粳稻穗部不同部位籽粒产量相关性状差异及其与内源激素的相关性
2022-01-18郑小龙1周菁清滕颖章林平邵雅芳胡培松1魏祥进
郑小龙1 周菁清 滕颖 章林平 邵雅芳 胡培松1 魏祥进*
粳稻穗部不同部位籽粒产量相关性状差异及其与内源激素的相关性
郑小龙1,2周菁清3滕颖4章林平2邵雅芳2胡培松1, 2魏祥进2, *
(1江西农业大学 农学院, 南昌 330045;2中国水稻研究所, 杭州 314001;3浙江环境监测工程有限公司, 杭州 310015;4江苏省常州市金坛区种植业技术推广中心, 江苏 常州 213200;*通信联系人, E-mail: weixiangjin@caas.cn)
【】根据灌浆期水稻内源激素水平解析稻穗不同部位籽粒产量性状的差异。以相同环境下种植的嘉58等6个浙江省常规粳稻品种为实验材料,将稻穗籽粒按一次枝梗在主穗上的节点划分为稻穗上部、中部和下部籽粒,测定了不同部位籽粒水杨酸(SA)、脱落酸(ABA)、吲哚乙酸(IAA)和生物量的动态变化及收获后籽粒千粒重、整精米率、长宽比、胶稠度和碱消值等相关性状。籽粒在灌浆期内0~35 d处于持续增重状态,且表现为上部>中部>下部,花后35 d各品种不同部位籽粒生物量的变异系数为1.66%~10.26%,35 d以后不同部位籽粒生物量的差距逐渐缩小但仍有差异,籽粒成熟后千粒重仍表现为上部籽粒(25.08~33.88 g)>中部(24.04~32.49 g)>下部(23.46~31.78 g);不同部位间籽粒整精米率与长宽比略有差异,且千粒重与长宽比呈极显著正相关(= 0.89,< 0.01),与整精米率呈极显著负相关(= −0.68,< 0.01);在胶稠度、碱消值方面,品种间的差异显著大于部位间的差异;三种内源激素在不同部位籽粒中的差异均较为显著,且SA差异最为稳定,全周期不同部位间籽粒SA含量变异系数为0.59%~25.90%,ABA和IAA变异系数依次为1.35%~66.25%和0.16%~81.04%。不同部位SA含量在灌浆初期与末期正相反,前期表现为上部籽粒>中部>下部,末期却表现为下部籽粒>中部>上部。不同部位籽粒的内源激素及产量等相关性状差异均较为显著,激素含量差异以SA最为稳定。SA与ABA和IAA分别呈显著正相关(=0.54,< 0.05)和极显著正相关(=0.59,< 0.01),与千粒重呈显著负相关(=−0.53,< 0.05),其与长宽比和整精米率也分别为极显著负相关(= −0.66,< 0.01)和极显著正相关(= 0.95,< 0.01)。
粒位;籽粒生物量;内源激素;差异性;相关性
水稻是我国主要粮食作物之一,水稻产量、品质及中间产物均受基因与环境共同调控[1-6],理论上同一品种在同一环境下稻米产量及品质应无差别,但实际检测结果却略有差异。杨建昌等[7]对颖花开花时间与胚乳发育的关系进行研究,发现胚乳细胞增殖速度和胚乳细胞数的差异是造成不同时期开花籽粒粒重差异的直接原因,而不同部位间籽粒颖花的开花时间是存在显著差异的。陈新红等[8]认为,一般情况下稻穗不同部位的结实特性均表现为上部>中部>下部且存在显著差异。袁莉民等[9]研究表明不同部位籽粒胚乳结构形态的建成顺序是不同的,基本表现为上部籽粒早于中部籽粒和下部籽粒。Wobus等[10]研究认为不同部位籽粒发育的差异与内源激素含量的关系较为密切。