2020年~2021年我国副猪嗜血杆菌和链球菌的血清型、耐药基因型流行情况分析
2022-02-04董婉玉王馨雨张聚民王晴雨周莹珊于浩然宋厚辉王晓杜
董婉玉,王馨雨,张聚民,王晴雨,肖 敏,任 华,卢 昌,周莹珊,杜 静,于浩然,仲 伟,宋厚辉*,王晓杜*
(1.浙江农林大学动物科技学院/动物医学院/浙江省畜禽绿色生态健康养殖应用技术研究重点实验室/动物健康互联网检测技术浙江省工程研究中心/浙江省动物医学与健康管理国际科技合作基地/中澳动物健康大数据分析联合实验室,浙江 杭州 311300;2.杭州市临安区畜牧农机发展中心,浙江 杭州 311300;3.江西正邦农业科学院,江西 南昌 330096)
副猪嗜血杆菌(Haemophilus parasuis,HPS)[1]和猪链球菌(Streptococcus suis,SS)[2]是猪场最常见的细菌性疾病病原,严重危害保育猪的健康,主要导致猪的多发性浆膜炎、脑膜炎、关节炎、心内膜炎、败血症、支气管炎等[3-4],常伴随猪繁殖与呼吸综合征病毒、猪圆环病毒2 型等病毒感染后期的继发感染[5],给养猪业造成重大经济损失。
HPS 是一种兼性厌氧的革兰阴性菌,依据其荚膜多糖基因的不同[6],利用分子生物学技术可以将其分为1~15 个血清型[7],其中优势血清型包括:血清2、4、5、12、13 型等; SS 是一种有荚膜的革兰阳性球菌,由于荚膜多糖基因的差异[8],利用PCR 技术 可 以 将 其 分 为1~34 和1/2 共35 种 血 清 型[9],2014年之前优势血清型包括:2、3、7、8、9、28、29型。HPS 和SS 不同血清型之间的抗原交叉保护性均较弱,不能仅利用某一血清型疫苗做到有效防控HPS 或SS 的感染。因此,临床上常用抗生素治疗和预防性给药,常见兽用抗生素包括β-内酰胺类、氨基糖类、四环素类、大环内酯类、喹诺酮类、甲氧苄啶类、磺胺类和其他抗菌药物,但长期预防性给药易导致细菌耐药性的增强[10-11]。细菌耐药性是由其耐药基因编码的酶所决定,细菌长期在抗生素的压力下,其耐药基因不断突变、耐药基因转导等原因,导致多重耐药菌株不断出现,给养殖场临床用药造成较大困难。
