广西红花檵木炭疽病病原鉴定及其生物学特性
2022-02-08叶云峰杜婵娟赵廷昌刘思情
叶云峰,杜婵娟,杨 迪,赵廷昌,刘思情,付 岗
(1.广西农业科学院园艺研究所,南宁 530007;2.广西农业科学院植物保护研究所,南宁 530007;3.中国农业科学院植物保护研究所,北京 100093;4.南阳师范学院生命科学与农业工程学院,河南 南阳 473061)
【研究意义】檵木是金缕梅科檵木属常绿灌木植物,主要分布于我国、印度和日本等亚洲地区。我国的檵木主要有白花檵木(LoropetalumchinensisOliv.)和红花檵木(L.chinensevar.rubrum)两大类。白花檵木常生长于山野灌木丛中,主要作为中药材使用[1-2]。而红花檵木是特产于我国的观赏性植物,叶红花艳、花期长,深受园林绿化市场欢迎。自20世纪80年代开始产业化栽培以来,已在我国华南、西南、华东和华中等地20多个省(区)市大量应用,并出口至美国、日本、韩国和德国等多个国家,成为我国园林花卉产业的特色产品之一[3-4]。在广西地区,红花檵木已在南宁市、桂林市、柳州市、梧州市和北海市5大城市绿化中广泛应用[5],但随着种植面积的不断扩大,其病虫害发生情况也日趋严重。2021年6—8月,广西农业科学院香蕉病害研究团队课题组调查发现广西南宁市城市绿化带及多个苗木生产基地的红花檵木发生一种严重的叶部病害,发病率在30%以上,引起大量叶片萎蔫干枯甚至脱落,影响树势生长,严重影响苗木生产及园林美观。因此,开展该病害的病原菌分离鉴定和生物特性研究,明确病原菌的分类地位和生物学特性,对红花檵木病害的科学防治及其产业的健康发展具有重要意义。【前人研究进展】目前,对红花檵木的研究主要集中在组培快繁、栽培管理、生理特性和滞尘能力等方面[6-9],关于红花檵木病害及病原鉴定的研究报道较少。张国辉等[10]采用核糖体转录间隔区(ITS)保守序列测序和形态学鉴定方法将贵州省凯里市的红花檵木炭疽病病原鉴定为胶胞炭疽菌(Colletotrichumgloeosporioide)。Qiu等[11]结合形态学和多基因[ITS、钙调蛋白(CAL)、几丁质合酶(CHS-1)、甘油醛-3-磷酸脱氢酶(GAPDH)和肌动蛋白(ACT)]系统发育分析法将江西南昌梅岭风景区的红花檵木原种(白花檵木)炭疽病病原鉴定为果生刺盘孢(C.fructicola)。卢东升等[12]采用显微镜观察分生孢子形态的方法将河南省信阳市的红花檵木灰斑病和褐斑病病原分别鉴定为拟盘多毛孢菌属(Pestalotiopsis)和尾孢菌属(Cercospora)真菌,但未在种水平进行鉴定。此外,余国清等[4]介绍红花檵木的主要病害有4种,包括炭疽病、立枯病、花叶病毒病和煤污病等,描述了这些病害的危害症状并提出具有针对性且切实可行的综合防治技术措施。