过表达IDO骨髓间充质干细胞分泌外泌体通过不同分子轴调节DC及T细胞
2022-02-13刘星佑陈鑫昊黄尹霞李敏撒亚莲贺继刚
刘星佑 陈鑫昊 黄尹霞 李敏 撒亚莲 贺继刚
1云南中医药大学(昆明650000);2云南省第一人民医院心脏大血管外科(昆明650000)
20 世纪以来,世界范围内随着人口老龄化、缺血性心脏病、糖尿病和肥胖等危险因素的增加,心力衰竭(heart failure,HF)的发病率不断上升[1]。据统计,全球成年人中约有2 600 万心力衰竭患者,尽管已有治疗心力衰竭的一些方法出现,如药物治疗、双心室起搏、左心室辅助装置和全人工心脏,但心脏移植仍是治疗终末期心力衰竭的“黄金标准”,也是提高终末期心力衰竭患者生存率和生活质量的有效途径[2]。而器官排异反应是影响移植器官存活的主要问题。目前研究认为间充质干细胞的旁分泌是间充质干细胞完成其生物功能的重要机制[3-4]。课题组前期已经证明过表达吲哚胺2,3 双加氧酶(indoleaminae 2,3-dioxygenase,IDO)的骨髓间充质细胞(bone marrow mesenchymal stem cells,BMSC)分泌的外泌体(exosome)即BMSCIDO-exosome 可以有效抑制免疫排斥反应[5-6]并发现BMSCIDO-exosome 中miRNA-540-3p 表达明显增高且差异具有统计学意义,其功能涉及抗免疫排斥反应,可能是抑制免疫排斥反应的关键分子。笔者采用靶基因预测数据库、PCR、Western blot(WB)及双荧光素酶的方法证实miRNA-540-3p对应的靶基因为CD74,进而采用Western blot 的方法对CD74 下游的p-ERK1/2、p-p65、p-AKT 的表达进行检测。现报道如下。
1 材料与方法
1.1 实验材料、试剂定量PCR 仪购自美国Light-Cycle 96;紫外分光光度计购自美国NanoDropND-1000;凝胶成像仪、垂直电泳槽、湿式转膜槽均购自BIO-RAD;CD74 抗体购自abcam ab202844(批号:GR3206250-17 有效期至2022年9月);CD28 抗体购自abcam ab243228(批号:GR3277276-3 有效期至2022年9月);CCL4 抗体购自abcam ab25129(批号:GR189200-6 有效期至2021年9月);p-p65抗体购自abcam ab86299(批号:GR3204852-21 有效期至2022年9月);p-ERK1/2 赛默飞MA5-15174(批号:BA 06044196 有效期至2022年9月);引物合成来自广州invitrogen;exosome 提取试剂盒购自锐博(批号U0122 有限期至2022年3月)。
1.2 实验方法
1.2.1 模型建立按照文献[3]建立过表达IDO 大鼠骨髓间充质干细胞并提取外泌体、对照组分泌的外泌体,以及大鼠DC 细胞、T 细胞。
1.2.2 共培养及分组与DC 细胞共培养实验分为5 组(每组6 例):A,DC 组;B,DC+脂多糖(LPS)组;C,BMSC-exosome+DC+LPS 组;D,BMSC空载体-exosome+DC+LPS 组;E,BMSCIDO-exosome+DC+LPS 组。与T 细胞共培养实验分为5 组(每组6 例):F,T 组;G,T+脂多糖(LPS)组;H,BMSC-exosome+T+LPS组;I,BMSC空载体-exosome+T+LPS 组;J,BMSCIDO-exosome+T+LPS 组。48 h 后采用流式细胞仪检测共培养组DC 细胞及T 细胞内的miR-540-3p 的表达情况。采用WB 检测共培养DC 细胞及T 细胞内CD74、CD28、CCL4、p-ERK1/2、p-p65 及p-AKT 表达情况。
