不同微生境下季风常绿阔叶林附生维管植物多样性特征
2022-01-26王艳红陈昭铨刘万德
李 敏,王艳红,唐 荣,陈昭铨,刘万德*
(1 中国林业科学研究院资源昆虫研究所,昆明 650224;2 南京林业大学,南京 210037;3 云南普洱森林生态系统国家定位观测研究站,昆明 650224)
附生植物指那些生长在其他植物体即宿主上而不吸取其营养,生活史的全部或者部分时期生长在空气中、不与地面接触的,利用雨露、水汽及有限腐殖质腐烂的枯枝残叶或动物排泄物等为生的一类自养植物[1]。附生植物是森林生态系统中的重要结构性成分,在森林生态系统物种多样性形成格局、养分和水分的循环过程中具有重要的生态意义[2]。由于附生植物没有直接与地面相连的根系,在获取水分和养分的过程中受环境条件的限制很大,绝大多数附生植物生长速率低,建群过程慢,对环境条件要求苛刻,对干扰也非常敏感[3]。近年来,在人为活动干扰下附生植物适生生境呈现出不断退化的趋势,进而导致附生植物物种数量减少甚至消失[4]。
尽管附生植物存在类别鉴定及定量研究上的难度,但近年来,有关附生植物的研究逐渐增加。附生植物的种类与分布受植被类型的影响[5]。研究显示,温带森林中苔藓类为主要附生类群,而在热带森林中,蕨类及兰科物种更多[6]。以往研究表明,附生植物的分布与水分有关。与乔木、灌木或草本植物相比,附生植物对水分供应的响应更快,在季节性较低的湿润森林中发现的附生植物多样性最高[7]。亚马逊盆地中,附生植物在潮湿的低地中也具有更高的物种多样性[8]。在有关附生植物分布研究中,Pittendrigh[9]首次观察到附生植物在树冠内的非随机分布,大树内附生植物的分布与枝条的大小、小气候有关[10]。在附生植物多样性及其影响因素研究中,Gonzlez等[11]通过定量评估巴拿马西部热带山地森林中附生植物生物量与物种多样性,结果表明,低温可能有利于苔藓植物的生长和降低分解速率。此外,光照强度也是影响附生植物多样性的重要因子,较高的光照强度会通过提高环境温度和水分蒸发速率,降低环境湿度,影响附生植物物种组成和丰富度[12]。同时,附生植物宿主特性(径级大小、高度、树皮特征等)的差异也会影响附生生境的稳定性,从而影响它们作为附生宿主的适宜性[13]。热带森林中的研究表明,单棵宿主上附生植物的丰富度随宿主径级的增加而增加[14]。在智利沿海雨林中的研究中,也得出了相似的结果,附生植物物种数随树木大小变化,较大径级的树木拥有更多的附生维管植物物种和更高的生物量,保留了大部分附生维管植物的多样性[15]。
热带与温带区附生植物多样性的研究显示,不同区域或不同森林类型下,附生植物多样性及其主要类群存在较大差异。在海南铜铁岭蕨类植物区系特点研究中表明,铜铁岭的蕨类植物区系的地理成分以热带和亚热带为主[16],然而,目前有关过渡区域森林内附生植物多样性研究还极少见报道。过渡区域兼具了两种不同气候带的气候和植被特征,对附生植物多样性必定会产生一定影响。作为热带和亚热带过渡区域的主要植被类型,季风常绿阔叶林是中国最复杂、生产力最高、生物多样性最丰富的地带性植被类型之一[17],对保护环境和维持全球碳平衡等都具有极重要的作用。近年来,有关季风常绿阔叶林的研究逐渐增多,但有关季风常绿阔叶林附生维管植物的研究鲜有报道。本研究通过对云南太阳河省级自然保护区内季风常绿阔叶林的附生维管植物的野外调查与数据分析,结合地形数据和其他环境数据,研究不同微生境下附生维管植物的物种组成和多样性,旨在回答:(1)不同生境下附生维管植物的物种组成和多样性特征;(2)影响附生维管植物多样性的主要因子。研究结果能够为全面研究季风常绿阔叶林生物多样性提供数据支持,为探索季风常绿阔叶林生物多样性保护措施提供科学依据。