因此,本研究采用PCR 方法检测2020 年~2021年分离自国内10 个省份的HPS 和SS 分离株的血清型和耐药基因,采用药敏试验检测二者的耐药性,并将检测结果与2014 年~2020 年的相应检索结果比较分析,以期为规模化养殖场HPS 和SS 的血清型和耐药基因的研究和相关疾病的防控奠定基础。
1 材料与方法
1.1 分离菌株52株HPS和69株SS由浙江农林大学动物预防医学与公共卫生实验室分离自2020年~2021年国内浙江、江西、湖北、湖南、广东、广西、云南、新疆、辽宁、江苏10 个省的规模化养殖场不同发病仔猪的肺脏,且同一个养殖场均分离到了HPS和SS(表1)。分离纯化的各菌株均于-80 ℃保存。
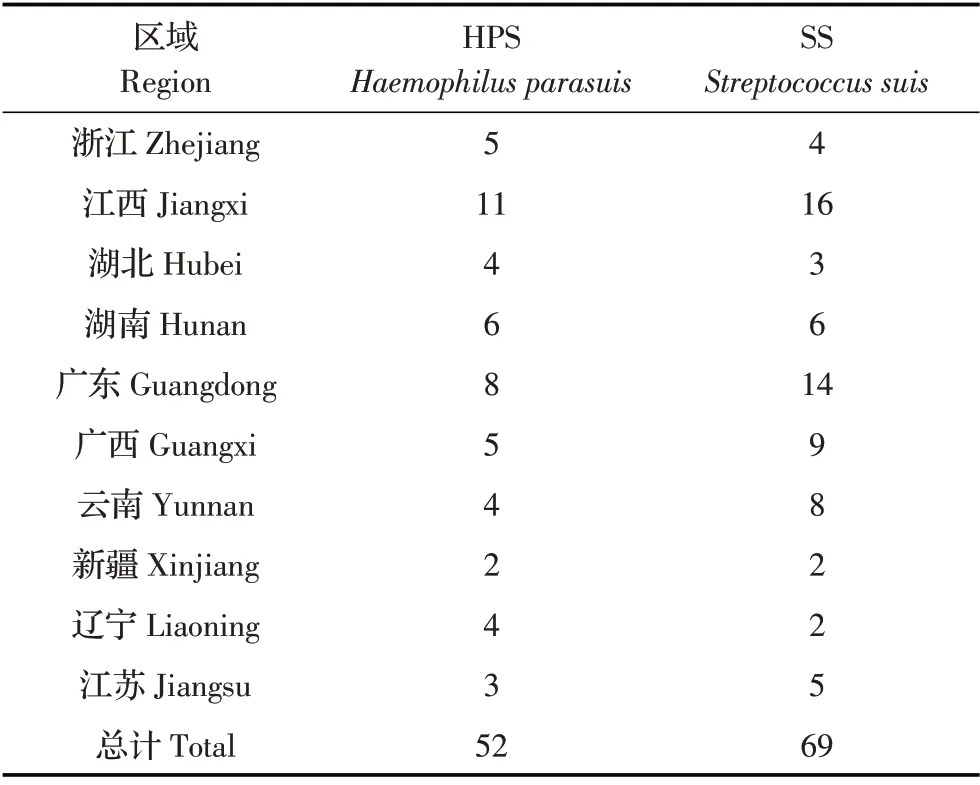
表1 52株HPS和69株SS信息Table 1 Information on 52 isolates HPS and 69 isolates of SS
1.2 主要试剂NAD 辅酶和大豆酪蛋白琼脂培养基(TSA)购自北京索莱宝科技有限公司;胎牛血清购自Thermo Scientific(中国)有限公司;胰蛋白胨大豆肉汤(TSB)购自广东环凯为神武科技有限公司;药敏片购自杭州微生物试剂有限公司;KOD plus Neo购自日本东洋纺生物有限公司;PCR Mix 和细菌基因组DNA 小量提取试剂盒购自生工生物工程(上海)股份有限公司。
1.3 HPS 和SS 分离菌血清型的PCR 鉴定提取HPS 分离株基因组DNA 作为模板,采用文献[12]中PCR 方法检测分离HPS 的血清型;提取SS 分离菌株基因组DNA 作为模板,采用文献[9]中PCR 方法检测分离SS 的血清型。
1.4 HPS 和SS 的药敏试验采用WHO 推荐的Kir⁃by-Bauer(K-B)纸片扩散法分别检测HPS 和SS 对临床常用9 类(β-内酰胺类、大环内酯类、氨基糖苷类、四环素类、喹诺酮类、磺胺类、酰胺醇类、多肽类、林可酰胺类)共计30 种药物的敏感性。