王燕和朱发仁[13]、王燕[14]分别对湖南省长沙市和常德市的红花檵木花叶病毒病危害症状和发生规律进行系统研究,但未对病原进行分类鉴定。许璐等[15]于2021年对红花檵木病害的研究进展进行了综述,概述其病害共8种,包括7种真菌病害和1种病毒病害,其中,立枯病、炭疽病、煤污病、黑斑病、灰斑病和褐斑病等6种真菌和花叶病毒病均分布在我国,另外1种真菌病害枯梢病(Phytophthoraramorum)分布在美国[16]。目前,将红花檵木病害病原菌鉴定到种水平的只有炭疽病和枯梢病,对其余病害的病原均未进行系统鉴定和研究。可见,国内外对红花檵木的研究多停留在表现症状的初步调查鉴定阶段,未进行深入探究病原菌的分类地位和生物学特性。【本研究切入点】红花檵木为广西重要的园林观赏植物,近年来发生一种严重的叶部病害,严重影响苗木生产及园林美观,但目前尚无关于该病害病原鉴定及生物学特性研究的报道。【拟解决的关键问题】分离红花檵木叶部病害的病原菌,测定其致病性,并结合形态学特征和多基因系统发育分析结果对病原菌进行分类鉴定。同时,对病原菌的生物学特性进行测定,为该病害的正确诊断和科学防治提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 红花檵木病叶 供试红花檵木病叶采集于广西南宁市西乡塘区路西村苗木生产基地不同发病时期的红花檵木植株。
1.1.2 培养基 马铃薯葡萄糖培养基(PDA)[17]:马铃薯200.0 g,葡萄糖20.0 g,琼脂18.0 g,H2O 1.0 L,pH 7.0。基础培养基[18]:蔗糖30.0 g,KNO32.0 g,K2HPO41.0 g,KCl 0.5 g,MgSO40.5 g,FeSO40.01 g,琼脂17.0 g,H2O 1.0 L。
1.1.3 杀菌剂 供试杀菌剂共12种,分别为10%苯醚甲环唑水分散粒剂(先正达南通作物保护有限公司)、250 g/L丙环唑乳油(山东潍坊双星农药有限公司)、430 g/L戊唑醇悬浮剂(江苏七洲绿色化工股份有限公司)、30%吡唑醚菌酯悬浮剂(河南勇冠乔迪农业科技有限公司)、22.5%啶氧菌酯悬浮剂(安阳市锐普农化有限公司)、41.7%氟吡菌酰胺悬浮剂 [拜耳(中国)有限公司]、50%啶酰菌胺水分散粒剂(巴斯夫欧洲公司)、400 g/L嘧霉胺悬浮剂[拜耳(中国)有限公司]、80%乙蒜素乳油(河南科邦化工有限公司)、80%代森锰锌可湿性粉剂[陶氏益农农业(中国)科技有限公司]、50%多菌灵可湿性粉剂(上海悦联化工有限公司)和450 g/L咪鲜胺水乳剂(青岛中达农业科技有限公司)。
1.2 试验方法
1.2.1 病样采集及症状观察 2021年8月采集红花檵木病叶带回实验室用于病原菌分离。同时,对病害早期至晚期的症状进行观察、拍照和记录。
