不同土地利用方式对宁夏盐渍化土壤细菌群落的影响
2022-01-26陈晓娟锐2
李 明,马 飞,陈晓娟,王 锐2,*
(1 宁夏大学 食品与葡萄酒学院,银川 750021;2 宁夏大学 葡萄与葡萄酒研究院,银川 750021;3 宁夏大学 生态环境学院,银川 750021;4宁夏大学 农学院,银川 750021)
作为土壤生态系统中最活跃的部分,土壤微生物通过分解动植物残体参与土壤微生态的营养物质循环和能量传递,是衡量土壤肥力的重要指标之一[1]。土壤细菌作为土壤微生物中最大的类群,约占微生物生物量的70%~90%,可调节土壤结构与肥力的稳定性、养分利用效率和植物生长发育等[2]。同时,土壤细菌比较敏感,其群落结构组成及多样性易受土壤类型、土壤理化性质、施肥方式、耕作方式和植被类型等多方面因素的影响[3]。已有研究结果证实土地利用方式的改变会明显影响土壤微生物活性、代谢等,比如少免耕[4]、轻耕[5]、旋耕[6]、翻耕和深松耕[7]等耕作措施能不同程度改变土壤微生物的生物学特性。土地利用方式的改变会影响土壤养分的循环与供应,直接造成土壤质地和地下微生物群落结构的变化[8],进而引起土壤微生物多样性的改变。
为了消减作物连坐障碍,前人从改变耕作制度入手,做了大量研究,并一致认为:轮、间和套作系统可通过维持作物根际土壤微生物群落结构的多样性,抑制连作条件下的有害微生物增殖,进而增加有益微生物的数量和活性,提高作物产量[9]。比如西瓜和水稻轮作、脆甜豆与胡麻和小麦轮作[10],可通过改善土壤微生态环境,减少土壤虫害、降低作物病害;不同作物与黄瓜套作,可明显提高土壤微生物群落的均匀度和多样性指数,进而改善根际土壤微生态环境和系统[11]。因此,轮作、间作和套作等多样性栽培模式的研究、推广和应用,对缓解和消减作物连作障碍,改良盐渍化土壤都具有重要的理论和现实意义。
土壤盐渍化是一个世界性问题,也是土壤退化和肥力下降的重要表现,不仅造成资源的浪费,而且给生态环境和社会经济的发展带来严重的危害,成为制约区域生态环境与农业高效利用的重要因素[12]。宁夏地处内陆,是中国西北最主要的农业区与生态脆弱区,由于气候干旱,蒸发强烈,地势低洼,成土母质含盐量及地下水位较高等原因以及长期不合理的耕灌制度,使得区域盐碱地面积和盐渍化程度逐年增加,土壤盐渍化已成为制约宁夏地区土地资源利用的主要障碍因子[13]。开展轮作、连作及其相互结合等利用方式对盐渍化土壤微生物群落及土壤理化性质影响的研究是有效开展区域盐渍化土壤资源改良利用的基础。不同土地利用方式可以显著影响土壤细菌的群落结构及土壤理化性质,能够敏感地反映土壤微生态特征的变化,也可以作为宁夏盐渍化土壤改良和土地高效利用的指标之一。
