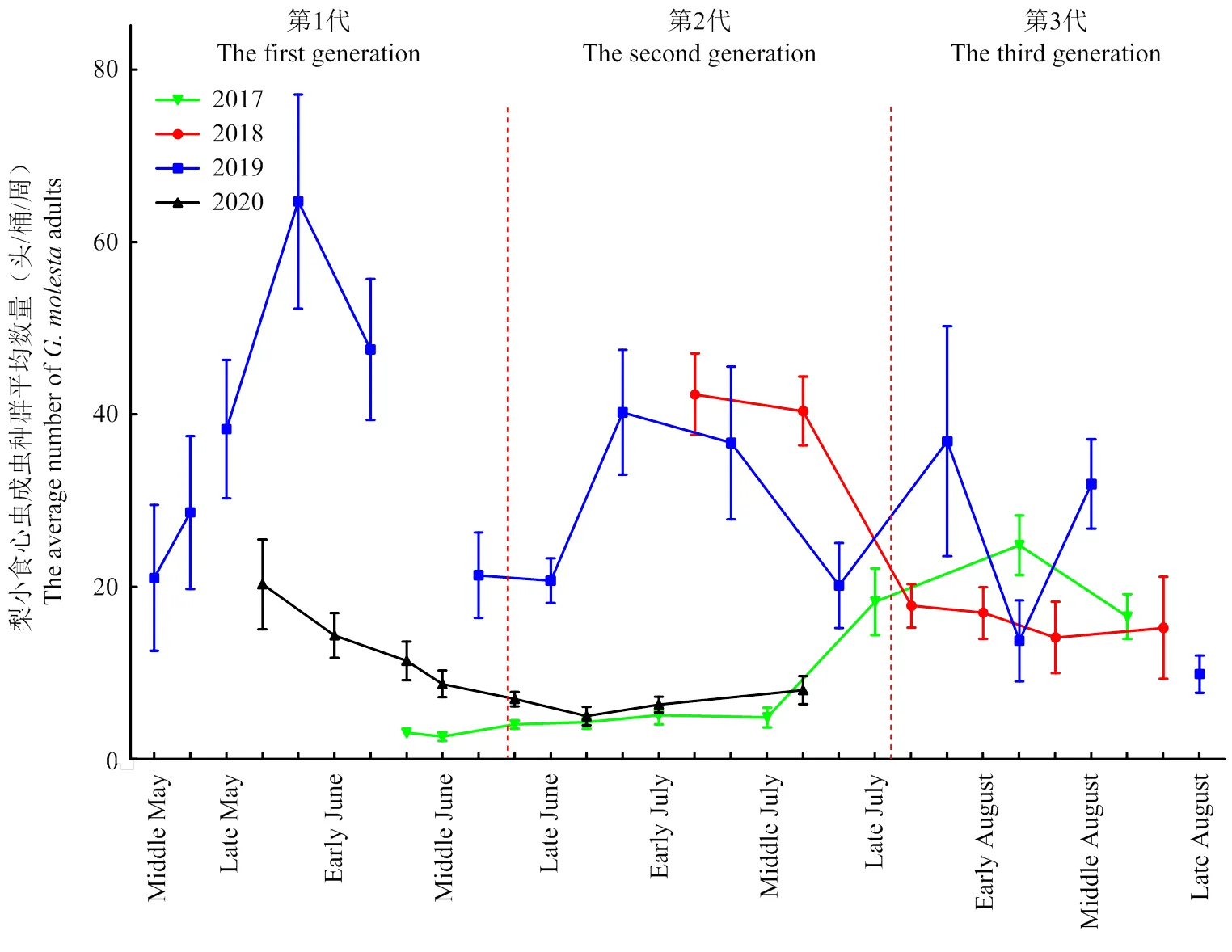
图中标记的误差值为标准误,纵坐标为每次调查5个诱桶所诱集到的梨小食心虫成虫数量平均值
2.3 景观格局对第1代梨小食心虫种群数量的影响
比较4个景观尺度上拟合模型的AICc值,0.5 km尺度上的景观变量拟合第1代梨小食心虫数量模型的AICc值相对较小(AICc=141.98),表明该尺度为最优尺度。其中,该尺度上的最优模型也是所有尺度上模型中的最优模型。模型平均结果显示,第1代梨小食心虫成虫种群丰富度与苹果园外的其他作物所占比例在2.0 km尺度下呈较弱的负相关关系(=0.062)(表2),即在2.0 km尺度下第1代梨小食心虫成虫种群数量随着其他作物比例的增加而减少(图3)。
2.4 景观格局对第2代梨小食心虫种群数量的影响
比较4个景观尺度上拟合模型的AICc值,0.5 km尺度上的景观变量拟合第2代梨小食心虫数量模型的AICc值相对较小(AICc=254.16),表明该尺度为最优尺度。模型平均的结果显示,梨小食心虫第2代成虫种群丰富度与苹果园外其他作物所占比例在4个尺度下均呈显著的负相关关系(0.5 km,<0.001;1.0 km,<0.001;1.5 km,=0.028;2.0 km,=0.043),与0.5、1.0 km尺度非作物生境所占比例负相关(0.5 km,=0.023;1.0 km,=0.019),与香农多样性指数(SHDI)正相关(0.5 km,<0.001;1.0 km,=0.005),与1.0、1.5 km尺度寄主作物面积占比负相关(1.0 km,=0.026;1.5 