1.2.2 病原菌分离纯化 采用组织分离法分离病原菌。在病叶的病健交界处切取大小为3 mm×3 mm的组织块,用75%乙醇消毒10 s,0.1%氯化汞消毒30 s,无菌水冲洗3次,置于PDA培养基上28.0 ℃培养,待长出菌丝后,挑取菌落边缘进行纯化,选取出现率高且形态一致的菌落进行单孢分离培养。
1.2.3 致病性测定 将单孢分离的菌株接种于PDA培养基上,28.0 ℃培养10 d,用灭菌打孔器打取直径为5 mm的菌饼,备用。选择红花檵木盆栽苗上的10张叶片,用灭菌针头轻微刺伤,将制备好的菌饼接种于伤口处,套袋保湿,以接种空白PDA菌饼的叶片为对照,所有接种植株置于气候箱内,30.0 ℃培养。定期观察和记录发病症状,并从发病部位进行病原菌再分离。
1.2.4 病原菌鉴定 形态学特征观察:将病原菌接种至PDA培养基,28.0 ℃恒温培养15 d,观察记录菌落颜色和形态,在光学显微镜下观察菌体形态,并随机测量100个分生孢子的大小。
多基因系统发育分析:采用真菌DNA提取试剂盒(北京索莱宝生物科技有限公司)提取病原菌总DNA。基因选择及PCR扩增参照Weir等[19]、徐丹丹等[20]的方法(略有改动),使用6对引物分别对病原菌代表性菌株H6的ITS、ACT、β-微管蛋白(TUB2)、GAPDH、CHS-1和CAL等基因序列进行PCR扩增,引物信息见表1。PCR反应体系25.0 μL:2×TaqPCR Master Mix 12.5 μL,无菌ddH2O 9.5 μL,上、下游引物各1.0 μL,DNA模板1.0 μL。扩增程序:94.0 ℃预变性5 min;94.0 ℃ 30 s,各引物(ITS:52.0 ℃;ACT和CHS-1:58.0 ℃;TUB2:55.0 ℃;GAPDH和CAL:59.0 ℃)30 s,72.0 ℃ 45 s,进行35个循环;72.0 ℃再延伸10 min。PCR产物经1.5%琼脂糖凝胶电泳检测确认有目标条带后,委托北京擎科生物科技公司进行测序。测序结果在NCBI数据库中进行BLAST同源性比对,并采用MEGA-X的Neighbor Joining法构建基于ITS、ACT、TUB2、GAPDH、CHS-1和CAL的多基因系统发育进化树,确定病原菌的分类地位。
1.2.5 病原菌生物学特性测定 温度对病原菌菌丝生长的影响:将病原菌菌株H6移至PDA培养基中,28.0 ℃培养5 d后,用直径5 mm的打孔器打取菌丝块,接种至PDA培养基上,分别置于4.0、7.0、10.0、13.0、16.0、19.0、22.0、25.0、28.0、31.0、34.0和37.0 ℃培养箱中,培养7 d后用十字交叉法测量各处理的菌落直径,计算菌落扩展直径,筛选最适宜菌株H6菌丝生长的温度。菌落扩展直径=菌落直径-接种的菌丝块直径(5 mm)。每个处理重复3次。