1.5 HPS 和SS 常见耐药基因的PCR 鉴定参照文献[13],由北京擎科生物科技有限公司合成细菌常见耐药基因引物。分别以1.3 提取的HPS 和SS 基因组DNA 为模板, 经PCR 扩增β-内酰胺类(bla-TEM、bla-OXA、bla-CTX、bla-SHV、bla-IMP、bla-DHA)、氨基糖苷类(aad-AI、aad-A2、strA、strB、aadB、aacC、Aph(3')-Ia)、四环素类(tetA、tetB、tetD)、喹诺酮类(gyrA、parC)、甲氧苄啶类(dhfrV、dhfrVII、dhfrXII、dh⁃frXIII)、磺胺类(SulI、SulII)和I 型整合子(3'CS、Int1、VR)等7 大类共27 种耐药基因。
1.6 部分分离菌株耐药表型与耐药基因相关性的分析选取10 个省11 株HPS 和14 株SS 临床分离株进行耐药基因(β-内酰胺类、氨基糖苷类、四环素类、喹诺酮类和磺胺类)与其耐药表型的相关性分析。耐药基因型与耐药表型符合率=(携带耐药基因并具有相应耐药表型的菌株数+不携带耐药基因且不具有相应耐药表型的菌株数)/总菌株数*100%。
1.7 本研究分离的HPS 和SS 与国内已分离HPS 和SS 血清型的比较分析检索2014 年~2020 年国内外发表的402 株HPS[14-17]和189 株SS[9,18]的血清型数据(表2),结合本研究的检测结果,利用Graphad Prism7.0 软件统计分析2014 年~2020 年我国HPS 和SS优势血清型的流行趋势。
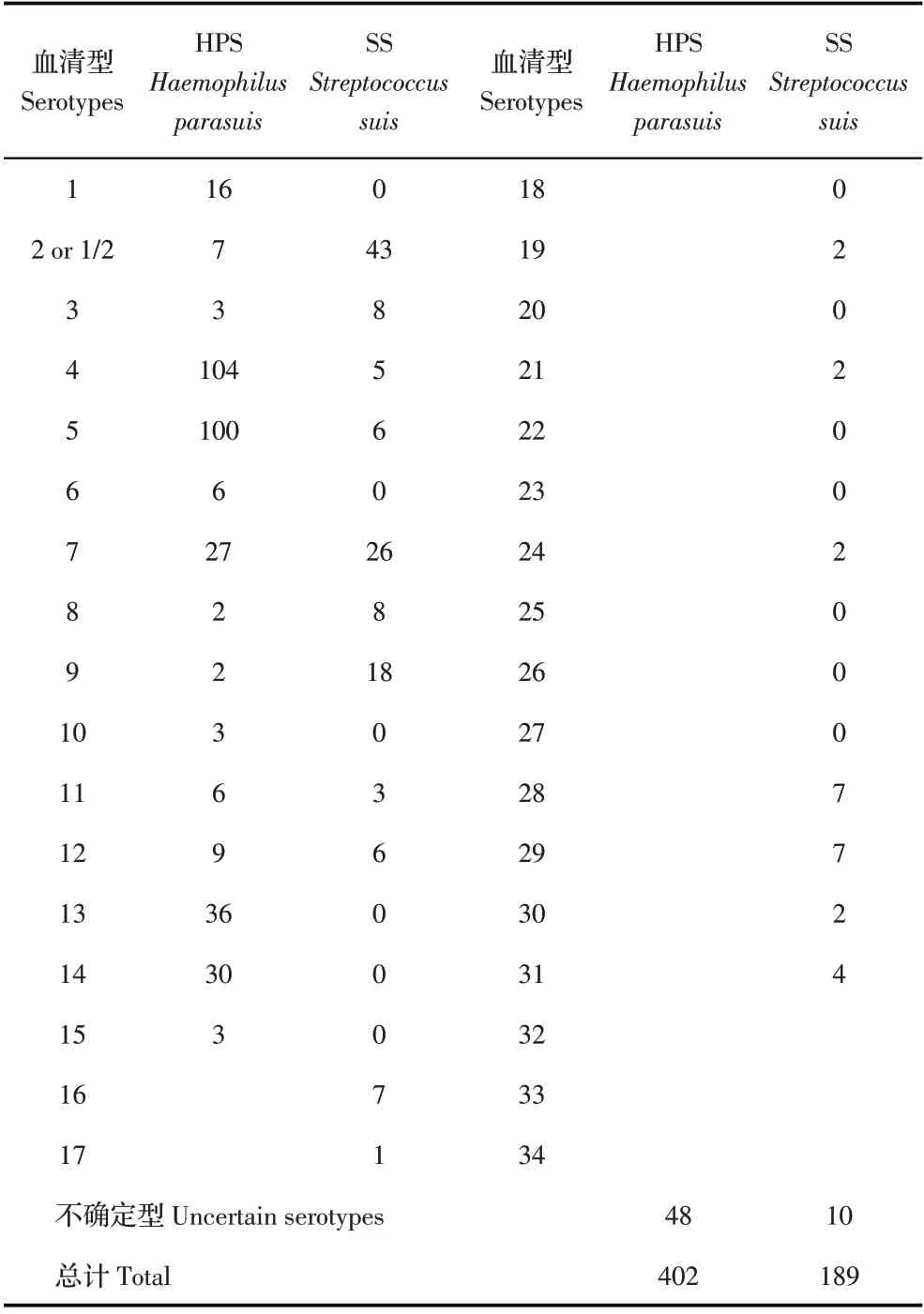
表2 2014年~2020年发表文献的HPS和SS的血清型统计结果Table 2 Statistical results of reported HPS and SS serotypes in 2014-2020
2 结 果
2.1 HPS 分离株血清型的PCR 鉴定及其与文献发表HPS 血清型的比较分析结果利用PCR 方法对52株HPS 进行血清型鉴定并经统计分析,结果显示,优势血清型为为4、5、12、13 型,分别各占15.38%(8/52)、15.38%(8/52)、30.76%(16/52)、11.54%(6/52)(图1)。