1.2.3 采用RT-PCR 检测共培养48 h A-J 组中miR-540-3p 表达量应用beacon designer 7.90 设计Q-PCR 引物,引物序列如下:U6:5'GCTTCGGCAGCACATATACTAAAAT3'(forward)5'CGCTTCACGAATTTGCGTGTCAT3'(reverse);miR-540-3p:5'GTCGTATCCAGTGCGTGTCGTGGAGTCGGCAATTGCACTGGATACGACGCCCAGG3'(RTprimer),5'GGAGGTCAGAGGTCGATC3'(gene-specific stem-loop primer)。完成总RNA提取,反转录为cDNA,完成目的基因扩增(95 ℃,10 min;95 ℃,2 s;60 ℃,20 s;70 ℃,10 s 共完成45 ~50 个循环)。收集信息,做Ct 值分析。
1.2.4 Western blot采用Western blot 实验检测共培养48 h DC 细胞及T 细胞内CD74、CD28、CCL4、p-ERK1/2、p-p65 及p-AKT 表达情况。
1.3 统计学方法使用SPSS 15.0 软件进行统计分析。数据以()表示。各组(每组6 例样本)间比较采用单因素方差分析,所有分组经检测均为正态分布。所有统计假设检验均为双侧的。以P<0.05 为差异有统计学意义。
2 结果
2.1 与DC细胞共培养48 h后DC细胞中miR-540-3p 的表达量各exosome 与DC 细胞共培养48 h 后采用RT-PCR 方法检测DC 细胞中miR-540-3p 的表达量,可见,BMSCIDO-exosome+DC+LPS 组DC 细胞内miR-540-3p 表达量最高。见图1。

图1 不同BMSCs 细胞分泌外泌体与DC 细胞共培养48 hFig.1 Different BMSCs secreted exosomes and co cultured with DC cells for 48 hours
2.2 与T 细胞共培养48 h 后T 细胞中miR-540-3p的表达量各exosome 与T 细胞共培养48 h 后采用RT-PCR 方法检测T 细胞中miR-540-3p 的表达量,可见BMSCIDO-exosome+T+LPS 组T 细胞内miR-540-3p 表达量最高。见图2。
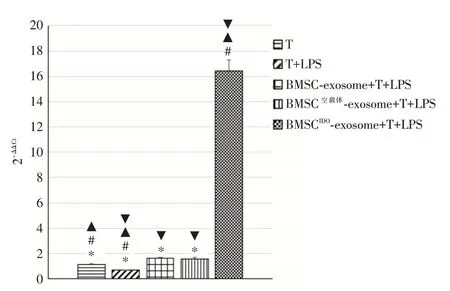
图2 不同BMSCs 细胞分泌外泌体与T 细胞共培养48 hFig.2 Different BMSCs secreted exosomes and co cultured with T cells for 48 hours
2.3 与DC 细胞共培养48 h 后,DC 细胞内CD74、CD28、CCL4 的表达量采用Western blot 检测BMSCIDO-exosome与DC细胞共培养48 h后,DC细胞内CD74、CD28、CCL4的表达量,可见BMSCIDO-exosome+DC 细胞培养组中CD74、表达较其它对照组明显降低,CD28 及CCL4 表达未见规律性变化(图3)。
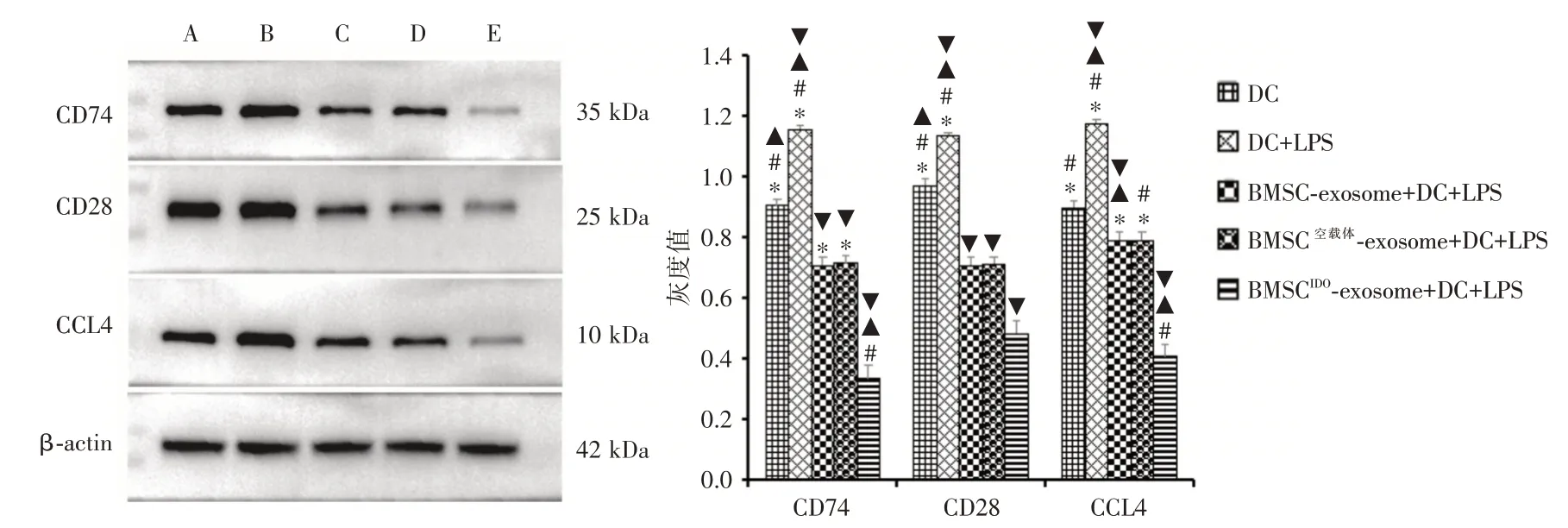
图3 Western blot 检测DC 细胞中miRNA-540-3p 靶基因蛋白CD74、CD28、CCL4 的表达Fig.3 The expression of miRNA-540-3P target gene proteins CD74,CD28 and CCL4 in DC cells was detected by Western blot
2.4 与T细胞共培养48 h后,T细胞内CD74、CD28、CCL4 的表达量采用Western blot 检测BMSCIDOexosome 与T 细胞共培养48 h 后,T 细胞内CD74、CD28、CCL4 的表达量。可见BMSCIDO-exosome+T 细胞共培养组中T 细胞内的CD74 表达明显降低,CD28 及CCL4 无规律性表达(图4)。

图4 Western blot 检测T 细胞中miRNA-540-3p 靶基因蛋白CD74、CD28、CCL4 的表达Fig.4 The expression of miRNA-540-3P target gene proteins CD74,CD28 and CCL4 in T cells was detected by Western blot
