黄芩素通过诱导活性氧产生抑制MET/Akt信号通路促进人结肠癌SW480细胞凋亡
2022-02-15徐佳丽路上云石东星邱服斌
徐佳丽,路上云,王 佳,石东星,邱服斌
(山西医科大学公共卫生学院营养与食品卫生学教研室,山西 太原 030001)
结肠癌是最常见的恶性肿瘤之一。近年来,结肠癌在我国的发病率和死亡率仍不断上升。尽管在诊断和治疗方面取得了很大进展,然而由于早期症状不明显,被发现时患者已经处于中晚期,治疗效果较差,结肠癌的防治形势仍较为严峻[1-2]。目前,结肠癌的治疗方法主要为手术切除、化学治疗和物理治疗等,易引发不良反应和各种并发症,患者预后较差,死亡率高[3]。
中药及其活性成分有多种生物活性,日益受到人们关注。黄芩素(baicalein)是中药黄芩的主要活性成分,具有抗炎、抗心血管疾病和抗菌等活性[4-6],广泛应用于多种疾病的预防与治疗。此外,越来越多的研究也表明,黄芩素具有很好的抗肿瘤活性,可通过调控磷脂酰肌醇3-激酶/蛋白激酶B(phosphatidylinositol 3-kinase/protein kinase B,PI3K/Akt)、丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)、雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)和NF-κB家族等信号通路,诱导肿瘤细胞凋亡[4,7-9],发挥抗乳腺癌、肝癌、甲状腺癌、肺癌、胃癌和宫颈癌等作用[10-15]。有研究发现,黄芩素能够促进MAPK的激活,调节Akt/mTOR信号通路,引起人结肠癌细胞的凋亡[16],防止大鼠结肠组织病变[17],表明黄芩素很可能在预防和治疗结肠癌的过程中发挥重要作用。
结肠组织中间充质表皮转化因子(mesenchymalepithelial transition factor,MET)和Akt等异常激活与结肠癌的发生、发展、转移和预后密切相关[18]。MET是一种关键的酪氨酸激酶[19],可以调控PI3K/Akt信号通路,抑制甲状腺癌细胞增殖[20],增加骨肉瘤细胞凋亡率[21]等,为此推测黄芩素可能通过调控MET/Akt信号通路来影响结肠癌细胞凋亡。已有研究表明,活性氧(reactive oxygen species,ROS)的产生及其对下游信号通路的调控与多种肿瘤细胞的凋亡密切相关[22-24]。但黄芩素是否通过诱导ROS生成抑制MET/Akt途径而促进结肠癌细胞发生凋亡尚不清楚。因此,本研究通过观察黄芩素对SW480细胞中ROS水平及MET和Akt蛋白表达的影响,探究其对结肠癌细胞抑制和凋亡诱导作用,为黄芩素的开发应用提供理论依据。
1 材料与方法
1.1 药品、试剂和主要仪器
黄芩素(纯度>98%),中国云南西力生物公司,溶于二甲亚砜配置成浓度为80 mmol·L-1的母液。DMEM高糖培养基、RIPA裂解缓冲液、BCA蛋白浓度测定试剂盒和0.25%胰酶溶液,武汉博士德公司;胎牛血清,杭州四季青公司;二甲亚砜和磷酸缓冲液(phosphate buffered sopution,PBST),北京索莱宝公司;兔抗人活化胱天蛋白酶3多抗、兔抗人活化聚腺苷二磷酸核糖聚合酶(ADP-ribosepolymerase,PARP)多抗、兔抗人磷酸化MET(phosphorylated-MET,p-MET)单抗、兔抗人磷酸化Akt(p-Akt)单抗和兔抗人磷酸化重组人源组蛋白(p-histone H3,p-H3)多抗,美国 CST公司;兔抗人β肌动蛋白多抗,南京巴傲得生物科技有限公司;小鼠抗人MET多抗和小鼠抗人H3多抗,沈阳万类生物科技有限公司;小鼠抗人Akt多抗,美国Santa公司;辣根过氧化物酶标记的山羊抗兔和山羊抗小鼠IgG抗体(二抗),武汉三鹰生物技术有限公司;超氧化物歧化酶(superoxide dismutase,SOD)和过氧化氢酶(catalase,CAT)生化试剂盒,南京建成生物工程研究所;Annexin V-FITC/PI细胞凋亡检测试剂盒,北京博尔迈生物技术有限公司;ROS免疫荧光测试剂盒,上海碧云天公司。生物安全柜(ZHJH-C1112B),上海智诚分析仪器制造有限公司;细胞恒温培养箱(CCL-170B-8),新加坡ESCO公司;倒置显微镜(T5),日本Olympus公司;离心机,美国Thermo Forma公司;电泳仪及转膜系统,美国Bio-Rad公司;化学发光成像仪(BG-gdsAUTO 710),北京百晶生物技术有限公司。