国内外文献报道2014 年~2020 年的402 株HPS中,优势血清型分别为4、5、7、13、14 型、不确定型(Undetermined serotype,US)分别各占25.90%(104/402)、 24.90%(100/402)、 6.70%(27/402)、8.95%(36/402)、7.50%(30/402)、11.94%(48/402)(图1)。结果表明,血清4、5、13 型仍是HPS 的优势血清型,可以作为临床HPS 灭活疫苗的候选株;与2014 年~2020 年文献报道的相比,2020 年~2021 年HPS 优势血清型中的12 型的流行率较高,且升高了28%,值得重点关注。

图1 本研究分离及文献报道的HPS血清型的统计结果Fig.1 Serotypes analysis of HPS isolated in this study and reported in the literature
2.2 SS 分离株血清型的PCR 鉴定及其与文献发表SS 血清型的比较分析结果利用PCR 方法检测66 株SS 分离株的血清型。结果显示,主其要优势血清型为2 或1/2、5、8、15、18、27 和US,分 别 占23.18%(16/69)、17.39%(12/69)、10.14%(7/69)、7.24%(5/69)、10.14%(7/69)、8.69%(6/69)、11.65%(8/69)(图2)。国内外文献报道2014 年~2020 年我国SS 流行的优势血清型分别为2 或1/2、7、9 和US,分别占22.75%(43/189)、13.75%(26/189)、9.52%(18/189)、5.29%(10/189)(图2)。结果表明,2 或1/2 仍然是我国SS的优势血清型;相比2014 年~2020 年的文献报道,2020 年~2021 年SS 优势血清型发生了变化,5、15、18、27 型明显增加,7 和9 型减少;US 仍较多,是否将其确定为新型需要进一步研究。
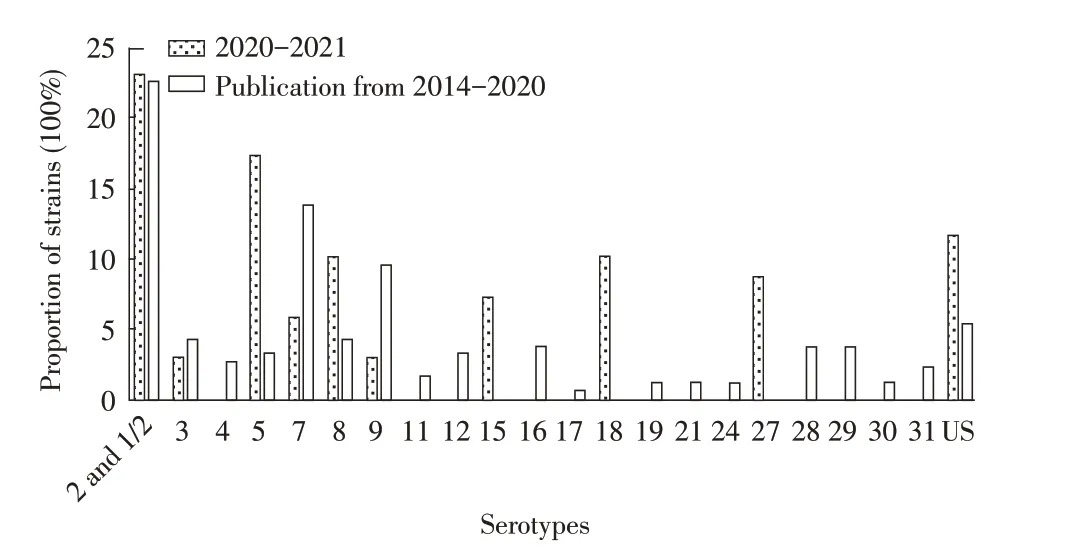
图2 本研究分离及文献报道的SS血清型统计分析Fig.2 Serotype analysis of SS isolated in this study and reported in the literature
