2015—2020年乳山近岸海域春、夏季大型底栖动物群落结构及变化
2022-01-25纪莹璐张海舟王俊健孙静宜
纪莹璐,宿 凯,张海舟,王俊健,孙静宜,张 亮*
(1.国家海洋局北海预报中心,山东 青岛 266061; 2.山东省海洋生态环境与防灾减灾重点实验室,山东 青岛 266061)
乳山市位于山东半岛东南部,介于北纬36°41′—37°08′,东经121°11′—121°51′之间,东西最大横距60 km,海岸线西起乳山口,东至浪暖口,近岸海域底质基本为黏土质粉砂,水体呈典型温带水域海水特征,季节变化明显,是我国重要的经济贝类牡蛎的养殖地[1-2].乳山牡蛎养殖区主要集中在沿岸浅海开阔水域内,而长期规模化的高密度贝类养殖,可能通过影响海洋生态系统的能量代谢和物质循环过程,导致局部海域营养结构失衡[3-4].近年来,针对乳山附近海域的大型底栖动物研究较少[5-6],缺少最新的基线数据,且缺乏年际调查资料,该研究作为乳山近海连续调查的阶段性结果,以2015—2020年乳山近岸海域春、夏季大型底栖动物为对象,分析其群落结构的年度变化及养殖活动对底栖群落的影响,为该海域提供了最新的大型底栖动物基线数据,以期为大型底栖动物群落的演变机制和近海生态系统的深入研究打下基础,同时为乳山近海生物资源的合理开发利用和环境保护提供科学依据.
1 材料与方法
1.1 野外采样与室内分析
从乳山近岸海域东西向空间尺度上看,若设置较多站位,站位间距离相对较近,群落结构相似度高,代表性不强,也增加了长期调查的成本.笔者根据乳山近岸海流流向基本呈现东南—西北的特点,由乳山口向西设置RS3站位,乳山湾口设置RS1站位,银滩砂质岸线浪暖口周边设置RS2站位,如图1所示,3个站位分别代表了乳山近海3个不同特点的底栖生境.大型底栖动物繁殖季节大多在春季,每年春、夏季开展生态调查以获得较为完整的种群组成和生物资源数据.每站位以0.05 m2抓斗采泥器采集4次,合为1个样品,保证采样的质量,采得沉积物用孔径为0.5 mm的网筛冲洗分选,留在网筛上的样品全部收集装瓶,并用体积分数为10%的福尔马林溶液固定.同时采集底层海水装瓶24 h内实验室测定,并采集一定量的表层沉积物放入-20 ℃低温冰柜冷冻保存.环境因子测定沉积物有机碳、表层叶绿素a(0~2 cm)、粒度,底层水pH、盐度、溶解氧、化学需氧量、磷酸盐、无机氮.样品带回实验室在体视显微镜下进行分类、鉴定、计数,使用感量为0.000 1 g的电子天平进行称量.以上所有样品的采集、处理、保存、计数、称量均参照《海洋监测规范》[7]和《海洋调查规范》[8]进行.

图1 乳山近岸海域大型底栖动物调查采样点
1.2 数据处理
采用相对重要性指数(IRI)来确定大型底栖动物优势种,计算公式为
IRI =(W+A)×F,
(1)
其中:W为某一种的生物量占大型底栖动物总生物量的百分比,A为该种的丰度占大型底栖动物总丰度的百分比,F为该种出现的频率[9],IRI>1 000的种定为优势种,100≤IRI≤1 000的种为重要种[10].多样性分析采用Margalef物种丰富度指数(d)、Shannon-Wiener多样性指数(H′)以及Pielou物种均匀度指数(J)[11-13].
d=(S-1)/log2N,
(2)
H′=-ΣPilog2(Pi),
(3)
J=H′/log2S,
(4)
其中:S为样品的总种数,N为样品的总个体数,Pi为样品中第i种的个体数与总个体数的比值.使用PRIMER 6.0进行大型底栖动物群落结构和丰度/生物量比较曲线分析,各季度大型底栖动物丰度数据log(x+1)转换后(x为丰度),经Bray-Curtis相似性等级聚类、非度量多维标度及相似性分析,比较群组间的相似性并筛选表征群落特征的物种[14].运用SPSS 19.0进行Kruskal-Wallis非参数检验.