2.5 双荧光素酶报告CD74 基因与miRNA-540-3p 的靶向结合力使用psi-check 双荧光素酶报告基因质粒系统,检测CD74 基因与miRNA-540-3p的靶向结合力,可见miRNA-540-3p 与CD74 基因具有靶向结合能力(图5)。

图5 双荧光素酶报告基因验证miRNA-540-3p 与CD74 基因具有靶向结合能力Fig.5 Double luciferase reporter gene verifies that miRNA-540-3P has targeted binding ability with CD74 gene
2.6 与DC细胞共培养48 h后DC细胞内p-NF-κB(p-p65)、p-ERK1/2、p-AKT 蛋白的表达采用Western blot 检测BMSCIDO-exosome 与DC 细胞共培养48 h 后DC 细胞内p-NF-κB(p-p65)、p-ERK1/2、p-AKT 蛋白的表达,可见BMSCIDO-exosome+DC 细胞培养组中p-p65 表达明显降低,p-ERK1/2、p-AKT表达未见规律性变化(图6)。

图6 Western blot 检测DC 细胞中下游蛋白p-p65、p-ERK1/2、p-AKT 的表达Fig.6 Expression of p-P65,P-ERK1/2 and P-Akt in the middle and lower reaches of DC cells were detected by Western blot
2.7 与T 细胞共培养48 h 后T 细胞内p-NF-κB(p-p65)、p-ERK1/2、p-AKT 蛋白的表达量采用Western blot 检测BMSCIDO-exosome 与T 细胞共培养48 h后T细胞内p-NF-κB(p-p65)、p-ERK1/2、p-AKT蛋白的表达,可见BMSCIDO-exosome+T 细胞共培养组中T 细胞内的p-AKT 表达明显降低,p-NF-κB(p-p65)、p-ERK1/2 无规律性表达(图7)。

图7 Western blot 检测T 细胞中下游蛋白p-p65、p-ERK1/2、p-AKT 的表达Fig.7 Expression of p-P65,P-ERK1/2 and P-Akt in the middle and lower reaches of T cells were detected by Western blot
3 讨论
器官移植是治疗终末衰竭器官的一种有效方法,但由于免疫排异反应及昂贵的免疫抑制剂使得该项技术发展非常缓慢。基于此,实验前期已经证实过表达IDO 基因的SD 大鼠骨髓间充质干细胞分泌的外泌体(BMSCIDO-exosome)可以有效抑制免疫排斥反应[3-4]。在此基础上本研究通过RTPCR 发现与BMSCIDO-exosome 共培养48 h 后SD 大鼠DC 细胞、T 细胞内miRNA-540-3p 表达明显增高且差异具有统计学意义,其功能涉及抗免疫排斥反应,可能是抑制免疫排斥反应的关键分子。
miRNA-540-3p 是一类研究不多的microRNA。人的PPARα,ACOX1,CPT1A,ABCD3 和PPARδ(过氧化物酶体增殖剂激活受体-δ)的3'UTR 靶标相同的种子序列的hsa-miR-6801 被鉴定为miRNA-540-3p 功能等同物[7]。据SUDHIR 等[8]报道:肝再生增强蛋白缺乏症可以通过诱导氧化应激和microRNA-540-3p 表达促进肝脂肪变性。目前国、内外未见miRNA-540-3p 与免疫调节的相关报道。