2.3 HPS 和SS 分离株的药敏试验结果药敏试验结果显示,两类分离菌中对β-内酰胺类(β-lac⁃tams)、大环内酯类(Macrolides)和磺胺类(Sulfon⁃amides)药物耐药高达88.31%(68/77),对氨基糖苷类(Aminoglycosides)和四环素类(Tetracyclines)耐药的分离株相对较高,分别为80.52%(62/77)和76.62%(59/77)。对酰胺醇类(Amide alcohols)和喹诺酮类(Quinolones)药物有一定程度的耐药,耐药菌株分别为11.69%(9/77)和32.47%(25/77)(图3A)。采用Graph⁃pad 软件分析HPS 和SS 分离株的多重耐药性。结果显示,HPS和SS临床分离株存在严重的多重耐药性,其中双重耐药的菌株占比最高(28.57%,22/77),分离菌株最多可同时耐8 种药物。其中4 重、5 重、6 重及8重耐药的菌株均为14.29%(11/77)(图3B)。上述结果表明,HPS 和SS 均存在同一菌株多重耐药现象,其中对β-内酰胺类、大环内酯类和磺胺类药物耐药的菌株最高。建议在临床上通过细菌体外分离培养与鉴定病原后,结合药敏试验结果有针对性的使用抗菌药物。

图3 HPS和SS分离株的药敏试验(A)及多重耐药分析(B)Fig.3 Antimicrobial susceptibility(A)and multidrug resistance analysis(B)of HPS and SS isolates
2.4 HPS 和SS 分离株耐药基因的检测结果利用PCR 分别扩增HPS 和SS 分离株7 大类27 种耐药基因,结果显示,两类分离菌均100%检出β-内酰胺类的bla-TEM和磺胺类的sulI耐药基因;100%HPS 分离株检出氨基糖苷类的Aph(3')-Ia、I 类整合子的3'CS和Int1耐药基因;超过50%的HPS 分离株检出氨基糖苷类的strB和磺胺类的sulII耐药基因。100%SS 分离株检出I 类整合子的VR耐药基因;约50%的SS 分别检出I 类整合子的3'CS和Int1耐药基因,且均为不同菌株中检出该两种基因,其他耐药基因均未在两类分离菌中检出(图4)。