km,=0.048)(表3),即在各尺度下梨小食心虫第2代成虫种群丰富度均随着其他生境比例的增加而减少(图4-A、5-A、6-A、图7);在0.5、1.0 km尺度下梨小食心虫第2代成虫种群丰富度随着非作物生境比例的增加而减少(图4-B、5-B),随着香农多样性指数的增大而增加(图4-C、5-C);在1.0、1.5 km尺度随着寄主作物比例的增加而减少(图5-D、6-B)。

图中梨小食心虫数量经过bestNormalize ()函数归一化转换。图4—图8同

表2 第1代竞争模型(ΔAICc<2)的模型平均结果及其变量的相对重要性
模型平均结果基于所有竞争模型(AICc<2)计算而来;相对重要性是竞争模型中与每个变量相关的Akaike权重之和Model average results are based on all competing models (AICc<2) calculated; Relative importance is the sum of Akaike weights associated with each variable in the competition model;显著性差异表示为Significant differences were expressed as ***<0.001,**<0.01,*<0.05
表3、表4同The same as Table 3, Table 4

图4 0.5 km景观尺度下景观变量对第2代梨小食心虫的影响

表3 第2世代竞争模型(ΔAICc<2)的模型平均结果及其变量的相对重要性
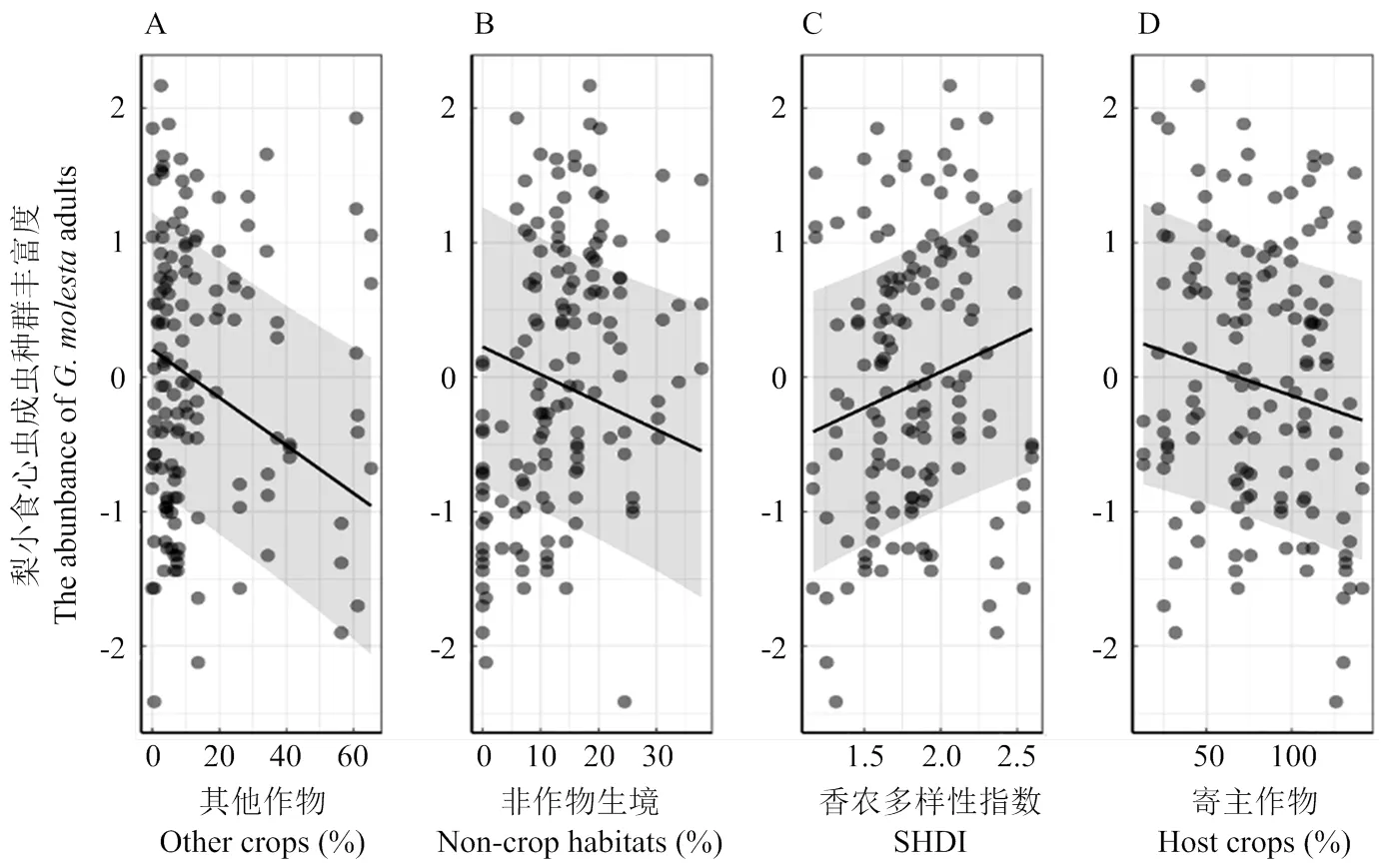
图5 1.0 km景观尺度下景观变量对第2代梨小食心虫的影响

图6 1.5 km景观尺度下景观变量对第2代梨小食心虫的影响
2.5 景观格局对第3代梨小食心虫种群数量的影响
比较4个景观尺度上拟合模型的AICc值,0.5 km尺度上的景观变量拟合第3代梨小食心虫数量模型的AICc值相对较小(AICc=284.94),表明该尺度为最优尺度。模型平均的结果显示,梨小食心虫第3代成虫种群丰富度与苹果园外0.5 km尺度下非作物生境所占比例呈显著负相关(0.5 km,<0.001)(表4),即在0.5 km尺度下梨小食心虫成虫种群丰富度随着非作物生境比例的增加而减少(图8)。
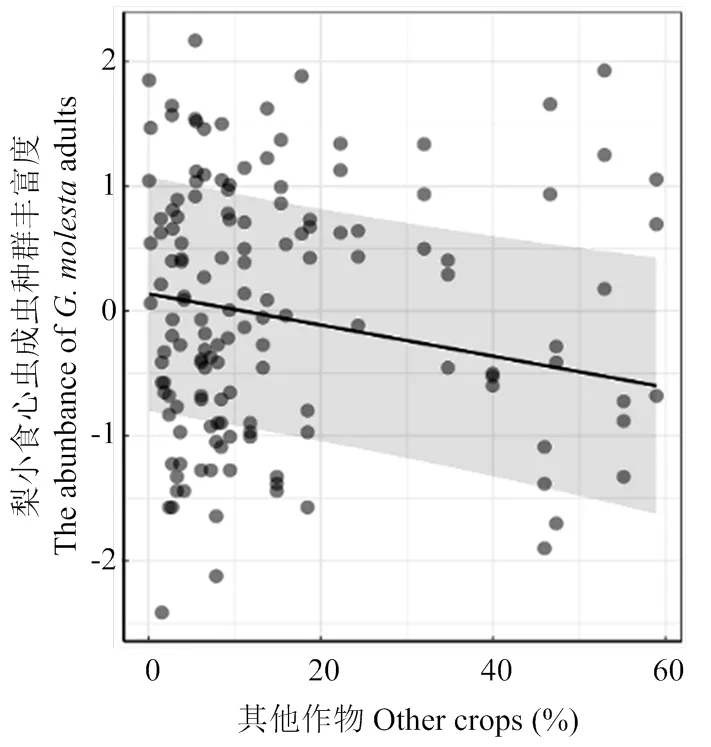
图7 2.0 km景观尺度下景观变量对第2代梨小食心虫的影响
3 讨论
3.1 景观格局是影响苹果园梨小食心虫发生的重要因素
梨小食心虫的越冬场所主要为桃园[16],农田生态系统中越冬代成虫的虫源关系比较明确。然而,梨小食心虫幼虫的寄主植物种类众多,后代虫源关系相对复杂,因此解析农田景观格局对其后代种群发生的影响,明确不同代次梨小食心虫在农田景观系统中的虫源关系,对洞悉梨小食心虫灾变机制,进行虫害预测以及综合防控至关重要。本研究通过分析新疆阿克苏地区农田景观变量与梨小食心虫第1、2、3代成虫种群数量的相关关系发现,农田景观格局对第1、2和3代成虫种群数量均有较为明显的影响。虽然农田景观格局影响苹果园梨小食心虫成虫种群丰富度的主要因素随着世代的不同而发生改变,但整体随着苹果园外寄主作物、其他作物以及非作物生境比例增加而减少,随着景观多样性的增大而增加。