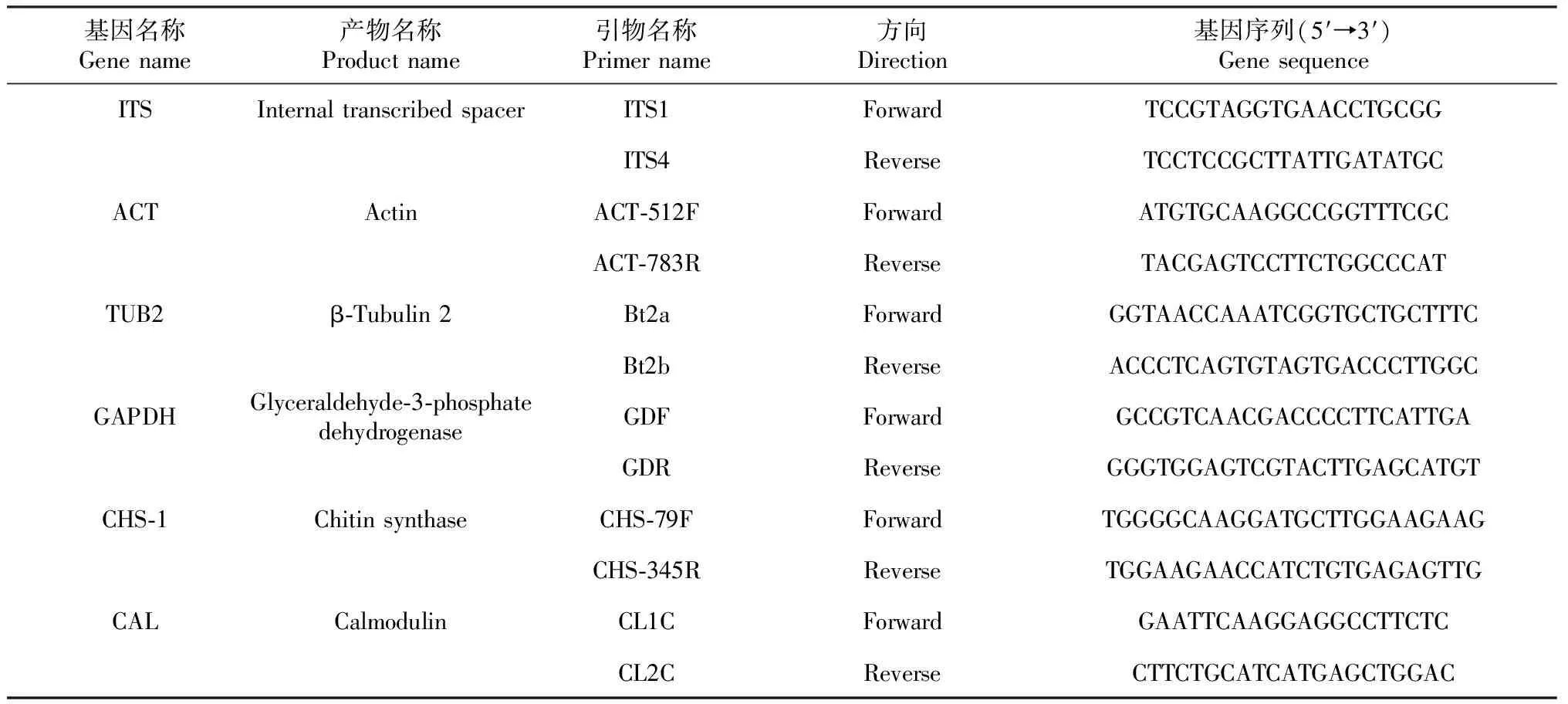
表1 菌株H6基因序列的PCR扩增所用引物
pH对病原菌菌丝生长的影响:将直径为5 mm的菌丝块分别接种至pH为3.5、4.0、 4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5、9.0、9.5、10.0和10.5的PDA培养基中,28.0 ℃培养7 d后用十字交叉法测量各处理的菌落直径,计算菌落扩展直径,筛选最适宜菌株H6菌丝生长的pH。每处理重复3次。
碳源对病原菌菌丝生长的影响:用蔗糖、葡萄糖、山梨醇、D-半乳糖、可溶性淀粉、木糖醇、木糖、甘油、果糖、乳糖和甘露醇等作为碳源,等量代替基础培养基中的碳源(蔗糖),制成含不同碳源的培养基,pH自然。用直径5 mm的打孔器取菌丝块接种于上述培养基中,28.0 ℃培养5 d后用十字交叉法测量菌落直径,计算菌落扩展直径,筛选最适宜菌株H6菌丝生长的碳源。每个处理重复3次。
氮源对病原菌菌丝生长的影响:用硝酸钠、亚硝酸钠、甘氨酸、L-脯氨酸、尿素、硝酸钾、氯化铵、硫酸铵、磷酸二氢铵和DL-丙氨酸等作为氮源,等量代替基础培养基中的氮源(硝酸钾),制成含不同氮源的培养基,pH自然。用直径5 mm的打孔器取菌丝块接种于上述培养基中,28.0 ℃培养5 d后用十字交叉法测量菌落直径,计算菌落扩展直径,筛选最适宜菌株H6菌丝生长的氮源。每个处理重复3次。
杀菌剂对病原菌菌丝生长的抑制效果试验:在PDA培养基中,分别加入1.1.3中的12种杀菌剂,制成含药培养基,不同杀菌剂最终稀释倍数和有效浓度见表2。将直径为5 mm的菌丝块接种至各含药培养基中,28.0 ℃培养5 d后用十字交叉法测量菌落直径,计算菌落扩展直径,分析不同杀菌剂对菌株H6菌丝生长的抑制效果。以不含药剂的PDA培养基为对照(CK),每个处理重复3次。
1.3 统计分析
试验数据采用DPS_V9.01的Duncan’s新复极差法进行单因素方差分析。
2 结果与分析
2.1 红花檵木炭疽病自然症状及病原菌接种症状
红花檵木炭疽病主要危害红花檵木的叶片,植株中下部叶片受害较重,最初在叶片上形成近圆形或不规则形褐色小斑,随后扩展成大斑,或多个病斑融合成更大的病斑,病斑内部为褐色或灰褐色,边缘为深褐色(图1-A)。病害严重时,病斑扩展至整张叶片,造成叶片萎蔫干枯甚至脱落。从不同发病时期的病叶中共分离获得6株形态一致的真菌菌株。将这些菌株的菌饼接种至红花檵木活体叶片上,接种第7天叶片上出现近圆形褐色病斑(图1-B),接种第10天病斑变灰褐色并干枯(图1-C),与自然发病症状相似。接种空白PDA菌饼的CK叶片均未发病(图1-D)。从接种发病的病斑上均能再次分离纯化出相同的病原菌。
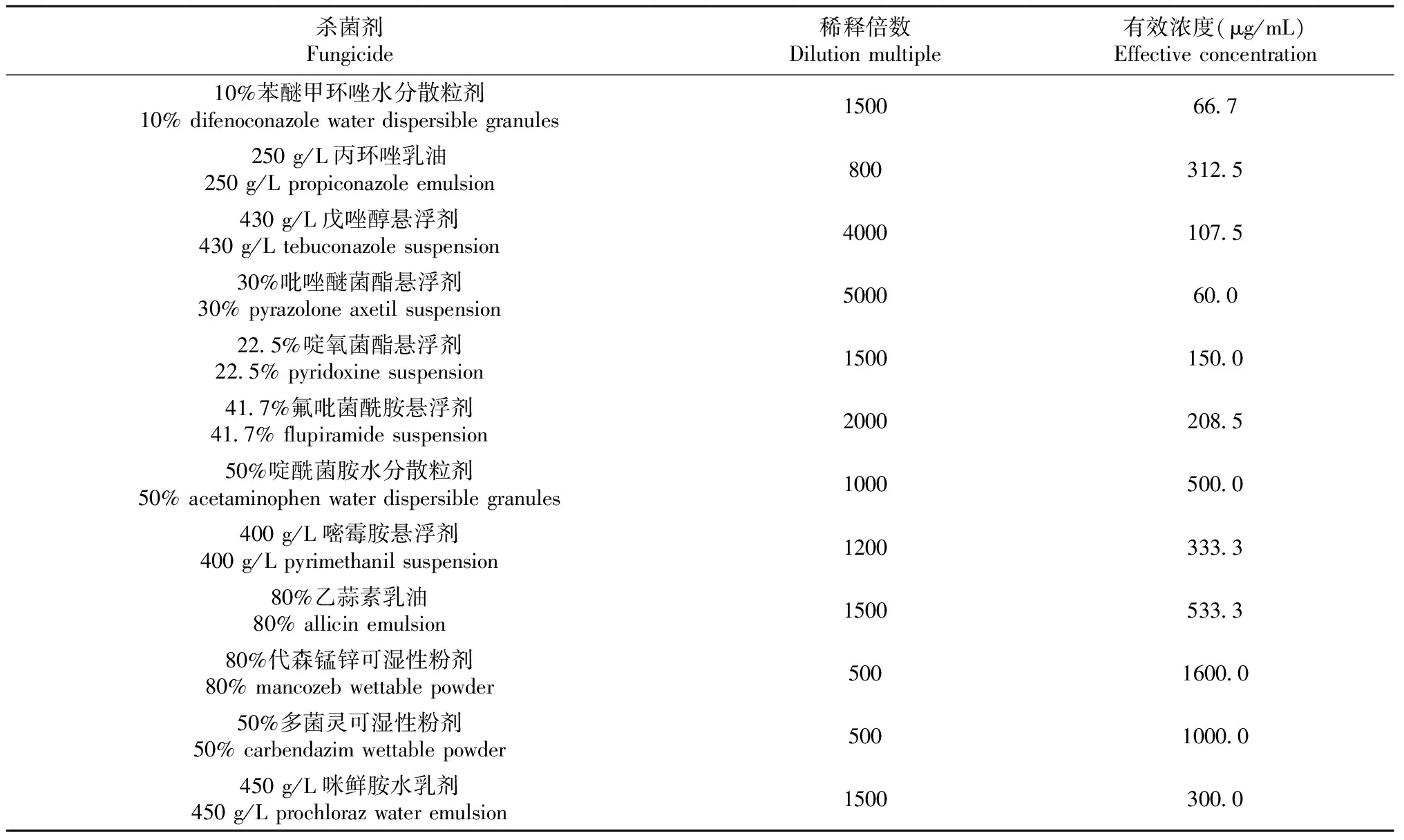