虽二者均未检测到喹诺酮类和四环素类耐药基因,但二者的部分分离菌株却对这两类药物中的氧氟沙星、环丙沙星、加替沙星、四环素、米诺环素耐药。且在HPS 和SS 中均存在同一菌株携带不同类型耐药基因的现象,再次表明这两类细菌均存在多重耐药现象;β-内酰胺类和磺胺类药物耐药基因检出率高,表明临床针对这两类药物耐药严重,需科学合理用药。
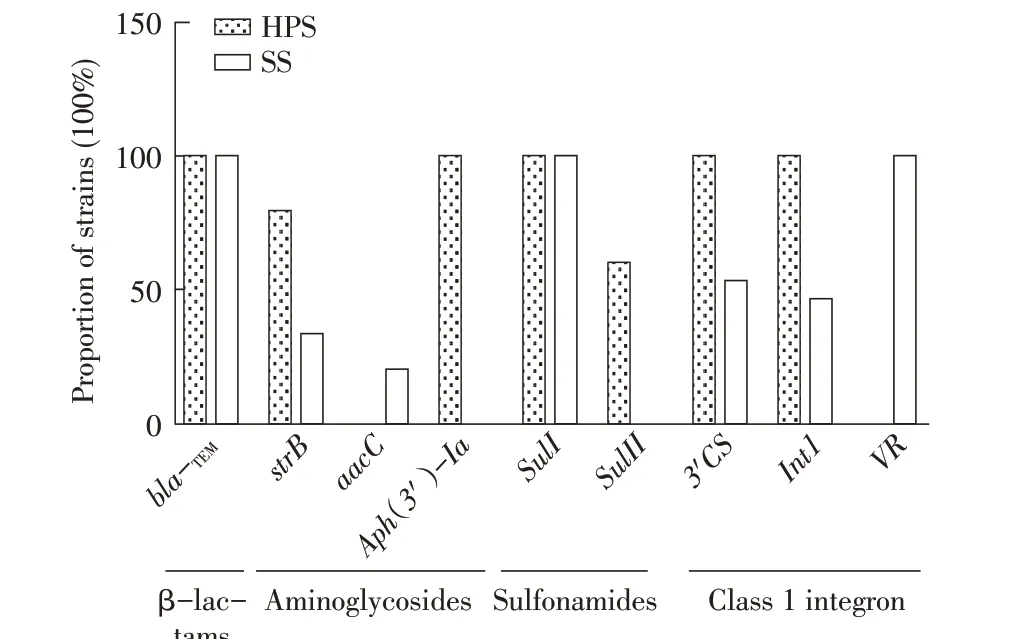
图4 HPS和SS耐药基因的检测结果Fig.4 Detection of drug resistance genes in HPS and SS isolates
2.5 HPS 和SS 分离株耐药表型和耐药基因相关性的分析本研究分别选取10 个地区共约25%的HPS和SS 分离株,分析二者耐药表型和耐药基因的相关性。结果显示,β-内酰胺类和磺胺类耐药表型与其耐药基因之间的符合率最高,均为88%;氨基糖苷类耐药表型与其耐药基因之间的符合率为72%;其他耐药基因和耐药表型之间不符合(表3)。表明HPS和SS 分离株对β-内酰胺类、磺胺类和氨基糖苷类药物的耐药性,可能是由其携带的耐药基因导致;但二者对四环素类和喹诺酮类药物的耐药性可能还存在其他的耐药机制。
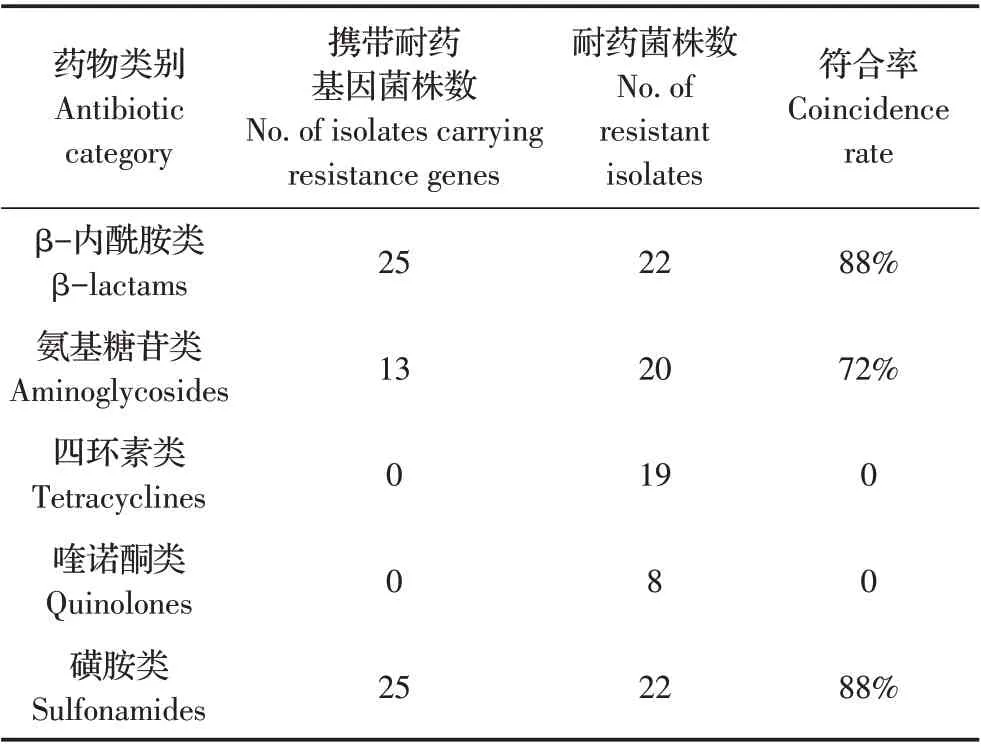
表3 部分HPS和SS分离株耐药表型和耐药基因相关性的分析结果Table 3 Analysis of correlation between of drug resistance phenotype and genotype of partly HPS and SS isolates
3 讨 论
自从2018 年我国暴发非洲猪瘟后,我国养猪产业人员的生物安全意识和水平得到极大了提升,疫病防控水平明显提高[19],但随着国家推行抗生素减量化行动,猪场饲料禁止添加抗生素,导致猪场细菌类疾病发病率上升,致保育猪和中大育肥猪死亡率明显升高。而导致中大猪死亡的细菌类病原主要为HPS 和SS 等,因此针对该类病原感染的防控是目前的热点之一。