本研究选取宁夏平罗盐渍化土壤,基于高通量测序结合常规土壤理化性质测定方法,开展不同土地利用方式对土壤细菌群落和土壤理化性质影响的分析,旨在揭示不同土地利用方式下,土壤微生物群落变化规律及其主要的影响因素,拟为该地区盐渍化土壤利用方式的优化和利用潜力的提高提供理论参考,促进盐渍化后备耕地资源的合理开发与高效利用。
1 材料和方法
1.1 研究区概况
宁夏银北地区分布有大量盐渍化土壤,是黄河中上游灌溉地区,该区域地势平坦,属干旱的暖温带季风气候,年均降水量为150~210 mm,年均温9.5 ℃,年蒸发量为1 875 mm,地下水埋深1.0~1.2 m,地下水主要含氯化物、硫酸盐。本研究选取宁夏银北平罗西大滩水稻和玉米种植区为试验区,采样信息见表1。土壤类型以盐土、碱土、灌淤土等为主,土壤透气和通水性差。原始荒地土壤理化性质为:pH 9.31、电导率10 970.00 μs/cm、有机碳6.83 g/kg、全氮0.73 g/kg、碱解氮21.89 g/kg、全磷587.53 μg/g、速效磷10.43 mg/kg、速效钾281.31 mg/kg。
1.2 土壤样品采集
2020年7月26日在银北西大滩试验样地采集土壤样品。本研究共设5个处理:(1)将原始盐碱荒地作为对照(CK);(2)水旱轮作12年玉米地(每2年倒茬1次,PD12Y);(3)连续种植12年水稻地(P12Y);(4)水旱轮作12年水稻地(每2年倒茬1次,DP12Y);(5)连续种植12年玉米地(D12Y)。每个处理3次重复。
根据地面作物的生长情况,选择5个30 m × 30 m的小区。在每个小区内,按“S”型选择6个采样点,在每个采样点用直径30 mm的不锈钢土钻钻取5个深度为0~20 cm的土柱,去除小石块和粗根,混合作为一个土壤样品,总共采集15个土壤样品,立即用冰盒将土壤样品带回实验室。采回的土壤样品均分为两部分:一部分于-20 ℃保存,用于土壤微生物DNA的提取;另一部分风干后用于测定土壤理化性质。
1.3 土壤理化性质的测定
将风干后的土壤磨细,分别过2 mm和0.25 mm筛,测定0~20 cm土层土壤有机质、碱解氮、有效磷、速效钾、全氮和全磷含量,具体测定方法参照鲍士旦《土壤农化分析》(第三版)[14]。有机质采用重铬酸钾容量-外加热法测定;碱解氮采用碱解扩散法测定;有效磷采用钼锑抗比色法测定;速效钾采用火焰光度法测定;全氮采用凯氏定氮法测定;全磷采用钼锑抗比色法测定。土壤机械组成采用比重计法测定:根据国际制土壤粒级划分标准将土壤颗粒级别划分为粘粒(<0.002 mm)、粉砂(0.002~0.02 mm)、砂粒(0.02~2 mm)。
1.4 土壤DNA提取
不同处理土壤样本中基因组DNA提取采用QIAamp DNA Stool Mini Kit试剂盒(QIAGEN,Hilden, German),提取方法按照试剂盒说明书进行,使用1.2%琼脂糖凝胶电泳检测抽提的基因组DNA质量合格后由诺禾致源Illumina HiSeq 2500平台进行测序。