表2 不同杀菌剂最终稀释倍数和有效浓度
2.2 病原菌鉴定结果
2.2.1 形态学特征 在PDA培养基中,6株菌株的菌落均近圆形,初为白色,培养8 d后,菌落中部灰色至深灰色,边缘灰白色,气生菌丝体疏松、絮状(图2-A);分生孢子为单细胞、透明、直、长椭圆形,一端钝圆,另一端钝圆或略尖(图2-B);大小为(8.52~18.83)μm×(4.25~6.03)μm。形态特征与前人[14-15]报道的果生刺盘孢形态一致。
2.2.2 多基因系统发育进化树分析结果 对代表性菌株H6的ITS、 ACT、TUB2、GAPDH、CHS-1和 CAL基因序列进行PCR扩增和测序,所得序列信息提交至GenBank数据库,获得登录号分别为OK327013、OK649310、OK649314、OK649313、OK649312和OK649311。通过BLAST同源性比对,这些基因序列与数据库中多个果生刺盘孢菌株的序列同源性均最高,达99%~100%。以Monilochaetesinfuscans作为外群,采用MEGA-X软件的Neighbor Joining法构建基于ITS、ACT、TUB2、GAPDH、CHS-1和 CAL的多基因系统发育进化树(图3),利用最大似然法(ML)进行系统发育分析,结果发现菌株H6与果生刺盘孢(C.fructicola)聚于同一最小分支。结合形态学和分子生物学鉴定结果,将红花檵木炭疽病病原菌鉴定为果生刺盘孢。

A:自然症状;B:接种病原菌第7天发病症状;C:接种病原菌第10天发病症状;D:空白对照A:Natural symptoms;B:Symptoms of leaves inoculated with the pathogen on the 7th day;C:Symptoms of leaves inoculated with the pathogen on the 10th day;D:Blank control图1 红花檵木炭疽病症状Fig.1 Anthracnose symptoms of L. chinense var. rubrum
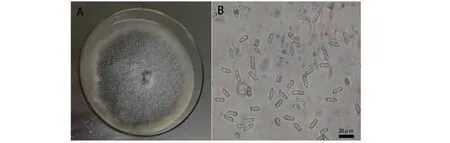
A:PDA培养基上的菌落形态;B:分生孢子形态A:Colony on PDA;B:Conidial morphology图2 病原菌形态特征Fig.2 Morphological characteristics of the pathogen
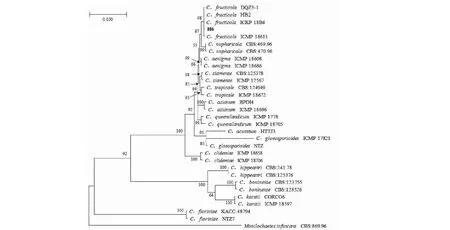
图3 基于ITS、ACT、TUB、GAPDH、CHS-1和CAL基因序列的炭疽菌菌株系统发育进化树Fig.3 Phylogenetic tree of anthrax strains based on the gene sequences of ITS,ACT,TUB,GAPDH,CHS-1 and CAL