Yang等[11]通过研究发现水稻灌浆期,尤其是灌浆前期,不同部位的内源激素确实存在显著差异。水稻籽粒中的各项生命活动都是由植物内源激素调节的,其中较为常见的内源激素有生长素(IAA)、脱落酸(ABA)、水杨酸(SA)等,这些内源激素在植物的生长发育过程中起着非常重要的作用,如SA可直接参与水稻开花、灌浆和离子转运等过程,从而影响植物水分代谢、矿质营养吸收和光合作用。冯志威[12]研究表明,外源SA提高谷子千粒重、穗粒重及产量,以及可溶性糖、可溶性蛋白含量和硝酸还原酶、SOD、POD活性。近年来,关于ABA的研究主要集中在ABA通过促进和抑制酚类物质的氧化而影响种子的休眠和萌发上,但也有研究表明ABA在灌浆初期有利于初期籽粒中ADPG焦磷酸酶和淀粉合成酶活性及淀粉含量增加,当ABA含量增加后,弱势粒灌浆速率显著加快,最大灌浆速率提前[13]。也有研究表明IAA 是内源激素中唯一的一个具有极性运输特点的调节激素,它总是从植物形态学上端向下端运输,不能反向运输,且对光合作用物质分配的调节具有定向的影响,张浩等[14]认为,穗中IAA含量的增加能够促进可输配物质的形成,抑制暂不输配物质的产生,对结构物质、呼吸消耗和光合产物输出速度有交替作用。因此,笔者认为造成水稻不同部位籽粒品质及产量结果差异的部分原因为籽粒内源激素差异。本研究旨在通过分析不同部位籽粒产量等相关性状与内源激素的差异及相关性以论证不同部位籽粒产量与相关品质性状差异的机理。同时也为后续栽培及育种工作提供一定的参考依据,通过缩小部位间的差异来提升水稻整体的品质水平。
1 材料与方法
1.1 实验材料
供试水稻品种为浙江省种植业管理局推荐的常规粳稻品种嘉58、嘉67、浙粳99、秀水121、秀水134和中嘉8号共6个品种,种子由浙江省勿忘农种业集团提供。
试验于2019年在杭州市富阳区中国水稻研究所试验场进行。耕层土壤的pH值为4.95,土壤有机质为22.88 g/kg,碱解氮为180.53 mg/kg,有效磷为4.89 mg/kg,速效钾为223.77 mg/kg。
1.2 田间管理
试验地为常规田块,每个地块为2.5 m×5 m的长方形田块。5月15日播种,6月14日移栽,行穴距为20 cm×20 cm,每穴3株苗,8月31日抽穗,9月6日开始第一次取样试验,每隔一周进行一次取样分析,灌浆期共取样6次,10月18日完成收割。水稻生长期间内进行统一的田间管理。抽穗后立即采用挂牌法对同步抽穗的稻穗进行标记。
1.3 取样
抽穗后7 d开始取样测试,每7 d取同日开花的稻穗各15穗直至水稻成熟。取样时间为11:00至13:00。利用液氮取灌浆期稻穗15穗,样品取下后先放置于盛有液氮的容器中并尽快转入–80℃冰箱。据田间试验统计,所有试验品种稻穗的枝梗结数量均为8~11。将稻穗按上、中、下分成三部分,从稻穗顶部往下的3个枝梗结为分割点,分割点以上的籽粒为上部籽粒(Top Spikelets,TS),从稻穗底部往上的3个枝梗结为分割点,分割点以下的籽粒为下部籽粒(Bottom Spikelets,BS),中间2~5个枝梗结上的籽粒为中部籽粒(Middle Spikelets,MS)(图1)。

图1 水稻穗结构
Fig. 1. Schematic representation of a rice panicle.