2 结果与分析
2.1 环境因子
2015—2020年各站位环境概况见表1所示.调查站位均设置在水深较浅(2~15 m)的海域,由底质类型的变化来看,2015年乳山近岸海域底质类型以黏土质粉砂为主,到2020年逐渐转为粉砂质,底质中黏土含量由25.86%下降至6.08%,海域沉积物粒径增大.叶绿素a含量一般用于评价沉积物中底栖动物的食物来源和栖息环境的质量,调查海域叶绿素a 含量为1.03~17.2 mg·g-1,整体呈现逐年上升的趋势.调查海域属于近海,受地表径流和浅海养殖等多方面因素影响,黏土环境因子的变化趋势较为复杂.根据《海水水质标准》[15]和《海洋沉积物质量》[16]对环境数据进行评价,水质pH、溶解氧、COD和磷酸盐均符合海洋一类水质标准,无机氮除2016和2019年符合海洋二类水质标准外,其余年份均有站位达到三类或四类水质;沉积物有机碳符合沉积物第一类质量要求.综上所述,乳山近岸海域海水中无机氮常年处于高值,可能存在污染风险.

表1 各站位主要环境因子
2.2 底栖动物种类组成和优势种
2015—2020年乳山近岸海域春、夏季共发现115种大型底栖动物,包括50种环节动物,36种节肢动物,20种软体动物,5种脊索动物,纽形动物、棘皮动物各2种,其中环节动物、软体动物和节肢动物占总种数的92.2%.从年际变化来看(图2),2015—2020年分别鉴定到大型底栖动物42,33,28,25,32,33种,种类数以2015年最高,2018年最低,2019—2020年种类数变化不大.种类组成上,2015,2016,2019,2020年环节动物多毛类均为最主要类群,节肢动物、软体动物次之,而2017和2018年节肢动物成为主要类群,其次为环节动物和软体动物,二者比例接近.综合来看,2017和2018年大型底栖动物总种数下降,虽然各类群种类组成未发生变化但比例与前后几次调查均存在差异.

图2 2015—2020年乳山近岸海域春夏季种类组成
乳山近岸海域春、夏季出现的大型底栖动物优势种共20种(表2),包括5种多毛类,6种软体动物,6种节肢动物,1种棘皮动物和2种脊索动物,但优势种的年际变化明显,多数优势种仅在某一年度出现.中蚓虫(Mediomastuscaliforniensis)、鹰爪虾(Trachypenaeuscurvirostris)和红鳗虾虎鱼(Taenioidesrubicundus)在3个年份为优势种,不倒翁虫(Sternaspisscutata)、寡鳃齿吻沙蚕(Nephtysoligobranchia)、日本毛虾(Acetesjaponicas)和中国毛虾(Aceteschinensis)在2个年份为优势种.

表2 各年度大型底栖动物主要优势种及其优势度
2.3 底栖动物丰度及生物量变化
乳山近岸海域春、夏季的大型底栖动物平均丰度为376 ind·m-2,平均生物量为72.11 g·m-2(表3).多毛类是该海域大型底栖动物丰度的主要贡献者,占据了总平均丰度的59.48%,甲壳类次之,占据了总平均丰度的25.26%,其丰度的年际变化较为明显,最高可达年丰度贡献量的51.03%,最低则为4.60%.不同类群生物量的贡献则与丰度有所差异,平均生物量以软体动物最高(占总生物量的43.64%),甲壳类(38.73%)的贡献次之.

表3 大型底栖动物平均丰度及平均生物量组成
从图3可以看出,大型底栖动物丰度在2015—2020年出现3个波峰,2015,2017,2019年丰度较高,生物量从2015年的34.86 g·m-2先升至2017年的220.26 g·m-2,而后降至2019年的27.47 g·m-2,其中2017年生物量较为异常,因为该年出现了大个体的脉红螺(Rapanavenosa)和个数较多的鹰爪虾,Kruskal-Wallis检验表明不同年度密度(p=0.062)和生物量(p=0.744)均不存在显著差异.2015,2016,2019,2020年丰度组成均以多毛类占较大优势,2017,2018年多毛类所占比例下降,甲壳类比例上升;2015年大型底栖动物生物量以棘皮动物占据优势地位,之后几个年度生物量组成发生变化,基本呈现软体动物和甲壳类占优势的特点.