2.3 果生刺盘孢的生物学特性
2.3.1 温度对果生刺盘孢菌丝生长的影响 从图4可以看出,果生刺盘孢可在10.0~34.0 ℃下生长,随着温度的升高,菌落的扩展直径呈先上升后下降趋势;25.0~31.0 ℃较适宜菌丝生长,而最适宜温度为28.0 ℃,菌落扩展直径达63.13 mm,且显著高于其他处理(P<0.05,下同);除温度为4.0、7.0和37.0 ℃时菌丝不能生长外,在其他温度下病原菌菌落的扩展直径间均具有显著差异。可见,最适宜果生刺盘孢菌丝生长的温度为28.0 ℃。
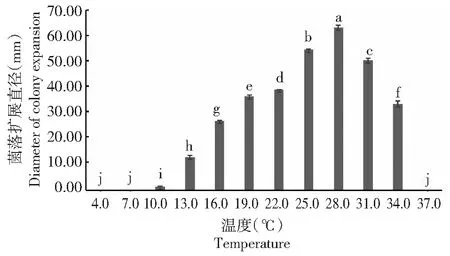
图4 温度对果生刺盘孢菌丝生长的影响Fig.4 Effects of temperature on mycelial growth of C. fructicola
2.3.2 pH对果生刺盘孢菌丝生长的影响 由图5可看出,果生刺盘孢菌丝在pH 3.5~10.5均可生长。其中,pH在4.0~9.5时,菌落均能良好生长,较适宜的pH为6.0~8.5;当pH为8.0时,菌落扩展直径最大,达53.88 mm,与pH 7.5时的菌落扩展直径差异不显著(P>0.05,下同),但均显著大于其他pH处理;pH为6.0、6.5和7.0时,菌落扩展直径间也无显著差异;pH在3.5、10.0和10.5时,菌落扩展直径显著小于其他pH处理。可见,最适宜果生刺盘孢菌丝生长的pH为8.0。
2.3.3 碳源对果生刺盘孢菌丝生长的影响 从图6可看出,果生刺盘孢菌丝在11种供试碳源培养基中均能生长,但在不同碳源之间生长差异明显。其中,在以木糖醇为碳源的培养基中菌落扩展直径显著大于其他处理,其次是山梨醇碳源培养基,再次是甘露醇碳源培养基,而在以D-半乳糖和果糖为碳源的培养基上生长较慢,二者的菌落扩展直径差异不显著,但均显著小于其他碳源培养基。可见,最适宜果生刺盘孢菌丝生长的碳源为木糖醇,山梨醇和甘露醇也可作为较适宜果生刺盘孢菌丝生长的碳源。
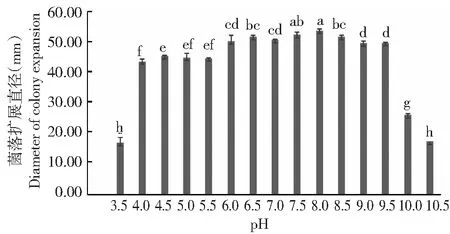
图5 pH对果生刺盘孢菌丝生长的影响Fig.5 Effects of pH on mycelial growth of C. fructicola
2.3.4 氮源对果生刺盘孢菌丝生长的影响 从图7可看出,果生刺盘孢菌丝在10种供试氮源培养基中均能生长,有机氮源与无机氮源均可利用。其中,在以L-脯氨酸为氮源的培养基中菌落直径扩展最快,且显著大于其他处理,其次是在尿素和硝酸钾培养基中,二者菌落的扩展直径差异不显著,而在以氯化铵、硫酸铵和磷酸二氢铵为氮源的培养基中生长较慢,三者间的菌落扩展直径差异不显著,但均显著小于其他处理。可见,最适宜果生刺盘孢菌丝生长的氮源为L-脯氨酸,尿素和硝酸钾也可作为较适宜果生刺盘孢菌丝生长的氮源。