1 研究地和研究方法
1.1 研究区概况
研究区域位于云南省中南部的普洱市,地理位置为22°02′~24°50′N,99°09′~102°19′E,海拔317~3 370 m。境内群山起伏,山地面积占98.3%,由于地处北回归线附近,地形十分复杂,受地形、海拔的影响,垂直气候特点明显,为亚热带季风气候,夏秋季多雨、冬春季干旱、年温差小、日温差大、干湿季明显。年均温15 ℃~20.3 ℃,年降雨量1 100~2 780 mm,年无霜期在315 d以上。普洱市森林覆盖率超过67%,光热条件良好,积温很高,适宜热带、亚热带生物生长,是全国生物多样性最丰富的的地区之一。区域内土壤以赤红壤为主。
季风常绿阔叶林是该地区主要的森林植被类型之一。由于人类活动的影响,季风常绿阔叶林常被砍伐作为薪炭、改造成茶园及农地等,只有在极偏远和保护区等地点还保存有部分原始林。季风常绿阔叶林中,乔木主要以短刺锥(Castanopsisechidnocarpa)、红锥(Castanopsishystrix)、西南木荷(Schimawallichii)等为优势树种[17];灌木则以珍珠伞(Ardisiamaculosa)、小叶干花豆(Fordiamicrophylla)等为主;草本多为毛果珍珠茅(Scleriaherbecarpa)及蕨类(Pteridophyta)等;同时群落中乔木的树干上附生有大量蕨类和兰科(Orchidaceae)植物。
1.2 野外调查
野外调查选择在普洱季风常绿阔叶林30 hm2动态监测样地内进行。该样地于2016年开始建设,2019年完成全部建设和本底调查工作,相关样地信息详见李帅锋等[17]和王艳红等[18]。根据王艳红等[18]对30 hm2动态监测样地微生境划分的结果(山脊、陡坡、缓坡、高谷和沟谷),在每种微生境内按照均匀分布的原则,选择30个20 m×20 m的样方进行附生维管植物调查,共调查150个20 m×20 m的样方。5种生境地形特征见表1。
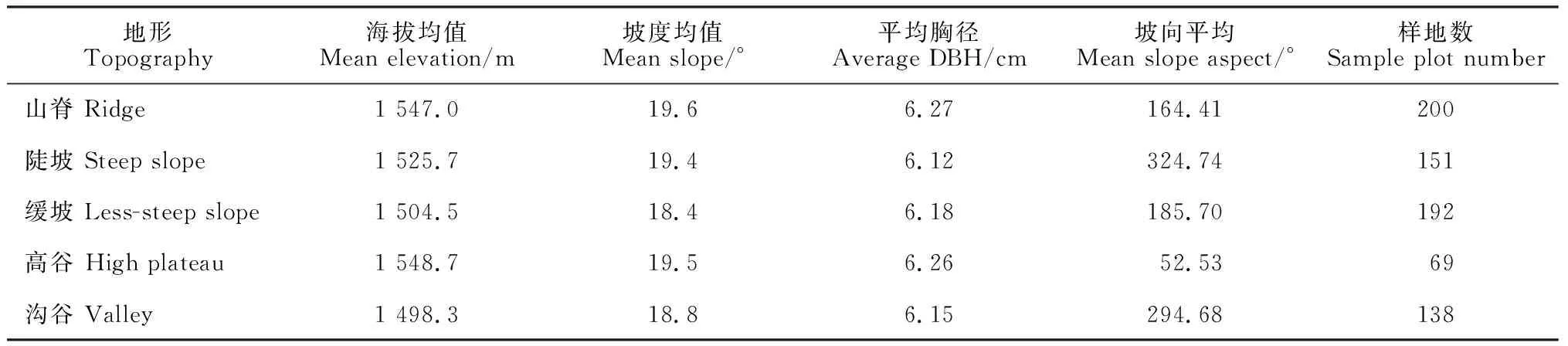
表1 研究区5类生境基本信息
针对选定的样方,调查样方内所有乔木树干及树冠上的附生维管植物。通过单绳攀爬技术,结合望远镜和激光测距仪[19],记录所有树木上出现的附生维管植物的物种名称、分布高度、株数,同时记录乔木树号信息。标本鉴定主要参考《中国植物志》,对未能鉴定的物种,请教有关专家协助鉴定。在调查过程中,部分附生植物的株数容易计数,如隐柄尖嘴蕨(Belvisiahenryi)、粗茎毛兰(Eriaamica)等,部分附生植物由于成团或成片附生在一起很难区分株数,如石豆兰属(Bulbophyllum)。因此,本研究利用已被广泛接受的Sanford界定方法:相同种的一群与另外一群具有明显的边界则区分成不同的株;不同种杂生在一起的,则分别计作一株[20]。附生植物宿主乔木信息根据记录的树号在30 hm2动态监测样地数据库中查询(通过查询可获得乔木的物种名称、胸径、树高及其位置信息)。
环境因子测定包括附生维管植物所在样方的海拔、坡度、坡向,样方中心点的光照强度、空气温度(T)。每个样方海拔为4个样方顶点海拔平均值;坡度为每个样方4个顶点任取3个顶点可以组成一个平面,可以获得4个不同平面,利用这4个平面与样方投影面夹角平均值可计算样方坡度[18];坡向则结合地理信息系统与野外实测对照获取;样方中心点光照强度通过LI-1500辐射照度测量仪来测定,各样方的空气温度利用温湿度测量仪,通过测定样方对角线三点的数据以均值来表示各样方的空气温度。