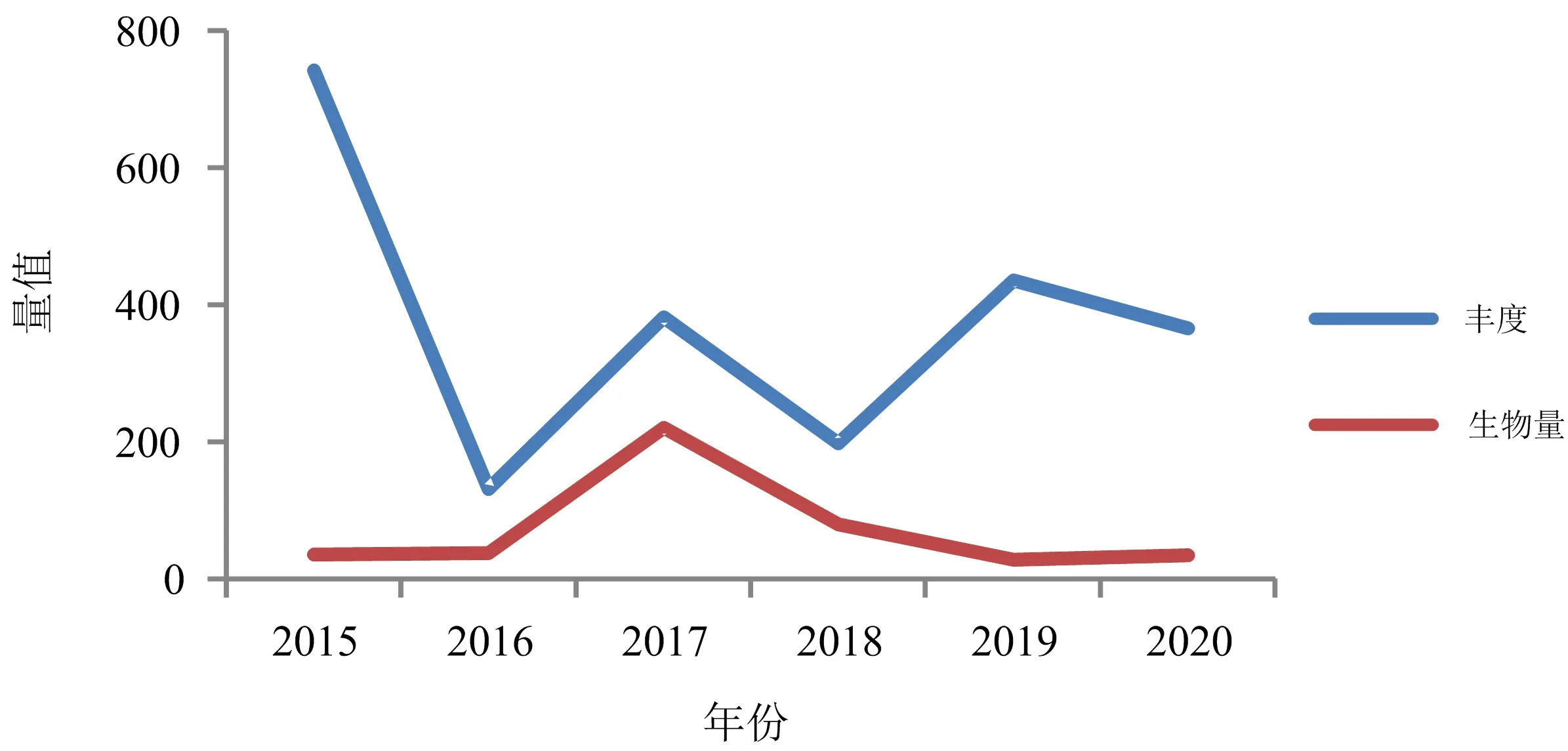
图3 2015—2020年乳山近岸海域春、夏季大型底栖动物丰度和生物量变化
2.4 生物多样性指数
2015—2020年乳山近海春、夏季大型底栖动物多样性指数见表4所列,其中Shannon-Wiener多样性指数(H′)平均为3.04,最小值为2.03,最大值为3.86,分别出现在2017年和2015年;Pielou物种均匀度指数(J)平均为0.860,最小值为0.718,最大值为0.926,分别出现在2017年和2020年;Margalef物种丰富度指数(d)平均为2.44,最小值为1.37,最大值为3.39,分别出现在2017年和2015年.各站位间多样性指数差异较小,不同年份间差别较大,与2015年相比,2016—2018年多样性指数下降,年平均值均小于3,2019年之后多样性指数和丰富度指数逐渐上升并趋于平稳.