目前已知miRNA 可以有多个对应的靶基因,其通过下调靶基因蛋白的表达,影响受体细胞的生物学功能。进而课题组采用12 个miRNA 靶基因预测数据库(miRWalk、Microt4、miRanda、mirbridge、miRDB、miRMap、miRNAMap、Pictar2、PITA、RNA22、RNAhybrid、Targetscan)对miRNA-540-3p的靶基因进行预测(筛选条件:同一个靶基因必须被≥10 个数据库检索到,同时靶基因与miRNA 的匹和度≥8),可见miRNA-540-3p 对应的靶基因为CD74、CD28 和CCL4。进一步通过Western blot 检测证实,在体外与BMSCIDO-exosome 共培养的DC 细胞、T 细胞内CD74 蛋白表达明显减少。通过双荧光素酶报告基因实验证明miRNA-540-3p 与CD74基因具有靶向结合能力。
CD74 是一种具有Ⅱ型拓扑结构的跨膜蛋白,其在抗原呈递中的作用已得到广泛的研究[9]。目前研究认为CD74 是MHCⅡ(如DC 细胞,B 细胞和巨噬细胞)抗原呈递细胞中参与抗原呈递最强分子之一[10]。最近发现CD74 不仅对MHCⅡ抗原呈递途径很重要,而且对交叉表达的病毒MHCⅠ类抗原呈递途径也有重要意义。在缺乏CD74 的情况下,DC 细胞的交叉启动能力明显受损,因为MHCⅠ类(MHCⅠ)复合物不能有效地靶向于内源性溶酶体区。CD74 对DC 细胞迁移的影响在体内也得到证实,并且在其它表达CD74 的细胞如B 淋巴细胞中也是有效的。CD74 的胞质N 末端结构域可介导这些效应。CD74 在DC 细胞和B 细胞中与肌球蛋白Ⅱ相互作用,肌球蛋白Ⅱ是一种参与DC 细胞迁移的运动蛋白。在CD74 缺陷的DC 细胞中,内吞囊泡的数量显著增加,但它们的体积明显缩小[11]。另外,据报道CD74 受体可以与MIF2(巨噬细胞迁移抑制因子2)形成复合物,两者形成复合物可启动下游信号通路(ERK1/2)NF-κB、PI3K/AKT[12-14]。
本研究进一步采用Western blot 检测与BMSCIDOexosome 共培养48 h 后DC 细胞、T 细胞内的ERK1/2、NF-κB、AKT 的表达,可见在DC 细胞中p-NF-κB(p-p65)的表达呈规律性下降,具有显著性差异,虽然ERK1/2、AKT 在BMSCIDO-exosome 共培养48 h后DC 细胞中也有下降。但在BMSC-exosome 与BMSC空载体-exosome 比较中,其差异也有统计学意义,所以不能排除是由于载体引起的ERK1/2、AKT的变化。在与BMSCIDO-exosome共培养48 h后T细胞中p-AKT 的表达呈规律性下降,具有显著性差异。虽然ERK1/2、p-NF-κB(p-p65)在BMSCIDO-exosome共培养48 h 后T 细胞中也有下降。但在BMSCexosome 与BMSC空载体-exosome 比较中,其差异也有统计学意义,所以不能排除是由于载体引起的ERK1/2、p-NF-κB(p-p65)的变化。
NF-κB 信号通路:NF-κB/Rel 蛋白包括NF-κB2 p52/p100、NF-κB1 p50/plo5、c-Rel、RelA/p65、RelB。这些蛋白均形成二聚体转录因子,它们控制的基因调控众多的生物学过程如先天性和获得性免疫、炎症、应激反应、B 细胞形成、淋巴器官的生成。在细胞炎症反应、免疫应答等过程中NF-κB起到关键性作用。NF-κB 的错误调节会引发自身免疫病、慢性炎症以及很多癌症[15]。
Akt 网络的关键要素之一是丝氨酸/苏氨酸蛋白激酶,被称为哺乳动物雷帕霉素靶蛋白(mTOR)。当免疫系统错误地将自身组织识别为异物并触发免疫反应时,就会发生自身免疫性疾病。免疫系统的自身反应性是多因素的,可能导致致病性自身免疫和相关的自身免疫性疾病[16]。尽管自身免疫性疾病发病机理的确切机制尚不清楚,但T 细胞、B 细胞和髓系细胞的过度激活和功能异常已被广泛研究[17]。
综上所述:BMSCIDO-exosome 通过miRNA-540-3p(+)/CD74(-)/p-p65(-)分子轴抑制DC 细胞的激活,通过miRNA-540-3p(+)/CD74(-)/p-AKT(-)分子轴抑制T 细胞的激活,从而抑制免疫反应。