1.3 数据分析
附生维管植物的物种丰富度用其物种数表示,个体多度用其株数表示。为定量描述本研究中附生维管植物的多样性特征,本研究采用Shannon-Wiener(H)分别计算不同微生境下附生维管植物的多样性指数,计算公式如下:
式中:S为样方中物种数目;Pi为样方中属于第i种的个体的比例,如样方总个体数为N,第i种个体数为ni,则Pi=ni/N。
根据宿主植物胸径大小,将其划分成6个阶段:DBH<10 cm,10 cm≤DBH<20 cm,20 cm≤DBH<30 cm,30 cm≤DBH<40 cm,40 cm≤DBH<60 cm及60 cm≤DBH;根据附生维管植物分布高度(H)大小划分成6个阶段:H<2 m,2 m≤H<4 m,4 m≤H<6 m,6 m≤H<8 m,8 m≤H<10 m及10 m≤H)。计算不同胸径大小及不同高度附生植物多样性。
根据野外调查结果,整理各样方内各物种株数,利用 R 3.6.1 中 vegan 包的 specaccum 函数绘制物种-面积累积曲线。对调查区域内物种,按照多度由高到低的顺序排列,绘制物种-多度曲线图。以每个样方作为统计单位,根据各样方内附生维管植物物种丰富度、多度及多样性指数数据,比较不同生境附生维管植物的多样性差异,以每类生境作为统计单位,比较不同宿主径级、不同分布高度下附生维管植物物种多样性,大小比较采用方差分析进行,利用aov函数建立单因素方差分析模型,利用TukeyHSD函数进行均值的多重比较分析,通过字母标记法表示显著性差异,误差线代表平行测定值的标准误。根据150个样方的环境数据及各样方的物种丰富度及多度数据,使用Pearson相关分析方法,通过cor函数计算相关系数,探索环境因子对附生维管植物丰富度及多度的影响,利用corrplot函数进行可视化处理,根据相关性分析结果,建立回归方程,进一步研究附生维管植物丰富度、多度与环境因子之间的关系及变化规律。所有数据分析均在R 3.6.1(R Development Core Team, 2019)中完成,显著性水平为P<0.05。
2 结果与分析
2.1 季风常绿阔叶林附生维管植物物种组成
季风常绿阔叶林内共有附生维管植物16科45属97种12 302株。主要类群为兰科植物和蕨类植物。其中,兰科物种最丰富,包含45属54种,占总物种数的56%,但个体多度仅占19.2%;蕨类包含12科22属39种,占总物种数的40%,但个体多度高达80.7%,其中水龙骨科(8属17种)占附生蕨类植物物种数的44%。除以上两大类外,还记录到其他维管植物3科3属4种,主要是苦苣苔科(Gesneriaceae)和天南星科(Araceae)物种。
从种-面积曲线来看(图1),随着调查面积的增加,季风常绿阔叶林附生维管植物的物种丰富度已基本接近渐近线,说明本次调查样本量对于季风常绿阔叶林的附生维管物种丰富度已经足够,季风常绿阔叶林内绝大部分附生维管植物物种都在调查记录的范围之内。季风常绿阔叶林附生维管植物物种-多度曲线(图2)表现出急剧变化,表明优势物种明显,仅有少数几个物种占有明显优势,低密度种占较大比例。
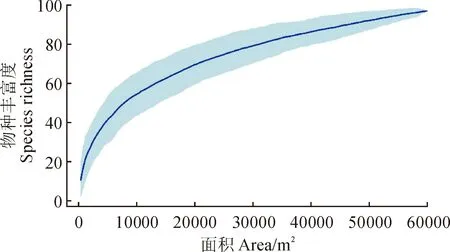