1.5 nova-seq PE250
以1%的凝胶浓度、170 V电压和30 min电泳时间为基础条件,进行如下的25 μL反应体系和扩增程序。其中对细菌16S V3-V4区域扩增的引物为338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)。
按指定测序区域(细菌16S V3-V4),合成带有barcode的特异引物,或带有错位碱基的引物。
全部样本按照正常试验条件进行,每个样本3次重复,将同一样本PCR产物混合,2%琼脂糖凝胶电泳检测,AxyPrepDNA凝胶回收试剂盒(Axygen)切胶回收PCR产物,Tris HCl洗脱;2%琼脂糖电泳检测。通过Illumina HiSeq 2500平台进行测序(北京诺禾致源公司http://www.novogene.com/index.php, Beijing, China)。

表1 采样地信息
1.6 数据统计与分析
对Illumina HiSeq测序结果序列进行质控、过滤和拼接,得到优化序列。利用USEARCH软件将优化序列在97%相似度下进行聚类,得到各样品OUT代表序列。参照文献[15]的方法,计算覆盖度(Coverage,反映测序结果是否代表样本的真实情况)。利用mothur(Version1.33.3)进行alpha多样性分析:Shannon、Simpson等估算样品中微生物物种多样性、Chao-1指数估算样品中微生物物种总数、ACE估算样品中微生物物种总数。
Venn图绘制,Beta多样性分析(PCA),主成分分析(PCA),最大程度反映样本间差异的一种应用方差分解。运用USEARCH和mothur软件将OTU代表序列与Silva128数据库进行物种分类学分析。采用R语言的Vegan软件对土壤优势细菌门群落和土壤环境因子进行冗余度分析。
使用Excel 2017对土壤不同组样品的氮磷钾、有机质等土壤理化性质数据进行平均值、标准差计算。对土壤不同组样品的氮磷钾、有机质的数据进行平均值、标准差进行计算。文中组间差异均使用SPSS 20.0软件LSD法计算(P<0.05表明组别之间无统计学差异;P>0.05则表明组别之间无统计学差异),数据表示方式均为“平均值±标准差”。
2 结果与分析
2.1 不同土地利用方式下土壤基本理化性质
与对照相比,全氮、碱解氮、全磷在PD12Y和D12Y处理中最高,其中碱解氮D12Y处理中显著高于其他处理组(P<0.05),全磷在PD12Y处理组最高,达到显著水平(表2)。速效磷、有机质在PD12Y处理组显著高于其他处理组(P<0.05);速效磷在PD12Y处理组中显著高于其他处理组(P<0.05)。同时,土壤pH、电导率值、含水量在PD12Y处理组最低,其中,pH值差异达到显著水平(P<0.05)。砂粒所占比例在P12Y和D12Y中增加,分别增加了8.18%和9.94%;粉沙所占比例在PD12Y、P12Y、DP12Y和D12Y中都增加,其中在D12Y中增加显著,为33.54%;粘粒所占比例在PD12Y、P12Y、DP12Y和D12Y中都降低,其中D12Y下降最为显著,为52.27%。