1.2 细胞、细胞培养、分组和处理
人结肠癌细胞SW480由山西省肿瘤医院动物实验室馈赠。将SW480细胞在含10%胎牛血清的DMEM高糖培养基中,37℃,5%CO2及饱和湿度条件下进行常规培养。细胞以每孔1×106的密度接种于6孔板,使其24 h内生长密度达到60%~80%。将黄芩素(80 mmol·L-1)储备液用DMSO稀释成40 mmol·L-1工作液备用。在培养基中分别加入工作液使黄芩素终浓度分别为0(细胞对照组),20,40和80 μmol·L-1,处理细胞48 h。
1.3 光镜和结晶紫染色法检测细胞形态和存活率
取1.2分组处理的细胞,在光镜下观察细胞状态并拍照,然后弃培养液,每孔加2 mL 1%戊二醛固定15 min,弃戊二醛,然后每孔加2 mL 0.02%结晶紫溶液染色30 min,洗净晾干,加入8 mL 75%乙醇溶解4~6 h,酶标仪595 nm 下检测吸光度(A595nm)值,75%乙醇溶液为空白,计算细胞存活率。细胞存活率(%)=(给药组A595nm-空白组A595nm)/(细胞对照组A595nm-空白组A595nm)×100%。
1.4 免疫荧光检测ROS水平
细胞以每孔1×106的密度接种于6孔板,分为细胞对照组,黄芩素20,40和80 μmol·L-1组,干预20 h。弃细胞上清液,每孔加入 200 μL(稀释比 1∶1000)的2',7'-二氯二氢荧光素二乙酸酯(2,7-dichlorodihydrofluorescein diacetate,DCFH-DA),37℃培养箱孵育20 min。用无血清细胞培养液洗涤细胞3次,充分去除未进入细胞的DCFH-DA,用荧光显微镜观察,以荧光强弱反映ROS水平。
1.5 生化试剂盒检测SOD和CAT活性
收集1.2分组处理细胞,提取各组细胞总蛋白,并用BCA蛋白浓度测定试剂盒检测各组蛋白浓度。根据试剂盒说明书操作要求检测细胞内SOD和CAT的活性。
1.6 流式细胞仪检测细胞凋亡率
取1.2分组处理的细胞,胰酶消化,收集细胞,反复洗涤后加入85 μL结合缓冲液重悬细胞,转入流式细胞管内,避光加入5 μL碘化丙啶(propidium iodide,PI)染液和 10 μL Annexin Ⅴ,避光孵育30 min,最后加入400 μL结合缓冲溶液,混匀后使用流式细胞仪检测细胞凋亡率。
1.7 Western印迹法检测凋亡相关蛋白水平和磷酸化水平
取1.2分组处理的细胞,提取各组细胞总蛋白。用BCA蛋白浓度测定试剂盒检测各组蛋白浓度。变性后进行聚丙烯酰胺凝胶电泳,湿法转膜,5%脱脂牛奶室温封闭2 h,分别加入相应一抗(抗活化胱天蛋白酶3、活化PARP、Akt、p-Akt、MET、p-MET和p-H3抗体,均1∶1000;抗H3抗体,1∶400;抗β肌动蛋白抗体,1∶8000),4℃孵育过夜;孵育结束后PBST洗涤3次,每次10 min,加入相应二抗(1∶10 000),室温孵育2 h,PBST漂洗3次后ECL发光液显色,于凝胶成像系统下成像,用ImageJ软件检测蛋白条带积分吸光度值,以目标蛋白与内参蛋白条带积分吸光度比值表示目标蛋白相对表达水平,以磷酸化蛋白与总蛋白积分吸光度比值表示蛋白磷酸化水平。
1.8 统计学分析
实验结果数据以±s表示。采用SPSS 25.0软件进行单因素方差分析(One-way ANOVA),组间两两比较采用Tukey检验,量效关系采用pearson相关分析。P<0.05为差异具有统计学意义。