3.2 寄主植物多样性高可加重苹果园梨小食心虫发生
景观配置方面,多样性的景观格局促进了第2代梨小食心虫成虫在苹果园的发生。本试验研究区域内苹果种植面积相对其他寄主作物较大,在寄主作物中占比约为57.8%—61.7%,多样性的农田景观系统中,桃、李、梨等多种梨小食心虫的寄主作物大面积均匀种植,有利于农田系统中梨小食心虫获取连续性的食物资源;第2代幼虫主要在桃、梨等寄主上危害[17-18],因此多样性的农田景观可以为迁入苹果园的梨小食心虫提供更多虫源。此外,多样性的景观系统中,大量的开花植物能为梨小食心虫成虫提供花粉、花蜜,促进雌虫产卵[19],进一步增加了梨小食心虫种群数量。

表4 第3世代竞争模型(ΔAICc<2)的模型平均结果及其变量的相对重要性
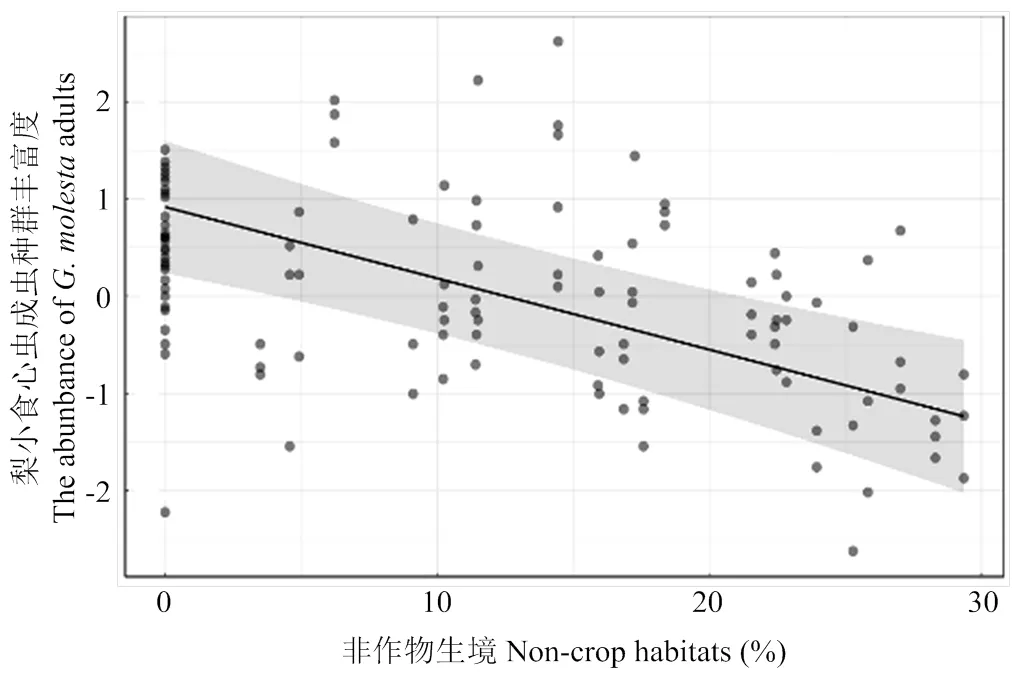
图8 0.5 km景观尺度下景观变量对第3代梨小食心虫的影响
3.3 苹果单一化种植利于稀释苹果园梨小食心虫发生
景观组成方面,寄主作物面积的增大对第2代梨小食心虫有稀释作用,随着寄主作物面积占比的增加梨小食心虫成虫种群数量减少。本研究与Grez等[20]研究蚜虫种群密度与寄主作物面积关系时得到的结果一致,研究发现寄主植物的单一化种植对一些植食性害虫的迁入有稀释作用。苹果是阿克苏地区梨小食心虫寄主作物中面积比例最大的寄主,本试验中寄主作物比例的增加,主要是苹果面积的增加。6月中旬,南疆地区苹果进入果实膨大期,充足的食物资源吸引第2代梨小食心虫迁入苹果园[21-24],此时,大面积的苹果意味着梨小食心虫迁入单位面积苹果园的数量降低,表现出稀释效应。