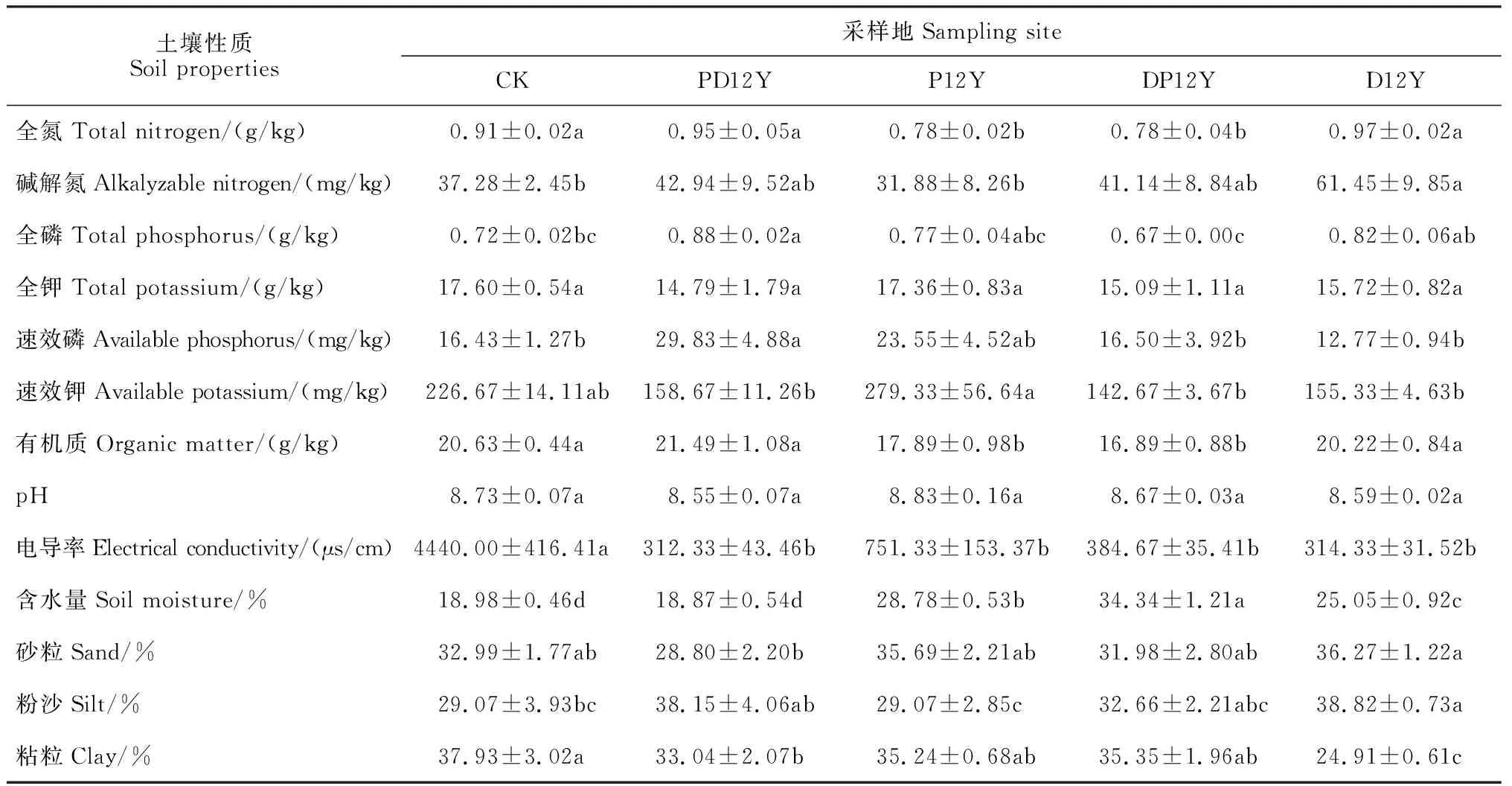
表2 不同土地利用方式土壤基本理化性质
2.2 不同土地利用方式下土壤细菌操作分类单元(OTU)聚类分析
通过不同土地利用方式下盐渍化土壤细菌OTU的聚类可知(图1),土壤样品中共检测到细菌24 400种,5种处理方式的共有种类为1 422种,约占到总数的5.80%。DP12Y拥有的特有性细菌最多,为883种;其次为连续种稻12年的土壤,有697种;特有性细菌最少的处理为连续种植12年玉米地的旱地土壤。同样,DP12Y所测得细菌种类也最多,并高于其他4个处理,为5 570种。其他4个处理细菌丰富度由高到低为P12Y(5 475种)、D12Y(5 180种)、PD12Y(4 877种)和CK(3 298种)。表明种植水稻相对于原始荒地和种植玉米对土壤细菌群落结构影响较大。

图1 不同土地利用方式盐渍化土壤细菌OUT聚类图Fig.1 OUT cluster diagram of bacteria in salinized soil under different land use patterns
2.3 不同土地利用方式下门水平的丰度变化
结果表明,在5种不同土地利用方式下土壤微生物中相对丰度排名前十的优势细菌分别为变形菌门(Proteobacteria),其相对丰度占总细菌群落的21.25%~46.87%;其次为芽单胞菌门(Gemmatimonadetes),其相对丰度占7.10%~25.36%;放线菌门(Actinobacteria)为第三大优势类群,占5.30%~18.87%;其余依次为拟杆菌门(Bacteroidetes)>酸杆菌门(Acidobacteria)>绿弯菌门(Chloroflexi)>未鉴别古菌(Unidentified Archaea)>厚壁菌门(Firmicutes)>疣微菌门(Verrucomicrobia)>硝化螺旋菌门(Nitrospirae)(图2)。

图2 不同土地利用方式盐渍化土壤细菌各门类相对丰度Fig.2 Composition and relative abundance of the bacterial community at the phylum level in the salinized soils with different cultivation methods
2.4 不同土地利用方式下属水平的丰度变化