1.4 测定项目及方法
利用美国SPEX 6875 液氮研磨仪将样品研磨至粉状后−80℃冰箱保存备测。采用《水稻中43种植物激素的测定液相色谱-串联质谱法》(NY/T 2871−2015)对各时期样品中的水杨酸、脱落酸和吲哚乙酸含量进行检测,对灌浆期各时间点的生物量进行测定,水稻成熟后对种子进行考种,分析产量等性状。根据《食用稻品种品质》对不同部位籽粒品质进行测定,同时分析其品质差异。植物激素、产量性状考查和稻米品质测定均设置3个重复。
1.5 统计分析方法
数据的整理及统计分析采用Excel 2010、SPSS 19.0数据处理系统软件,差异性分析采用DPS 7.05,检验方法为检验。
2 结果与分析
2.1 不同部位籽粒鲜质量动态变化
从图2中可以看出,灌浆期不同部位籽粒鲜质量差异较为显著,籽粒从花后7 d至28 d灌浆较快,粒重显著增加。花后35 d至42 d,各部位籽粒进入灌浆后期,籽粒生物量的差距逐渐缩小但部分品种仍有显著差异。
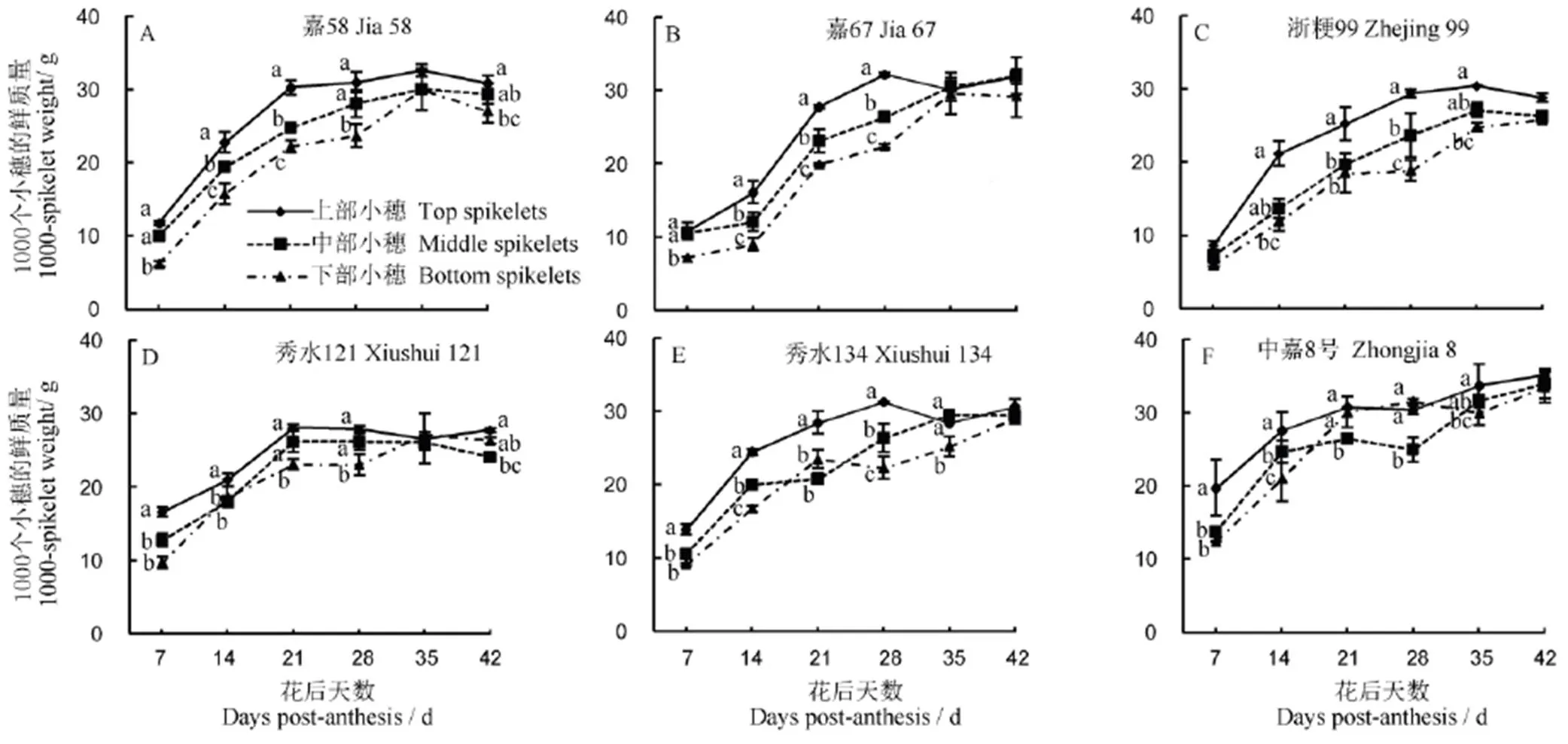
同一时段内相同小写字母表示不同部位籽粒间差异未达显著水平(P<0.05)。
Fig. 2. Dynamic changes of fresh grain weight in different parts of panicle.