HPS 15 个血清型可以被划分为:高毒力型为1、5、10、12、13、14 型;中等毒力型为2、4、8、15型;3、6、7、9、11 型为无毒力型。国外流行的HPS 优势血清型主要为1、2、4、5、12、13 型[20-22],2007 年~2014 年我国HPS 的优势血清型为2、4、5、12、13 型等[7],2014 年~2020 年为4、5、7、13、14型和US[14],7 型作为无毒力型,也能从发病猪中分离到,间接说明HPS 可能是一种条件致病菌[23]。本研究结果显示,2020 年~2021 年HPS 的优势血清型为4、5、12、13 型,特别是12 型的再次出现,值得关注。
SS 作为猪上呼吸道的常在菌,常与猪繁殖与呼吸综合征病毒、猪圆环病毒2 型、猪流感病毒等混合感染,导致猪的发病率和死亡率明显升高[24]。研究表明,虽然血清型不是SS 致病性的唯一因素[25],但是血清2 型、9 型SS 仍是导致欧洲和美国猪发病的优势血清型[26];2014 年之前我国流行的SS 优势血清型为2、3、7、8、9、28、29 型[9],而2014 年~2020 年则为2 或1/2、7、8、9 和US 型[9,18],比国外更加复杂。随着我国非洲猪瘟区域防控措施的实行及猪引种和流动的变化,2020 年~2021 年我国SS 优势血 清 型 为2 或1/2、5、8、15、18、27 和US,5、15、18、27 型的流行率升高。
研发流行优势血清型的细菌多联多价疫苗是防控HPS 和SS 感染的有效措施,尤其是针对怀孕母猪开展优势血清型细菌疫苗的免疫,将有效降低HPS和SS 感染的发病率。然而,猪场仍主要以抗生素防治HPS 和SS 的感染,导致抗生素滥用现象严重,细菌耐药性逐年增加,特别是多重耐药菌株的出现,致猪场能用的抗生素种类逐步减少。本研究药敏试验统计结果表明,HPS和SS中均存在多重耐药现象,对β-内酰胺类、大环内酯类和磺胺类药物耐药的菌株占比最高。Zhao 等的研究结果显示,75%以上的HPS 存在耐药性,对萘啶酸耐药的菌株占比达82.52%,相关耐药基因的检出率达100%,其中bla-TEM-1、bla-ROB-1、ermB、ermA、flor、catl、tetB、tetC、rmtB、rmtD、aadA1、aac(3')-llc、sulI、sulII等14 种耐药基因均被检出[11],而本研究的两类细菌中耐药基因bla-TEM和sulI均100%检出。