由表3可知,unidentifiedActinomarinales、Salinimicrobium和Candidatusentotheonella等的相对丰度在几种不同耕作处理的土壤样品中显著地下降(P<0.05)。其中,耕作使得unidentifiedActinomarinales相对丰度显著下降,相对于CK处理,PD12Y处理下降最多为98.28%;Candidatusentotheonella相对丰度在DP12Y处理中下降最多,为89.32%。硫杆菌属(Thiobacillus)、硫碱螺旋菌属(Thioalkalispira)、亚硝化螺菌属(Nitrosospira)等的相对丰度在几种耕作处理中相对于CK显著上升(P<0.05)。
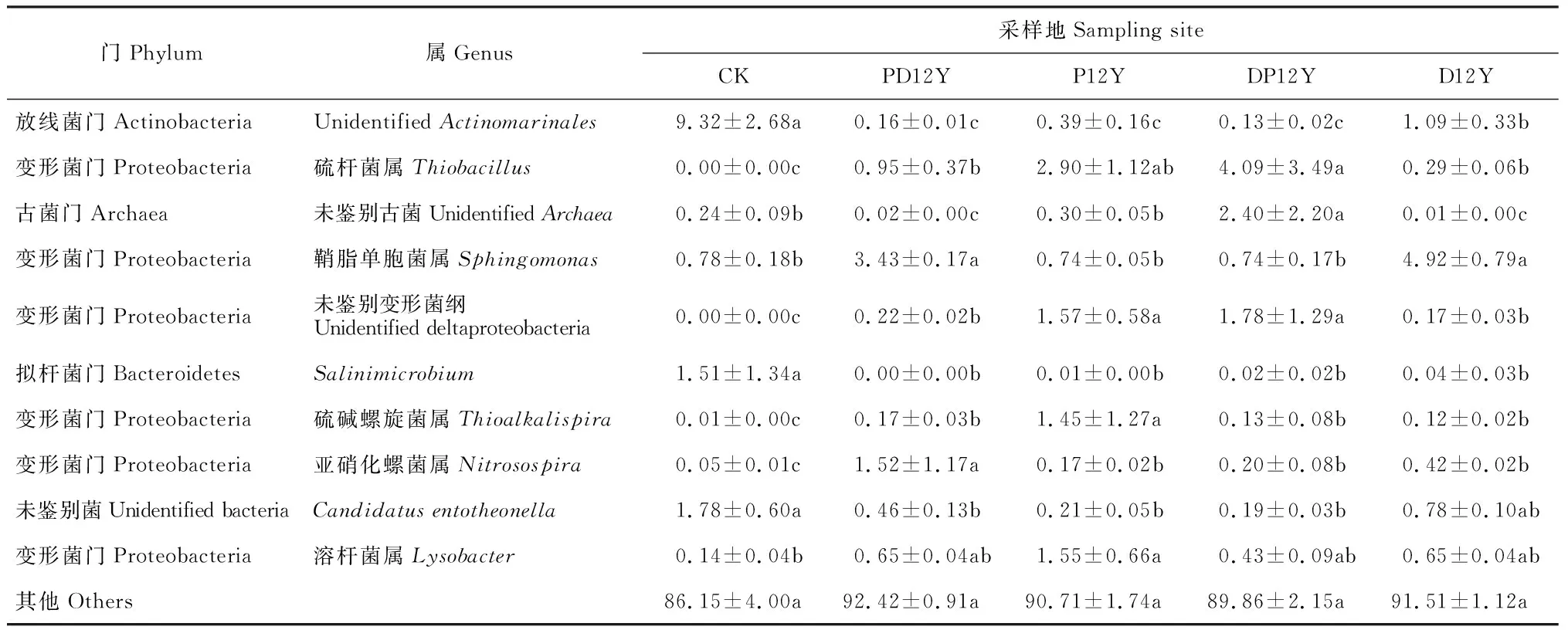
表3 不同土地利用方式土壤细菌群落在属水平上的丰度变化
2.5 Alpha多样性指数及其与土壤理化性质的相关性分析
对不同样本在97%一致性阈值下的Alpha diversity分析指数进行统计分析(表4)。如表所示,P12Y样品的Shannon、Chao-1和ACE指数都高于其他处理组,但没有达到显著水平。observed species、Shannon、Simpson、Chao-1和ACE指数在CK样品中显著低于其他处理组(P<0.05)。

表4 不同土地利用方式土壤样品alpha多样性分析
Pearson相关性分析(表5):Shannon、Simpson、Chao-1、ACE指数均与电导率值呈显著负相关(P<0.05),Shannon指数与土壤含水量呈显著正相关(P<0.05)。
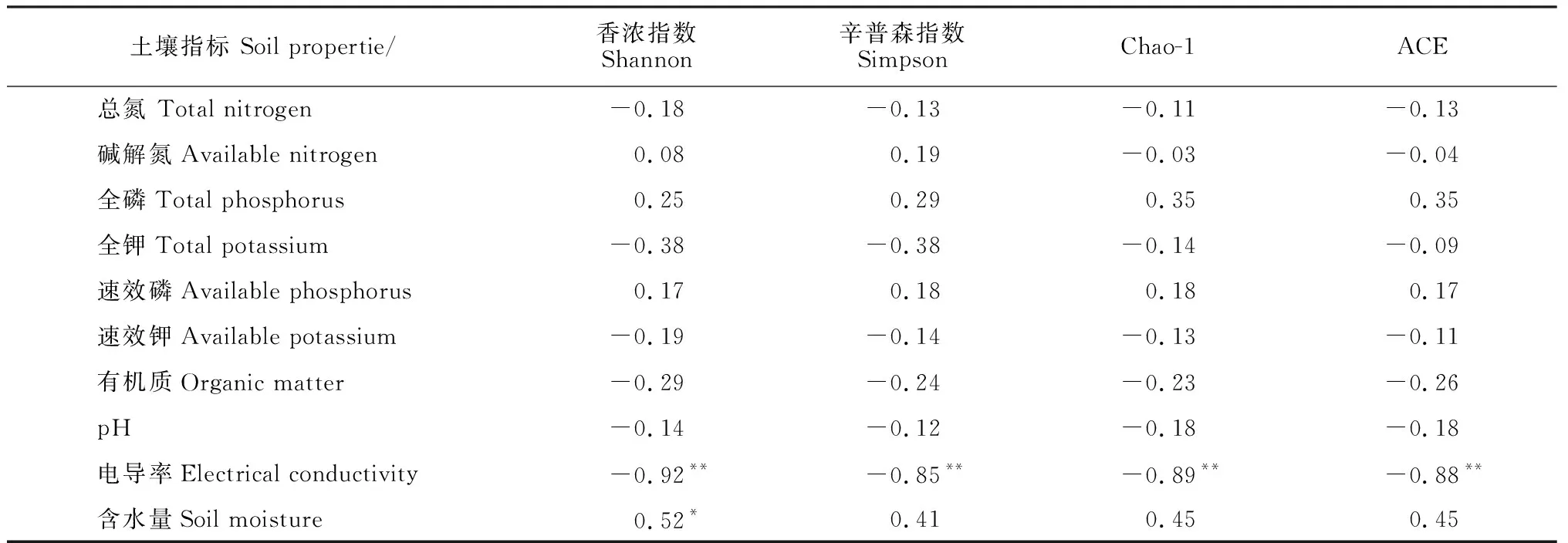
表5 不同土地利用方式土壤样品alpha多样性与土壤理化性质的Pearson相关性分析
2.6 PCA分析和UPGMA聚类分析
第一主成分对样本差异的贡献率为16.15%,第二主成分为12.95%。由图3可见,CK与其他耕作方式的土壤相距较远,表明其细菌群落结构与其他处理组的土壤差异较大。D12Y与PD12Y距离较近,因此D12Y和PD12Y处理组的细菌群落结构较为相似。DP12Y与P12Y相距较近,表明厌氧环境中的细菌群落结构较相似。
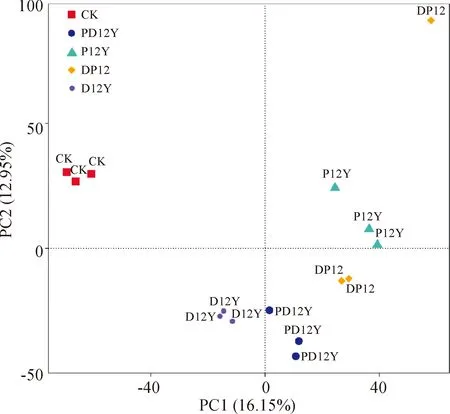
图3 不同土地利用方式下盐渍化土壤细菌群落结构的主成分分析Fig.3 Principal component analysis of bacterial community composition of salinized soils with different cultivation methods
相对丰度排名前十的物种组成分布的样本聚类树如图4可见:CK处理与其他的4个处理组差异较为显著,单独聚为一类。连续种植12年玉米地(D12Y)和水旱轮作12年(PD12Y)的土壤中优势细菌群落聚在一类PD12Y和D12Y。P12Y和DP12Y土壤中优势细菌群落聚在一类。UPGMA聚类分析和主成分分析呈现的结果基本一致。