图1 研究区附生维管植物的种-面积曲线Fig.1 Species-area curve of epiphytic vascular plants in the study area

图2 研究区附生维管植物的物种-多度曲线Fig.2 Species-abundance curve of epiphytic vascular plants in the study area
2.2 不同微生境下附生维管植物多样性
在5种微生境下,附生维管植物的物种数分别为陡坡60种,缓坡56种,高谷47种,沟谷61种,山脊53种。根据方差分析结果,山脊的物种丰富度及个体多度均值最大,而其余4种微生境间附生维管植物物种丰富度和个体多度无显著性差异(图3)。山脊的Shannon-Wiener指数为2.53,同样高于其他4种生境(P<0.05)。表明在山脊中,附生维管植物物种多样性较高,这可能与不同生境宿主植物密度及地形条件的差异有关。
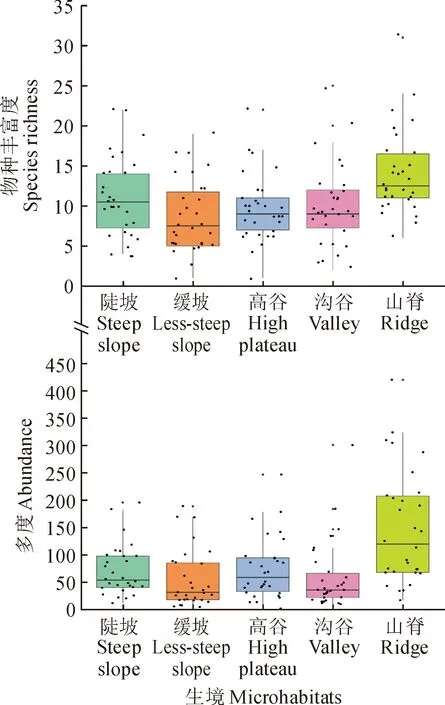
图3 不同生境附生维管植物分布Fig.3 Distribution of epiphytic vascular plants on different microhabitats
2.3 不同宿主特征下附生维管植物多样性
附生维管植物多样性和宿主胸径密切相关。不同径级下,附生维管植物的丰富度呈现出不同的变化趋势(图4)。整体上看,研究区域中宿主树木胸径在20 cm以下,附生维管植物的丰富度较高,物种组成相对丰富,随着宿主胸径的增加,附生维管植物的物种累积数在逐渐减少。不同胸径范围内,DBH<10 cm与10 cm≤DBH<20 cm间附生维管植物多样性无显著性差异(P>0.05),但10 cm≤DBH<20 cm附生维管植物多样性显著高于其余胸径范围(P<0.05)。

图4 不同径级宿主上附生维管植物分布Fig.4 Distribution of epiphytic vascular plants on different DBH size classes of host trees
普洱季风常绿阔叶林附生维管植物在宿主植物的树干和树冠上均有分布,但多数物种主要分布在宿主树干中下部,随着高度的增加,附生维管植物多样性逐渐降低(图5)。在林冠下部的树干上,附生维管植物种类丰富,主要有隐柄尖嘴蕨及众多兰科植物,随着高度的增加,附生植物种类逐渐降低。

图5 附生维管植物的空间分布Fig.5 Spatial distribution of epiphytic vascular plants
2.4 附生维管植物多样性的环境解释
2.4.1 附生维管植物物种丰富度、个体多度与环境变量的相关性分析根据相关性分析结果(图6),在所选光照强度、温度、海拔、坡度和坡向5个环境变量中,海拔、光照强度与附生维管植物的物种丰富度有显著相关性,其中,海拔与物种丰富度为正相关(r=0.37),光照强度与物种丰富度为负相关(r=-0.56)。海拔、温度、光照强度与附生维管植物的个体多度有显著相关性,其中,海拔与多度为正相关(r=0.36),温度与多度为正相关(r=0.26),光照强度与多度为负相关(r=-0.33)。
不同的环境变量之间也存在一定的相关性。光照强度与海拔、坡向与温度之间存在相关性,海拔与温度之间存在极显著的相关性。