表4 不同站位大型底栖动物多样性指数
2.5 底栖动物群落结构组成和相似性分析
对2015—2020年乳山近岸海域春、夏季大型底栖动物种类组成进行聚类和SIMPER分析,结果如图4所示,在15%的相似性上,可将6个年份分为2组.2017年和2018年为群落Ⅰ,该组平均相似性为43.23%,特征种为日本毛虾(贡献率33.33%)和中国毛虾(贡献率28.00%);2015,2016,2019,2020年为群落Ⅱ,该组平均相似性仅为19.87%,特征种为中蚓虫(贡献率16.97%)、寡鳃齿吻沙蚕(贡献率7.55%)、不倒翁虫(贡献率7.15%)、多丝独毛虫(Tharyxmultifilis)(贡献率6.54%)、纽虫一种1(Nemertinea sp.1)(贡献率5.41%)、海稚虫科一种(Spionidae)(贡献率5.38%)和双壳类幼体(Bivalvia)(贡献率5.08%).

图4 2015—2020年乳山近岸海域春、夏季大型底栖动物种类组成聚类分析
3 讨论与结论
3.1 大型底栖动物优势种的历史变化
2006—2007年乳山湾及邻近海域大型底栖动物优势种包括中蚓虫、不倒翁虫、寡鳃齿吻沙蚕、滑理蛤(Theoralubrica)和轮双眼钩虾(Ampeliscacyclops)等,且以多毛类为主要优势类群[17].2009年8月王宗兴等[6]对乳山湾外海的大型底栖动物按照其出现频率和相对丰度进行筛选,得出用于CCA排序的32个主要物种,相较之前调查除不倒翁虫仍占优势以外,其他优势种已被新物种所取代.2011年,赵宁等[18]对乳山湾内外大型底栖动物生态调查结果显示,中蚓虫、足刺拟单指虫(Cossurellaaciculata)、矮小稚齿虫(Prionospiopygmaea)、多丝独毛虫(Tharyxmultifilis)、不倒翁虫和寡鳃齿吻沙蚕为优势物种.笔者的研究显示,乳山近岸海域大型底栖动物种类丰富,3个站位6年间出现IRI>1 000的优势种20种,且连续两年以上出现的优势种种数仅占该海域优势种总种数的35%,群落聚类分析显示不同调查频次间大型底栖动物群落结构相似度较小,在一定程度上也反映出该海域群落结构的变化较大.2015年之前的优势种中大部分为多毛类,其他类群如软体动物和节肢动物只有个别种为优势种,此后特别是2017和2018年,软体动物和节肢动物取代多毛类成为群落结构中的主要成分,其余年间多毛类和其他类群优势种占比基本持平.结合以往研究结果分析,除2009年以外,乳山近岸海域大型底栖动物中蚓虫、不倒翁虫和寡鳃齿吻沙蚕在多个年份均为优势种,这与王洪法等[19]对2000—2009年胶州湾大型底栖动物优势种的分析结果十分相似,但从优势类群上看乳山海域已由多毛类占绝对优势逐渐向多毛类、软体动物和节肢动物占比均衡转变,胶州湾优势类群依旧为多毛类,软体动物中菲律宾蛤仔在养殖区附近海域作为优势种出现,而节肢动物和棘皮动物未出现达到优势种判别值的物种.