大量的研究表明小麦、棉花等其他作物生境以及草地、林地等非作物生境的存在常有利于天敌群落的发展而提高生物防治的效果[25-27],这可能是梨小食心虫成虫种群丰富度随着其他作物和非作物生境比例的增大而减少的主要原因。
4 结论
农田景观格局对苹果园梨小食心虫成虫种群发生存在明显的调控作用,寄主作物苹果和其他作物占比增大稀释了苹果园梨小食心虫数量,非作物生境占比增大有利于减少梨小食心虫数量。梨、桃、李等其他寄主植物的多样性种植会增大苹果园成虫发生数量。景观格局对梨小食心虫的影响是一个动态的过程,梨小食心虫在寄主间的迁入、迁出使得不同代次的梨小食心虫成虫种群对于农田景观的响应发生了很大改变。在未来的研究中,探索不同寄主作物果园中天敌和害虫群落动态,可以更好地解释景观格局对于虫害发生的调控过程以及建立梨小食心虫虫害发生的预测模型。在未来生产实践中合理设计农田景观格局,适当增加非作物生境和其他作物面积占比,减少桃、梨、苹果的混栽可长期有效地控制梨小食心虫的发生。
[1] Kheirodin A, Cárcamo H A, Costamagna A C. Contrasting effects of host crops and crop diversity on the abundance and parasitism of a specialist herbivore in agricultural landscapes. Landscape Ecology, 2020, 35(5): 1073-1087.