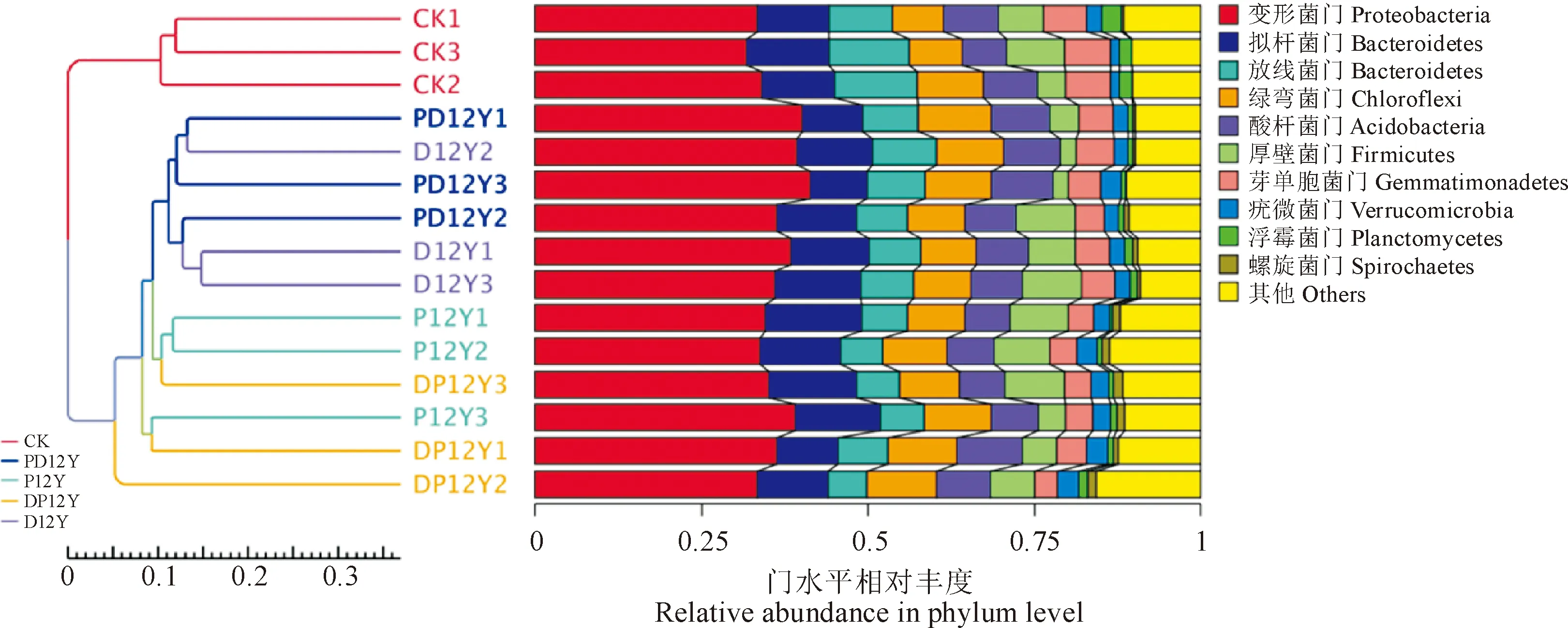
右边为样品的群落结构柱状图图4 基于Unweighted Unifrac距离的UPGMA聚类树The histogram of community structure is on the rightFig.4 UPGMA clustering tree based on Unweighted Unifrac distance
2.7 不同土地利用方式下细菌群落结构与土壤理化因子之间的相关性
通过R软件对环境因子和土壤细菌群落进行dbRDA分析。由图5可知,第一排序轴和第二排序轴分别解释了细菌群落变化的46.92%和30.97%,经过蒙特卡罗检验,土壤含水量(R2= 0.56,P= 0.01)和总氮(R2=0.51,P=0.01)是土壤微生物群落的主要影响因子。
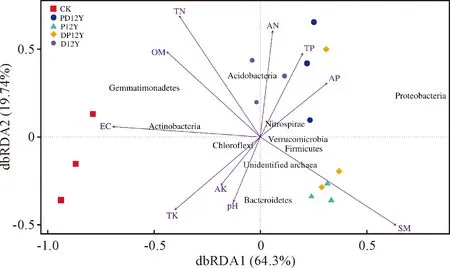
图5 不同土地利用方式盐渍化土壤性质与细菌群落的distance-based RDA分析Fig.5 Results from distance-based redundancy analysis (dbRDA) to explore the relationship between soil bacterial community and soil physicochemical characteristics
Spearman相关性分析表明(图6),排名前十的优势细菌门中,Gemmatimonadetes与总氮(P=0.01)显著正相关,与土壤含水量(P=0.02)显著负相关;Actinobacteria与总氮(P=0.00)、有机质(P=0.03)显著正相关,与含水量(P=0.00)显著负相关;Bacteroidetes和总氮(P=0.04)、碱解氮(P=0.03)显著负相关;Acidobacteria与速效钾(P=0.01)、电导率值(P=0.01)显著负相关;unidentified archaea与总氮(P=0.02)、碱解氮(P=0.00)和总磷(PS=0.04)显著负相关,与pH(P=0.03)、电导率(P=0.01)显著正相关;Firmicutes与总氮(P=0.03)、有机质(P=0.01)显著负相关,与土壤含水量(P=0.00)显著正相关;Verrucomicrobia与速效钾(P=0.02)显著负相关;Nitrospirae与电导率(P=0.00)显著负相关。其中,对优势菌门影响最大的土壤理化因子是总氮,其次是土壤含水量和电导率。