2.3.5 不同杀菌剂对果生刺盘孢菌丝生长的影响 从图8可看出,不同杀菌剂对果生刺盘孢菌丝的生长均具有明显影响。其中,果生刺盘孢菌丝在含有吡唑醚菌酯、啶氧菌酯、乙蒜素、多菌灵和咪鲜胺的培养基中均不能生长,说明这几种药剂对病原菌具有较强的抑制效果;果生刺盘孢菌丝在含有其他药剂的培养基中均能生长,但在含有丙环唑和代森锰锌的培养基中菌落扩展直径较小,仅分别为3.69和4.95 mm,说明这2种药剂对果生刺盘孢也有较好的抑制效果;啶酰菌胺对果生刺盘孢的抑制效果最差,菌落扩展直径与CK无显著差异。可见,吡唑醚菌酯、啶氧菌酯、乙蒜素、多菌灵和咪鲜胺对果生刺盘孢菌丝生长的抑制作用较佳。

图6 碳源对果生刺盘孢菌丝生长的影响Fig.6 Effects of carbon source on mycelial growth of C. fructicola
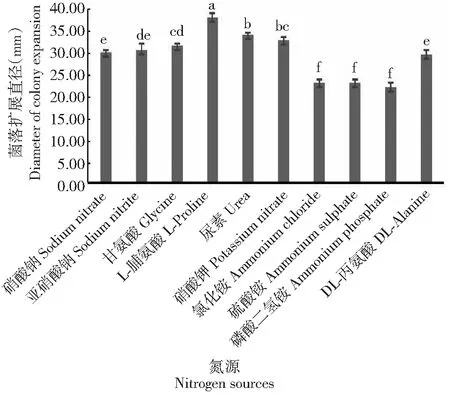
图7 氮源对果生刺盘孢菌丝生长的影响Fig.7 Effects of nitrogen source on mycelial growth of C. fructicola
3 讨 论
本研究结合形态学特征和多基因系统发育分析结果将广西红花檵木炭疽病病原鉴定为果生刺盘孢,与张国辉等[10]对贵州省凯里市红花檵木炭疽病病原鉴定结果为胶孢炭疽菌不一致,表明引起红花檵木炭疽病的病原菌不止1种,在不同地区的优势病原菌种类不同,可能与病原菌的地域适应性有关。此外,在分子生物学鉴定方面,张国辉等[10]仅对ITS序列进行测序,而本研究对ITS、ACT、TUB2、GAPDH、CHS-1和 CAL等多基因序列进行测序并构建多基因系统发育进化树进行分析,因此,本研究的鉴定结果相对更精确。随着分子生物学技术的发展,近年来多数学者均采用形态学与多基因系统发育进化树分析相结合的方法对植物炭疽病病原菌进行种的鉴定[20-23],因为炭疽菌属的分类较复杂,其中,胶孢炭疽菌为复合种。Weir等[19]于2012年通过多基因联合建树的方法将胶孢炭疽菌复合种划分为22个种和1个亚种,由于各复合种的形态特征易受培养条件影响而发生变异,且种间基因序列的相似度较高,仅依靠形态学特征和单一的基因分析鉴定方法不易进行种间区分。本研究鉴定的果生刺盘孢与张国辉等[10]鉴定的胶孢炭疽菌均为胶孢炭疽菌的复合种之一,因此,在分子生物学鉴定方面,采用多基因系统发育分析方法有利于提高鉴定结果的可靠性。