2 结果
2.1 黄芩素对SW480细胞形态和存活率的影响
光镜下观察各组细胞形态(图1A)显示,细胞对照组细胞形态正常,细胞轮廓清晰可见,均处于贴壁状态;黄芩素20,40和80 μmol·L-1组随浓度增加贴壁细胞数量逐渐减少,部分细胞轮廓模糊。结晶紫染色结果显示,与细胞对照组相比,黄芩素20,40和80 μmol·L-1组贴壁细胞数量明显减少(图1B1)。图1B2结果显示,黄芩素可浓度依懒性降低细胞存活率(r=-0.928,P<0.01)。
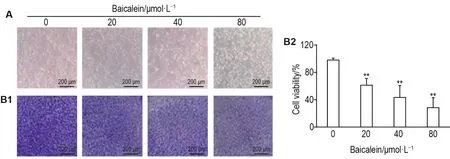
Fig.1 Effect of baicalein on cell viability of SW480.SW480 cells were treated with baicalein for 48 h.A was the images observed under a light microscope.B1 was the images of SW480 cells detected by crystal violet staining,B2 was the the semi-quantitative result of B1.Cell viability(%)=(A595 nmof baicalein group-A595 nmof blank group)/(A595 nmof cell control group-A595 nmof blank group)×100%.±s,n=3.**P<0.01,compared with cell control(0)group.
2.2 黄芩素对SW480细胞ROS生成的影响
与细胞对照组相比,黄芩素20,40和80 μmol·L-1处理20 h后,细胞内荧光强度显著增强(P<0.01),表明黄芩素可诱导ROS生成(图2)。

Fig.2 Effect of baicalein on reactive oxygen species(ROS) generation in SW480 cells by fluorescence probe.The cells were treated with baicalein for 20 h.B was the semi-quantitative result of A.FI:fluorense intensity.±s,n=3.**P<0.01,compared with cell control group.
2.3 黄芩素对SW480细胞SOD和CAT活性的影响
与细胞对照组相比,黄芩素处理48 h后,SW480细胞SOD和CAT活性明显降低(P<0.05,P<0.01),表明黄芩素诱导ROS的产生可能与抑制SOD和CAT的活性有关(表1)。
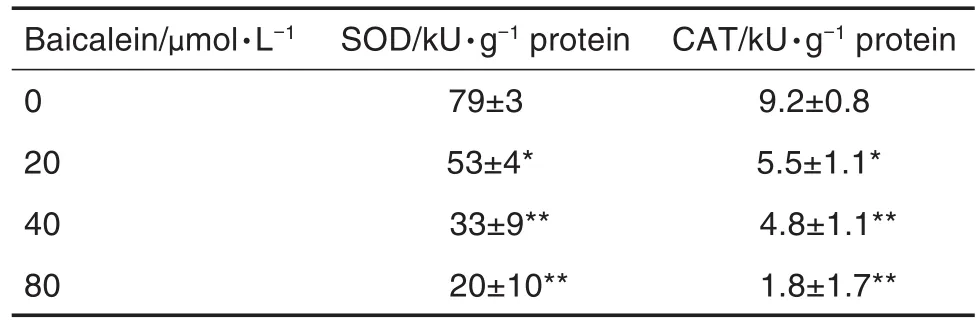
Tab.1 Effect of baicalein on activities of superoxide dismutase(SOD)and catalase(CAT)in SW480 cells
2.4 黄芩素对SW480细胞凋亡的影响
流式细胞仪检测结果显示(图3),与细胞对照组相比,黄芩素 40 和 80 μmol·L-1处理 48 h 后,SW480细胞凋亡率明显升高(P<0.01)。黄芩素可浓度依赖性诱导SW480细胞凋亡(r=0.988,P<0.01)。

Fig.3 Effect of baicalein on apoptosis of SW480 cells detected by flow cytermetry.See Fig.1 for the cell treatment.±s,n=3.**P<0.01,compared with cell control group.