图6 不同土地利用方式盐渍化土壤优势菌群与土壤理化性质Spearman相关性分析Fig.6 Spearman correlation coefficient between soil dominant bacterial communities and soil physicochemical characteristics
3 讨 论
土壤细菌群落结构组成受种植作物和土壤环境的影响,不同农田生态系统中细菌群落的组成和丰度存在一定差异。本研究结果表明,变形菌门、芽单胞菌门、放线菌门、拟杆菌门、酸杆菌门、绿弯菌门等为水稻土壤中优势类群,这与陈力力等[6]、袁红朝等[16]对湖南地区酸性稻田土壤细菌群落的研究结果相似。
变形菌门被认为是世界上微生物最丰富的菌门,是多数土壤中的优势门类,与碳利用有关[17],在本研究中耕作使得盐渍化土壤中变形菌门相对丰度显著上升,改变了研究区域土壤微生物群落的基本组成。放线菌门相对丰度在对照组中占比较大(9.32%),这符合以往对盐碱地微生物的研究[18]。其中,土壤放线菌对一些有害真菌表现出明显的抑制效果,具有广谱抗生性及促进植株种子萌发、胚根生长的作用[19]。芽单胞菌门和放线菌门在本研究的4种耕作方式土壤中相对丰度均显著低于对照,芽单胞菌门和放线菌门在干旱土壤中的数量较多[20],是土壤属性变迁的生物学指标之一。其中芽单胞菌门与土壤含水量显著负相关,芽单胞菌门被认为既可在有氧条件又可在无氧条件下生长,具备广泛的适应性。放线菌是好氧菌群,能够降解复杂的木质素和纤维素[21],水稻连续种植12年(P12Y)和水旱轮作12年的水稻土壤(DP12Y)中放线菌门的相对丰度最低,而这两个处理中的土壤含水量最高,可见厌氧环境抑制了放线菌门的生长。Spearman相关性分析显示放线菌门相对丰度与土壤含水量呈显著负相关,与土壤总氮和有机质显著正相关。拟杆菌门与总氮和碱解氮显著负相关,因此土壤总氮和碱解氮对拟杆菌门菌群丰度起抑制作用,这与以往的研究一致[22]。酸杆菌门在酸性环境及寡营养条件适合生存[23],可作为土壤贫瘠的评价指标,在本研究中酸杆菌门与代表土壤盐分的电导率值显著负相关。研究表明,酸杆菌门参与单碳化合物的代谢和铁循环,在PD12Y和D12Y样品中酸杆菌门相对丰度显著升高,人为干扰引起酸杆菌门细菌的增加将有利于土壤中有机质的降解和铁的循环[24]。
水旱轮作模式下,土壤环境为厌氧及好氧条件交替,土壤中的碳/氮循环加速,是水旱轮作模式土壤中的微生物多样性高于旱地轮作模式的主要原因[25],这与本研究结果不一致,Shannon指数在连续种稻12年(P12Y)的土壤中最高,这可能是由于在水稻连作土壤中,水稻秸秆和残茬还田为土壤提供了更加丰富的有机质,有利于多种微生物的活动和繁殖,所以其土壤中微生物物种丰富,多样性高。4种土地利用方式下,土壤微生物群落功能多样性都比盐渍化荒地土壤显著升高,但4种耕作模式下土壤微生物的物种多样性指数、均匀度指数和优势度指数之间差异都不显著,这可能是由于土地利用方式、作物根系的活动、土壤状况、气候条件以及田间管理等多种影响因子综合作用的结果,任何单一因素的分析都很难做出较全面的解释。