2.2 不同部位籽粒千粒重差异
从图3来看,除秀水121外,各品种千粒重基本表现为上部籽粒大于中下部,其情况与图2中各品种花后42 d的籽粒鲜质量较为相似,且各品种部位间千粒重均有不同程度的差异。嘉58、浙粳99和嘉67上、中、下三个部位籽粒千粒重差异最为显著,其品种不同部位间千粒重的变异系数分别为6.50%、6.23%和5.07%。秀水134和中嘉8号基本表现为上部与中下部存在显著差异且变异系数分别为4.46%和3.26%。秀水121表现为仅上部与中部存在显著差异,但其变异系数最低,仅为2.12%。
2.3 不同部位籽粒整精米率与长宽比
各品种不同部位籽粒整精米率如图4-A所示,不同部位间籽粒均无显著性差异的品种仅为中嘉8号。嘉58、秀水121和秀水134表现为上、中、下部位籽粒均有显著差异,嘉67和浙粳99表现为上部与下部位籽粒存在显著差异。从不同部位整精米率的表现形式来看,嘉58和秀水134表现为中部籽粒整精米率最高,秀水121却表现为中部最低,嘉67和浙粳99表现为上部最高,中部次之。从部位间变异系数来看,秀水121品种部位间变异系数最大,为9.44%,其次为秀水134,变异系数为6.10%,嘉58为4.90%,嘉67、浙粳99和中嘉8号变异系数分别为1.44%、2.08%和1.07%。