SS 则对3 种及以上抗生素耐药[10],耐药基因ermB和tetO与其对林可酰胺、大环内酯和四环素类药物的耐药性密切相关;耐 药 基 因aad(6)、ant(6)-Ia、ermB、tetO、patB、sat4、aph(3')-IIIa、aac(6')-Ie-aph(2'')-Ia与其对氨基糖苷类、大环内酯类和氟喹诺酮药物的耐药性相关[27]。本研究结果显示,100%的HPS 分离株均检出耐药基因Aph(3')-Ia、3'CS、Int1,超过50%的HPS 检出耐药基因strB和sulII,其中磺胺类耐药基因检出结果与上述研究结果相似,其他耐药基因检出结果有差异可能是由于细菌对某种或者某几种药物耐药导致养殖场应使用了新药,从而诱发产生了新的耐药基因;100%的SS 检出耐药基因VR,约50%的SS 分别检出耐药基因3'CS和Int1。与上述研究结果不同,可能是由于SS 分离株来源于不同的养殖场,而不同养殖场的用药背景有差异所致。
通过对部分分离株耐药基因和耐药表型的相关性分析显示,两类细菌对四环素类和喹诺酮类药物的耐药基因和耐药表型之间的符合率较低,其原因可能是菌株存在未检测的耐药基因;而两类细菌对β-内酰胺类、磺胺类和氨基糖苷类药物的耐药表型与耐药基因型之间的符合率较高,表明,这些菌株的耐药表型可能主要是由这些耐药基因所介导。本研究结果还显示,有些菌株携带相关耐药基因,但却未表现出对应的耐药表型,推测可能是这些菌株的耐药基因处于沉默状态需要在许多不同的遗传因素刺激下才会表达[28];而一些菌株虽表现出相应耐药表型却并未检测到对应的耐药基因,推测该菌株的耐药性可能并非由该基因决定,或许还存在其他未检测的耐药基因及其他导致该耐药表型的耐药机制。
综上所述,本研究分析了2020 年~2022 年我国HPS 和SS 的流行优势血清型,检测了二者的耐药表型及相关耐药基因,并证明其部分耐药表型与其携带的耐药基因呈较强的相关性。为规模化养殖场HPS 和SS 耐药基因变化规律的研究、临床用药以及疫苗的研发提供参考。