大量无机肥料特别是氮肥的投入,通常会导致旱地土壤细菌多样性减少[26]。本研究结果显示Shannon指数在PD12Y最低,但与其他处理差异不显著。分析结果发现全氮和碱解氮均在种植玉米的旱地土壤中最高(PD12Y、D12Y),这表明种植玉米的土壤中氮肥的投入最高,反而抑制了土壤细菌多样性[22]。土壤电导率值可以间接地反映土壤含盐量,在本研究中,土壤盐分与多样性指数呈显著负相关,过高的土壤盐分对根际微生物有一定的抑制作用[27]。
土壤机械组成及分布特征是重要的物理属性,对土壤水热条件、肥力状况、养分循环等过程有重要影响[28]。砂粒含量在连续种稻(P12Y)和连续种植12年玉米地(D12Y)的土壤含量显著增加,说明长期连续种植单一作物会导致土壤沙化加快,引起土壤质量下降,排水和通气良好,但易干旱。本研究中,粉粒在水旱轮作的玉米地土壤(PD12Y)和连续旱作玉米地土壤(D12Y)增加,粉粒增加引起土壤表面紧实和结壳[29]。水旱轮作的玉米地土壤(PD12Y)和连续旱作玉米地土壤(D12Y)的粘粒含量相比于对照盐渍化荒地土壤都显著下降,因此盐渍化土壤种植玉米降低了土壤的粘粒含量,由此可见旱作增加了对土壤结构的扰动,更利于土壤形成良好的结构,减少土壤干旱时的板结状态,但是长期粘粒下降会导致土壤持水量下降。
土壤理化性状差异能显著影响微生物群落组成。本研究中,不同耕作方式下水稻和旱地土壤微生物群落结构与环境因子关系的dbRDA和相关性分析表明:土壤含水量、总氮、有机质、碱解氮、速效钾、电导率、pH值是影响土壤微生物群落结构的环境因子,其中土壤含水量、总氮和电导率是影响细菌群落的重要因子。因此,土壤含水量、营养因子和盐碱程度可能是限制宁夏银北盐渍化土壤微生物生长的因素,植物对土壤大部分的环境因子都有影响,对细菌群落结构的形成也具有重要作用,不同土地利用方式的细菌群落结构具有明显差异,这值得我们进一步研究。
4 结 论
通过对水旱轮作不同土地利用方式下土壤理化因子和土壤微生物群落多样性调查分析发现,研究区内土地利用方式深刻影响着土壤质量状况。
本研究中变形菌门、芽单胞菌门、放线菌门、拟杆菌门、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)等为优势细菌门。主成分分析和冗余度分析表明土壤理化性质和微生物群落能较好地响应盐渍化土壤不同利用方式的变化,说明通过区域土地资源的合理利用、土地利用结构的优化,可以改善盐渍化土壤质量因子状况,进而提高盐渍化区域土壤生产力和生态功能。本研究区域属于典型盐渍化土壤,总体土壤养分状况不佳,综合分析结果表明耕作使得土壤质量有所提升,其中提升效果依次为:水旱轮作(玉米)>连续种玉米>水旱轮作(水稻)>连续种稻。