各品种不同粒位的籽粒长宽比如图4-B所示,各粒位的籽粒均无显著差异的品种有秀水121和秀水134,嘉58、嘉67和中嘉8号下部籽粒与中、上部位籽粒长宽比存在显著差异,浙粳99则表现为上部籽粒与下部位籽粒存在显著差异。不同粒位的籽粒长宽比变异系数并不显著,变异系数最大为嘉67,为2.05%;最小者为秀水134,变异系数为0%,其余品种变异系数均小于2.05%。但品种间的变异系数较大,高达17.40%。
2.4 不同部位籽粒胶稠度与碱消值
不同部位胶稠度指标如图5-A所示,嘉58、嘉67和秀水121等3个品种不同部位籽粒均无显著差异,但秀水134和中嘉8号不同部位籽粒差异均较为明显,其检测值分别为55.00~74.50和57.00~73.00,变异系数分别为15.04%和12.67%。浙粳99上部籽粒与中下部籽粒差异较为显著,其变异系数为5.84%,其余品种变异系数为1.49%~3.81%,品种间变异系数为6.35%(表1)。
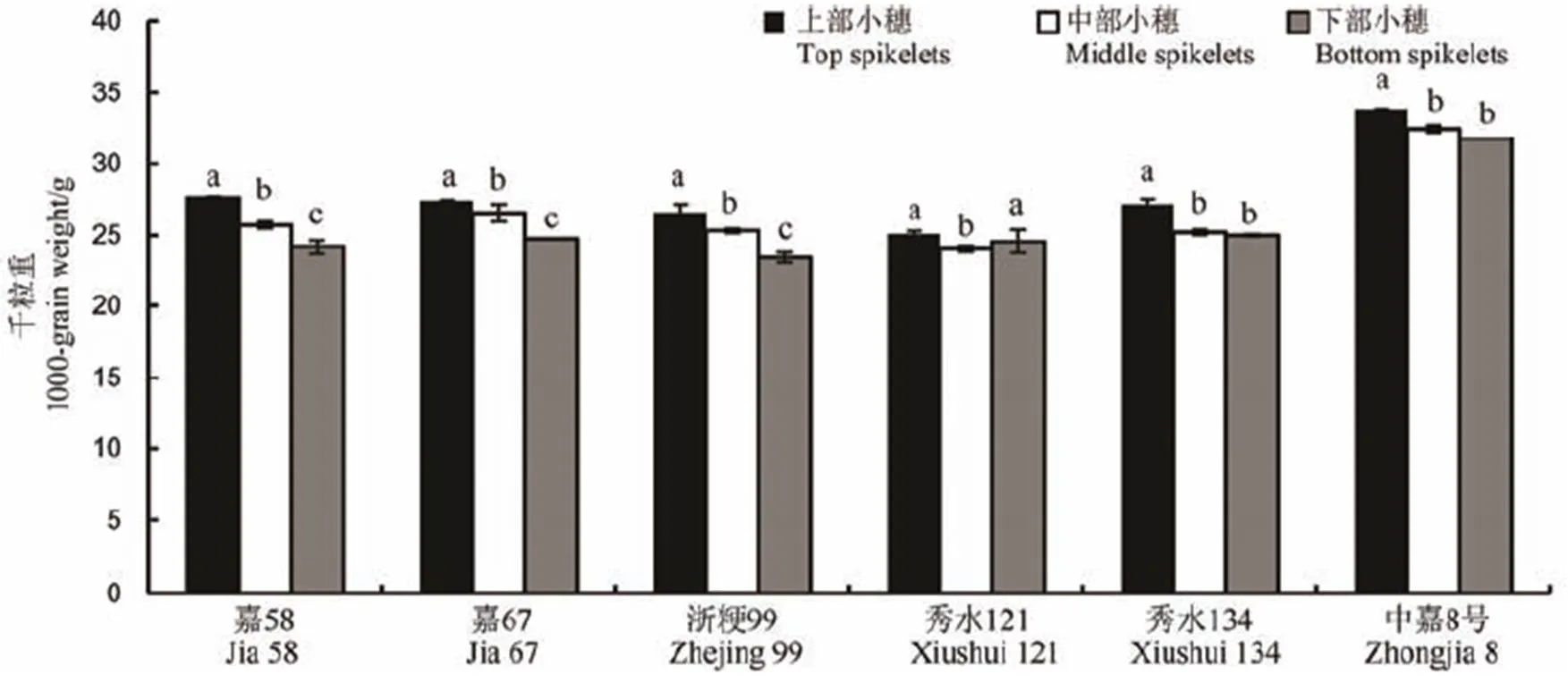
相同小写字母表示不同部位籽粒间差异未达显著水平(P<0.05)。下同。
Fig. 3. Difference in 1000-grain weight in different parts.