2.5 黄芩素对SW480细胞活化胱天蛋白酶3和活化PARP蛋白表达的影响
Western印迹结果显示(图4),与细胞对照组相比,黄芩素40和80 μmol·L-1组SW480细胞中活化胱天蛋白酶3和活化PARP蛋白水平明显升高(P<0.05,P<0.01),表明黄芩素可通过活化胱天蛋白酶依赖性凋亡途径诱导SW480细胞死亡。

Fig.4 Effect of baicalein on protein expression levels of cleaved-polymerase(PARP)and cleaved-caspase 3 in SW480 cells by Western blotting.See Fig.1 for the cell treatment.B was the semi-quantitative result of A.±s,n=3.*P<0.05,**P<0.01,compared with cell control group.
2.6 黄芩素对SW480细胞MET/Akt信号通路相关蛋白磷酸化水平的影响
与细胞对照组相比,黄芩素20,40和80 μmol·L-1组 SW480细胞中 p-MET/MET,p-Akt/Akt和p-H3/H3比值明显降低(P<0.05,P<0.01)(图5)。

Fig.5 Effect of baicalein on mesenchymal-epithelial transition factor(MET),protein kinase B(MET/Akt)pathway related protein phosplorylation levels in SW490 cells by Western blotting.See Fig.1 for the cell treatment.B was the semi-quantitative result of A.±s,n=3.*P<0.05,**P<0.01,compared with cell control group.
3 讨论
本研究结果显示,黄芩素可诱导ROS产生,抑制MET/Akt信号通路,从而抑制SW480细胞增殖,促进结肠癌细胞凋亡。
ROS是机体氧化应激时产生的主要分子,一直以来被认为是肿瘤发生、发展和复发的重要因素[25-27]。在本研究中,黄芩素干预会明显抑制SW480细胞增殖,诱导细胞皱缩并发生凋亡;SW480细胞内ROS荧光强度明显增加,而抗氧化酶SOD和CAT活力均明显下调,表明黄芩素可诱导结肠癌SW480细胞产生ROS,抑制抗氧化酶活力。提示ROS可能在调节SW480细胞凋亡中发挥重要作用。
胱天蛋白酶3是胱天蛋白酶家族的重要成员,是发生凋亡的标志性蛋白[28]。胱天蛋白酶3在内源性通路中的激活可以调控细胞凋亡的启动与执行[29]。本研究结果表明,黄芩素干预后,SW480细胞活化胱天蛋白酶3和活化PARP蛋白表达水平明显增加,说明黄芩素可诱导SW480细胞凋亡。
研究报道,调控细胞凋亡可能与一系列蛋白表达有关,例如MAPK,NF-κB和MET等。MET在细胞增殖、代谢及肿瘤产生、转移中扮演着重要角色,是近年来抗肿瘤研究的热门靶点。T-淋巴因子激活的杀伤细胞来源的蛋白激酶(T-lymphokine-activated killer cell-originated protein kinase,TOPK)在结肠癌、肺癌和乳腺癌等癌症中高度表达[30-32]。p-H3作为TOPK的底物,是最广泛使用的有丝分裂标记物[33]。研究表明,MET可以激活TOPK进而激活Akt促进结肠癌发生[25]。本研究结果显示,经黄芩素处理后,SW480细胞内p-MET,p-H3和p-Akt蛋白表达水平明显下调,表明黄芩素可能影响肿瘤细胞MET/Akt信号通路,进而诱导SW480细胞凋亡。
综上,本研究表明,诱导人结肠癌SW480细胞产生ROS,抑制MET/Akt信号途径,激活胱天蛋白酶3,可能是黄芩素诱导SW480细胞凋亡的机制之一。本研究为结肠癌防治及黄芩素的开发提供理论依据。