2.4.2 附生维管植物物种丰富度及个体多度与环境变量的回归分析根据相关性分析结果,对附生维管植物物种丰富度、多度分别与环境因子进行回归分析(图7),结果表明,物种丰富度与海拔高度(x)存在线性回归关系,其回归方程为y=-85+0.065x(R=0.37)。回归方程中一次项系数也表现出显著的正相关性,进一步说明物种丰富度与海拔高度之间为线性正相关。物种丰富度与光照强度也存在线性回归关系,用回归方程表示为:y=14-0.0079x(R=-0.56),回归方程中一次项系数也表现出显著的负相关性,进一步说明物种丰富度与光照强度之间为线性负相关。个体多度与光照强度、海拔高度、温度存在线性回归关系,回归方程分别为y=110-0.068x(R=-0.33),y=-1300+0.91x(R=0.36),y=-260+14x(R=0.26)。

图7 物种丰富度及个体多度与环境因子间的关系Fig.7 Relationships between species richness and abundance and environment variables
2.4.3 附生维管植物物种丰富度及个体多度与环境变量的逐步回归分析物种丰富度与环境变量之间的逐步回归分析显示,模型中存在两个环境变量,海拔高度(x1)、光照强度(x2),物种丰富度与环境变量之间的关系可以用回归方程表示为:y=0.05x1-0.007x2-59.842(R2=0.391,P=0.000)。
个体多度与环境变量之间的逐步回归分析显示,模型中存在两个环境变量,海拔高度(x1)、光照强度(x2),附生维管植物与环境变量之间的关系可以用回归方程表示为y=0.793x1-0.057x2-1 056.27(R2=0.204,P=0.000)。
3 讨 论
附生植物的物种组成及丰富度受森林类型及森林结构因素影响[12, 21]。本研究中,共调查到附生维管植物97种,单位面积(每hm2)物种数为16种,低于西双版纳(116种)[22]、云南省哀牢山湿性常绿阔叶林(30种)[23]和海南霸王岭(17种)[24]。不同地区附生植物的物种组成及数量存在较大差异,这主要与研究地的森林类型有关。在巴西阿克里州的热带森林中,对附生维管植物的区系组成进行研究,共记录附生植物2 446种,天南星科是最丰富的科(76种,占总数的23%),其次是兰科(55种,占总数的17%),水龙骨科(34种,占11%)[25]。温带森林中,蕨类植物(特别是膜蕨科植物)在附生植物多样性中占主导地位[15]。在本研究中,附生维管植物主要以兰科植物为主,其中石斛属和石豆兰属附生种数最多;其次是蕨类,其中水龙骨科物种丰富度及个体多度均占附生蕨类半数以上。这种科属组成和物种丰富度在一定程度上体现了兰科和蕨类植物对附生生境的适应性进化。与热带森林相比,季风常绿阔叶林结构相对简单,年降水量较少,其空气湿度、光照强度、郁闭度均与热带森林有显著差异;与中山湿性常绿阔叶林(哀牢山)相比,季风常绿阔叶林空气湿度较低,从而导致了季风常绿阔叶林中附生植物的多样性相对较低。附生维管植物在季风常绿阔叶林中的分布也呈现出一定的规律性,54.6%的附生维管植物分布在山脊和高谷中,山脊也具有最高的多样性指数,附生维管植物分布广泛,而沟谷中附生维管植物丰富度较高,但株数少,一方面可能是由于地形条件的差异,另一方面,可能也与光照强度、温度等环境因子有关,研究区域季风常绿阔叶林整体郁闭度较高,光照强度较弱,附生植物虽多生长在潮湿背阴的地方,但沟谷海拔低,光照强度过小,部分区域可能达不到附生维管植物的光补偿点,并不利于附生维管植物生长和分布,导致其附生植物数量偏低。