此外,胶孢炭疽菌的不同复合种常存在共同侵染现象,如广东的咖啡炭疽病由果生刺盘孢和C.siamense共同侵染,以果生刺盘孢为优势菌[20];成都的洒金珊瑚炭疽病由C.siamense、C.aenigma、C.karstii和果生刺盘孢共同侵染,以C.siamense为优势菌[22]。Qiu等[11]同样结合形态学和多基因序列发育分析方法将红花檵木原种(白花檵木)炭疽病病原鉴定为果生刺盘孢,本研究结果与其一致,表明该菌既能侵染红花檵木也可侵染白花檵木。此外,果生刺盘孢还可侵染咖啡、草莓、洒金珊瑚、甜柿、樱桃和甜茶等多种植物引起炭疽病[20-24],表明该病原菌寄主范围广泛。广西红花檵木炭疽病的病原除果生刺盘孢外,是否还存在胶孢炭疽菌的其他复合种,有待进一步探究。

图8 不同杀菌剂对果生刺盘孢菌丝生长的抑制效果Fig.8 Inhibitory effects of different fungicides on mycelial growth of C. fructicola
本研究结果表明,广西红花檵木果生刺盘孢对温度适应范围广,菌丝在10.0~34.0 ℃时均能生长,适宜生长温度为25.0~31.0 ℃,最适温度为28.0 ℃,温度低于7.0 ℃和高于37.0 ℃均不能生长,与宋丽丽等[21]对草莓果生刺盘孢和刘倩丽等[24]对檀香果生刺盘孢的研究结果基本一致。稍有不同的是,本研究试验设计的温度梯度间隔为3.0 ℃,而宋丽丽等[21]、刘倩丽等[24]设置的温度梯度间隔为5.0 ℃,其测定的果生刺盘孢生长最适温度均为30.0 ℃。
本研究发现,红花檵木果生刺盘孢对pH适应范围也较广,菌丝在pH 3.5~10.5均能生长,适宜生长pH为6.0~8.5,最适生长pH为8.0;红花檵木果生刺盘孢菌丝生长可利用多种碳源和氮源,其中,较适宜的碳源为木糖醇、山梨醇和甘露醇,较适宜的氮源为L-脯氨酸、尿素和硝酸钾,与宋丽丽等[21]、刘倩丽等[24]的研究结果存在差异。原因可能是不同来源菌株对生长条件的需求不同,试验设计和供试的碳氮源不同也可能造成研究结果存在差异。
徐丹丹等[20]报道咪鲜胺和吡唑醚菌酯对咖啡果生刺盘孢菌丝生长具有显著抑制效果,张童等[23]报道百菌清对甜柿果生刺盘孢菌丝生长的抑制效果最好,唑醚·代森联和嘧菌酯次之。本研究发现,吡唑醚菌酯、啶氧菌酯、乙蒜素、多菌灵和咪鲜胺对红花檵木果生刺盘孢菌丝生长具有较强的抑制作用。因此,以上药剂种类均可作为果生刺盘孢的防治药剂在生产中试验使用。
4 结 论
采用传统形态学与现代分子生物学相结合方法,首次明确引起广西园林观赏植物红花檵木炭疽病的病原菌为果生刺盘孢(C.fructicola)。红花檵木炭疽病病原菌菌丝生长受温度、pH、碳源、氮源和杀菌剂等的影响,其中,温度和杀菌剂的影响较突出,杀菌剂吡唑醚菌酯、啶氧菌酯、乙蒜素、多菌灵和咪鲜胺等对红花檵木果生刺盘孢菌丝生长具有较强的抑制作用,制定红花檵木炭疽病防治措施时需充分了解该菌的生物学特性。