[2] Tscharntke T, Tylianakis J M, Rand T A, Didham R K, Fahrig L, Batáry P, Bengtsson J, Clough Y, Crist T O, Dormann C F,. Landscape moderation of biodiversity patterns and processes—eight hypotheses. Biological Reviews, 2012, 87(3): 661-685.
[3] 李叶晨, 郭雅洁, 翁小倩, 林先云, 池金良, 陈红英, 吴松青, 张飞萍. 景观格局对松褐天牛种群密度的影响. 林业科学, 2020, 56(8): 80-88.
Li Y C, Guo Y J, Weng X Q, Lin X Y, Chi J L, Chen H Y, Wu S Q, Zhang F P. Influence of landscape patterns on population density of(Coleoptera: Cerambycidae). Scientia Silvae Sinicae, 2020, 56(8): 80-88. (in Chinese)
[4] Yang L, Liu B, Zhang Q, Zeng Y, Pan Y, Li M, Lu Y. Landscape structure alters the abundance and species composition of early-season aphid populations in wheat fields. Agriculture, Ecosystems and Environment, 2019, 269: 167-173.
[5] Chaplin-Kramer R, O’Rourke M E, Blitzer E J, Kremen C. A meta-analysis of crop pest and natural enemy response to landscape complexity. Ecology Letters, 2011, 14(9): 922-932.
[6] Yang L, Zeng Y, Xu L, Liu B, Zhang Q, Lu Y. Change in ladybeetle abundance and biological control of wheat aphids over time in agricultural landscape. Agriculture, Ecosystems and Environment, 2018, 255: 102-110.
[7] Yang L, Xu L, Liu B, Zhang Q, Pan Y, Li Q, Li H, Lu Y. Non-crop habitats promote the abundance of predatory ladybeetles in maize fields in the agricultural landscape of northern China. Agriculture, Ecosystems and Environment, 2019, 277: 44-52.
[8] Ardanuy A, Lee M S, Albajes R. Landscape context influences leafhopper and predatoryspp. abundances in maize fields. Agricultural and Forest Entomology, 2018, 20(1): 81-92.
[9] Santoiemma G, Mori N, Tonina L, Marini L. Semi-natural habitats boostpopulations and crop damage in sweet cherry. Agriculture, Ecosystems and Environment, 2018, 257: 152-158.
[10] 阿布力孜·布力布力, 叶丽珍. 新疆特色林果业转型升级发展研究. 山西农业科学, 2017, 45(2): 297-300.
ABULIZI B, YE L Z. Study on transformation and upgrading development of Xinjiang characteristic forestry and fruit industry. Journal of Shanxi Agricultural Sciences, 2017, 45(2): 297-300. (in Chinese)
[11] 李雪冬, 李玉春, 姜中武. 新疆阿克苏地区苹果生产现状及发展前景. 烟台果树, 2020(3): 10-12.
Li X D, Li Y C, Jiang Z W. Present situation and development prospect of apple production in Aksu area of Xinjiang, 2020(3): 10-12. (in Chinese)
[12] 顾沛雯, 辛明. 葡萄病理与昆虫学. 银川: 宁夏阳光出版社, 2018: 408.
Gu P W, Xin M. Grape pathology and entomology. Yinchuan: Ningxia Sunshine Publishing House, 2018: 408. (in Chinese)
[13] 林伟丽, 于江南, 薛光华, 王永平. 新疆阿克苏地区苹果蠹蛾和梨小食心虫消长规律的研究. 新疆农业科学, 2006, 43(2): 100-102.
LIN W L, YU J N, XUE G H, WANG Y P. Study on population fluctuations of(L.) andBusck in Aksy, Xinjiang. Xinjiang Agricultural Sciences, 2006, 43(2): 100-102. (in Chinese)
[14] 江世宏. 园林植物病虫害防治. 重庆: 重庆大学出版社, 2015: 326.