自20世纪末浅海筏式养殖技术推广后,乳山牡蛎的养殖区就一直集中于沿海一线的开阔水域内,随着“乳山牡蛎”获国家地理商标认证,牡蛎的养殖规模呈现逐年扩大的趋势[20].根据2006—2016年《山东省海洋环境质量公报》[21-31],2006年乳山湾内及湾外以南海域海水水质质量为轻度污染,湾外以北大部为较清洁海域,伴有轻度污染,主要污染因子为无机氮和活性磷酸盐,近岸海域沉积物总体状况一般,彭松耀等[5]同年研究也显示乳山近海底栖群落健康状况由虚弱趋于不稳定,大部分站位底栖群落存在轻度扰动,且多毛类成为绝对优势类群;2010年以后乳山近岸海域水质和沉积物质量有所改善,海水水质基本符合第一、二类海水水质标准,超标物质为活性磷酸盐,海域富营养化程度逐年减轻,沉积物符合第一类海洋沉积物质量标准.一般来说,多毛类优势种增加是海域出现有机污染的重要指示,虽然笔者的该次调查中海水无机氮依然有持续高值的情况,但整体来看,近10年乳山海洋环境质量有所提升,多毛类在优势种类群中不再占据绝对优势.根据大型底栖动物不同食性划分,乳山近岸海域重要的优势种中蚓虫和不倒翁虫为底泥食性,寡鳃齿吻沙蚕为肉食性,而底泥食性物种多分布在底质较细且有机质含量丰富的海域,具有适应性强和耐污染的特点,是转化外源性化学物质的重要物种[32-33],已有研究表明寡鳃齿吻沙蚕的数量与有机质含量亦存在正相关关系,肉食者作为高营养级其丰度较大的区域碳通量通常较高[34-35],乳山近岸海域沉积物有机碳符合沉积物第一类质量要求,但表层叶绿素a有逐年上升的趋势,上述3个物种的优势地位也可反映出乳山近岸海域沉积物有机质含量高的特点,这可能与浅海牡蛎养殖产生的残饵及代谢产物累积有关.
3.2 大型底栖动物群落和多样性的变化
聚类分析结果表明,2015—2020年间乳山近岸海域春、夏季大型底栖动物群落相似性程度低,2017,2018年与其他年份的群落结构有明显差异,表现为多毛类种数下降,软体动物尤其是节肢动物的种数、丰度和生物量均上升,且整个海域的总种数低于往年,群落结构的变化可能与位于乳山南黄岛的路岛海岸工程两年间的建设开发有关.一般来说海岸开发和海洋工程建设与运营使底栖动物的栖息地范围变小,航道疏浚和土石方倾倒造成严重的底质搅动和翻耕,导致沉积环境不稳定,大型底栖动物的丰度、生物量和多样性降低,海域生态功能逐渐衰退[36],2017年后各站位的沉积物粒径大小发生明显变化,表现为粒径较小的黏土含量下降,砂和粉砂含量上升,底质类型由黏土质粉砂为主变为砂质粉砂或粉砂.与乳山海域历史资料相比(表5),笔者的研究所获取的物种数明显少于2006—2007年的103种和2011年的161种,但种类组成除个别年份外多毛类一直为最优势类群,多毛类、软体动物和节肢动物构成的乳山近岸海域大型底栖动物主要类群未发生变化;大型底栖动物Shannon-Wiener多样性指数(H′)值由2006年的2.44升至2020年的3.78,丰度在2011年达到峰值后下降与之前数据基本持平,生物量则呈现先上升后下降的趋势.从群落多样性的产生与维护看,近10年来乳山近岸海域大型底栖动物多样性增加,自我调节能力提升,根据蔡立哲等[37]2002年提出的海洋底栖动物多样性指数污染程度评价标准,乳山近岸海域从轻度污染(H′在2~3之间)转为清洁(H′大于3).与此同时,调查海域物种数目的减少和群落结构相似性程度的降低也表明该海域底栖生态环境已发生了较大的改变,已有研究显示[6]中蚓虫和寡鳃齿吻沙蚕对高总氮生境的适应性强,而不倒翁虫则与总磷呈较强的负相关关系,分布的环境表现出水深、总磷低的特点,乳山近岸海域大型底栖动物种类组成中中蚓虫、不倒翁虫和寡鳃齿吻沙蚕等优物种持续占据优势,而其他物种的年际变化明显,说明乳山近岸海域常年处于高总氮低总磷的环境,这与该次调查的环境因子结果一致.结合海域大型底栖动物丰度基本维持原有水平但生物量低于以往调查的现状,说明底栖动物个体存在一定程度的小型化趋势,虽然多样性指数显示乳山近岸现为清洁海域,优势种各类群占比中多毛类、软体动物和节肢动物的占比趋向均衡,但其仍存在潜在的有机质污染风险,为防海域可能出现的生态环境恶化和物种资源持续衰退,促进乳山近海海洋生态系统的健康持续发展,需开展长期持续的调查监测,并进一步提高对海岸开发和海洋工程的管理力度,制定有效的海洋生态综合治理措施.

表5 该次调查结果与历史资料的对比