在以往对附生维管植物的森林调查研究中发现,生境条件的差异对附生维管植物的物种组成及多样性有较大影响,在局部尺度上,小气候的分化被认为是维管附生植物群落结构的主要机制之一[26-28]。附生维管植物分布在树干或树冠上,直接暴露在不断变化的气候环境下,微小的小气候差异也会影响附生维管植物群落的区系组成[29]。温度影响花芽分化过程,对附生兰科植物文心兰(Oncidiumhybridum)的研究表明,27 ℃时文心兰的生物量最高,过高或过低的温度都不利于其生长[30]。光照强度通过影响气孔导度影响附生植物对光适应的生态幅,不同附生植物对光的适应性不同,阴石蕨对高光表现出光抑制,表现出阴生植物的特性[31]。此外,地形条件如海拔、地形凹凸度等也是影响附生植物的重要因素,中海拔地区附生植物具有更高的生物多样性,呈现驼峰形状的物种丰富度模式,高海拔地区丰富度下降更明显[32]。在研究沿垂直树干高度和海拔高度两个环境梯度的乔木灌木地衣的生物量和分布特征中发现,这两个梯度都影响附生植物的生物量和分布,附生植物生物量主要分布在树木中等高度,与海拔高度相关的气候梯度也会影响附生植物群落结构[33]。本研究中,光照强度、海拔、温度、坡度、坡向是影响植物群落的主要环境因子,5种生境光照强度、温度、海拔等生境因子均存在较大差异,附生维管植物也呈现出不同的分布规律,研究结果中,附生维管植物的丰富度和多度均整体随光照强度的增强而呈下降趋势,与前人的研究结果基本一致,两者均随海拔及温度升高而逐渐增大,这种关系主要是由于本研究中海拔梯度、温度范围变化较小(海拔范围在1 300~1 500 m之间,温度范围在21~28 ℃),因此仅体现出研究区域局部温度、海拔梯度下附生维管植物丰富度变化规律。相关性分析结果中坡度、坡向与附生维管植物的丰富度、多度均没有显著相关性,可能是由于不同坡度、坡向下的环境异质性不是很大,因此对附生维管植物多样性的影响相对较小。
除生境因子外,生物因子(宿主)也是影响附生维管植物多样性的重要因素。宿主物种为附生植物的生长提供了不同的基质、条件和微气候,因此宿主特性是影响附生植物分布的重要因素[34-37]。附生植物的分布与宿主树种密切相关,在系统发育上相似的树木或共有形态或功能性状的树木可能拥有相似的附生植物物种[38]。在本研究中,附生维管植物更多地分布在中小径级的树木上,且分布高度较低。以往研究显示,因宿主径级影响着微生境异质性和附生维管植物的定殖面积,大径级树木能为附生植物提供更丰富的腐殖质和定殖空间,故附生维管植物更多地分布于径级较大的宿主植物上[13]。此外,附生植物在森林中的分布具有明显的高度偏好性,天南星科和几种蕨类植物多分布在下层,兰科大部分分布在宿主较高的位置[11]。本研究的结果主要与季风常绿阔叶林中树种特性及生长状况有关,在研究区中,宿主胸径整体较小,大径级树木的个体数较少,例如,隐距越桔(Vacciniumexaristatum)胸径大部分在15 cm以内,而其又是本研究中附生维管植物的主要宿主之一。因此不同径级宿主数量差异较大,大径级宿主比例极低,从而导致了附生维管植物更多地分布于中小径级树木上。同时,因季风常绿阔叶林林冠高度在25 m左右,在宿主较高位置,光照强度增强,温度升高,湿度降低,不利于附生维管植物生长,因此大部分附生维管植物分布在宿主树干中下部位。通过研究影响附生维管植物物种组成和多样性的主要生境因子和生物因子,反映出多样生境条件、宿主条件下的物种组成和分布规律,有利于今后评估季风常绿阔叶林森林受干扰程度和生境健康与否,也为附生维管植物的多样性保护提供依据。