JIANG S H. Plant pest control in gardens. Chongqing: Chongqing University Press, 2015: 326. (in Chinese)
[15] 帕热提·艾山. 阿克苏地区梨小食心虫发生规律及综合防治技术. 农村科技, 2020(4): 38-39.
PARETI A. Occurrence regularity and comprehensive control method ofin Aksu area. Rural Science and Technology, 2020(4): 38-39. (in Chinese)
[16] 宋博, 丁圣彦, 赵爽, 李子晗, 侯笑云. 农业景观异质性对生物多样性及其生态系统服务的影响. 中国生态农业学报, 2016, 24(4): 443-450.
SONG B, DING S Y, ZHAO S, LI Z H, HOU X Y. Effects of agricultural landscape heterogeneity on biodiversity and ecosystem services. Chinese Journal of Eco-Agriculture, 2016, 24(4): 443-450. (in Chinese)
[17] 鲍晓文, 郑峰, 蔡明飞, 仵均祥. 补充营养对梨小食心虫成虫生殖及寿命的影响. 西北农林科技大学学报(自然科学版), 2010, 38(8): 119-123.
BAO X W, ZHENG F, CAI M F, WU J X. Effects of complementary nutrients on adult’s reproduction and longevity of oriental fruit moth,Busck. Journal of Northwest A&F University (Natural Science Edition), 2010, 38(8): 119-123. (in Chinese)
[18] Myers C T, Hull L A, Krawczyk G. Seasonal and cultivar- associated variation in oviposition preference of oriental fruit moth (Lepidoptera: Tortricidae) adults and feeding behavior of neonate larvae in apples. Journal of Economic Entomology, 2006, 99(2): 349-358.
[19] 徐瑞斌, 林硕, 田厚军, 陈艺欣, 赵建伟, 陈勇, 杨广, 魏辉. 不同营养源对小菜蛾成虫嗅觉行为、触角电位反应和繁殖力的影响. 福建农业学报, 2016, 31(2): 161-165.
XU R b, LIN S, TIAN H j, CHEN Y x, ZHAO J w, CHEN Y, YANG G, WEI H. Effect of adult feeding on the olfactory behavior, electroantennogram and fecundity of the diamondback moth,. Fujian Journal of Agricultural Sciences, 2016, 31(2): 161-165. (in Chinese)
[20] Grez A A, Zaviezo T, Mancilla A. Effect of prey density on intraguild interactions among foliar- and ground-foraging predators of aphids associated with alfalfa crops in Chile: a laboratory assessment. Entomologia Experimentalis et Applicata, 2011, 139(1): 1-7.
[21] 庾琴. 影响梨小食心虫暴发为害的主要因子及其寄主选择研究[D]. 太谷: 山西农业大学, 2019.
YU Q. Studies on main factors of outbreak damage ofBusck and host selection[D]. Taigu: Shanxi Agricultural University, 2019. (in Chinese)
[22] 潘云飞. 南疆农田景观组成与农药使用对棉田天敌发生及其控蚜功能的影响[D]. 北京: 中国农业科学院, 2020.
Pan Y F. Effects of landscape composition and pesticide application on natural enemies and their aphid biocontrol services in cotton fields in southern Xinjiang[D]. Beijing: Chinese Academy of Agricultural Sciences, 2020. (in Chinese)
[23] 杨龙. 农田景观格局对华北地区小麦蚜虫天敌发生与控害的影响[D]. 北京: 中国农业科学院, 2016.
Yang L. Effects of landscape pattern on enemy abundance and its control of wheat aphids in northern China[D]. Beijing: Chinese Academy of Agricultural Sciences, 2016. (in Chinese)
[24] 何潭. 多样化农业生态系统在害虫治理中的作用调查. 西南农业学报, 1989, 2(1): 54-59.
He T. Effects of the diversified agricultural ecosystems on pest management. Southwest China Journal of Agricultural Sciences, 1989, 2(1): 54-59. (in Chinese)
[25] 俞晓平, 胡萃, Heong K L. 非作物生境对农业害虫及其天敌的影响. 中国生物防治, 1996, 12(3): 130-133.
Yu X P, Hu C, Heong K L. The effects of non-crop habitats on crop pests and their natural enemies. Chinese Journal of Biological Control, 1996, 12(3): 130-133. (in Chinese)
[26] 侯茂林, 盛承发. 农田生态系统植物多样性对害虫种群数量的影响. 应用生态学报, 1999, 10(2): 245-250.
Hou M L, Sheng C F. Effect of plant diversity in agroecosystems on insect pest populations. Chinese Journal of Applied Ecology, 1999, 10(2): 245-250. (in Chinese)
[27] 武晶, 刘志民. 生境破碎化对生物多样性的影响研究综述. 生态学杂志, 2014, 33(7): 1946-1952.
Wu J, Liu Z M.Effect of habitat fragmentation on biodiversity: A review. Chinese Journal of Ecology, 2014, 33(7): 1946-1952. (in Chinese)
Effects of agricultural landscape on the population dynamic ofadults in apple orchards in southern Xinjiang
Song BoWen1,2, Yang Long2, Pan YunFei2, Li HaiQiang2, Li Hao1,2, Feng HongZu1, Lu YanHui2
1College of Agronomy, Tarim University, Alar 843300, Xinjiang;2State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193
is an important fruit pest in apple orchards in southern Xinjiang, which seriously affects the yield and quality of apple. The influence of agricultural landscape configuration and composition on the population number ofin apple orchards was clarified to provide a theoretical basis for the rational design of agricultural landscape that reduces the harm ofunder the adjustment of cropping structure in southern Xinjiang.A total of 50 apple orchards were selected as experimental sites in Aksu area from 2017 to 2020. The landscape composition within a radius of 2.0 km of each site was investigated. The insect sex pheromone traps were used to investigate the population dynamics ofadult. Regression models of Shannon diversity index (SHDI), perimeter area ratio (PARA), edge density (ED), and the area proportion of non-crop habitats, host crops and other (non-host) crops in landscapes at four scales (0.5, 1.0, 1.5 and 2.0 km) were fitted with the number of adults of the first, second and third generations in apple orchards.In the study area, the proportion of host crops was highest (45.7%-55.0%), followed by other crops (18.2%-21.0%) and non-crop habitats (13.5%-19.7%). There was a negative correlation between the abundance of the first generation adult and the proportion of other crops at 2.0 km scale (=0.062). The abundance of the second generation was negatively correlated with other crops at four scales (0.5 km,<0.001; 1.0 km,<0.001; 1.5 km,=0.028; 2.0 km,=0.043), negatively correlated with the proportion of host crops at 1.0 and 1.5 km scales (1.0 km,=0.026; 1.5 km,=0.048), negatively correlated with the proportion of non-crop habitats at 0.5 and 1.0 km scales (0.5 km,=0.023; 1.0 km,=0.019), but positively correlated with Shannon diversity index (SHDI) (0.5 km,<0.001; 1.0 km,=0.005). The abundance of the third generation was negatively correlated with the proportion of non-crop habitats at 0.5 km scale (<0.001).Increasing the proportion of host crops, other crops, and non-crop habitats within agricultural landscape decreased the occurrence ofin apple orchards. However, landscape diversity (Shannon diversity index) promoted the population number of. Therefore, increasing the area of the other crops and non-crop habitats coupled with no mixed planting of host crops in landscapes could be beneficial to the management of.
agricultural landscape; phytophagous insect; host plant;; apple orchard

10.3864/j.issn.0578-1752.2022.01.008
2021-05-14;
2021-06-16
国家自然科学基金(U2003212)
宋博文,E-mail:499453107@qq.com。杨龙,E-mail:yanglong9005@163.com。宋博文和杨龙为同等贡献作者。通信作者冯宏祖,E-mail:fhzzky@163.com。通信作者陆宴辉,E-mail:luyanhui@caas.cn
(责任编辑 岳梅)