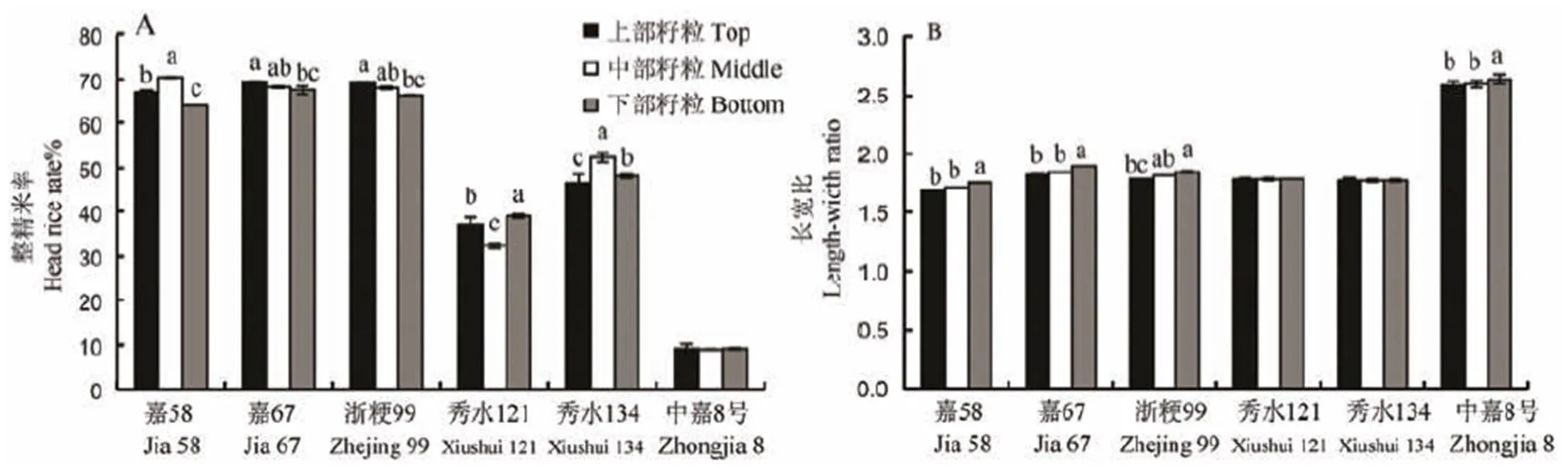
图4 稻穗不同部位籽粒间整精米率和长宽比的差异
Fig. 4. Difference in head rice yield and length-width ratio of seeds in different parts.
不同部位碱消值性状如图5-B所示。浙粳99、秀水134、中嘉8号等3个品种各粒位籽粒均不存在显著差异,其含量值为6.75~7.00,嘉58表现为上部籽粒与中下部籽粒存在显著差异,嘉67表现为中部与下部存在显著差异,秀水121表现为上部与下部籽粒存在显著差异。从不同粒位碱消值来看,嘉58和秀水134表现为中部籽粒碱消值最高,其余品种均表现为下部籽粒碱消值最高。从变异系数来看,嘉58品种变异系数最大为2.57%,嘉67和秀水121次之,变异系数均为1.86%,其余品种变异系数为0.70%~1.23%,品种间变异系数与部位间差异不大,仅为1.22%。
2.5 不同部位间籽粒水杨酸含量的变化及差异
水杨酸(SA)为植物体内最为常见的内源激素之一,在不同植物或相同植物不同生长发育时期内都具有较大的差异。针对SA在植物体内的作用,前人已做了大量的基础研究,发现其在植物的生长、发育、代谢等阶段均存在不同程度的作用[15]。从图6中可以看出,SA激素含量随灌浆的进行呈倒V字型变化,总体为先升高后降低。花后7 d各品种SA含量为0.61~1.46 mg/kg,花后3 d(约为灌浆中期)达到顶峰,为1.27~4.92 mg/kg,42 d后下降至0.41~1.15 mg/kg。从部位间与品种间的差异来看,品种间的差异显著大于部位间的差异。嘉58上、中、下部位从灌浆初期至中期,SA含量分别上升了266.67%,371.57%和486.59%,平均上升了374.94%。中嘉8号的上、中、下部位从灌浆初期至中期,SA含量分别上升了126.23%、73.97%和73.33%,该品种平均上升了91.18%。另外,SA含量在部位间的差异虽不及品种间,但结果显示同期部位间籽粒SA含量也存在一定的差异。表2显示,从花后7 d至花后42 d,SA含量在6个品种不同部位间的变异系数依次为10.89%~19.20%,1.81%~12.80%,0.59%~21.14%,4.67%~16.49%,3.01%~8.85%和1.47%~16.66%,品种间的变异系数依次为24.80%、16.15%、45.11%、41.26%、34.88%和31.22%,从变异系数中也可以看出不同部位间的差异不及品种间的差异(表2)。
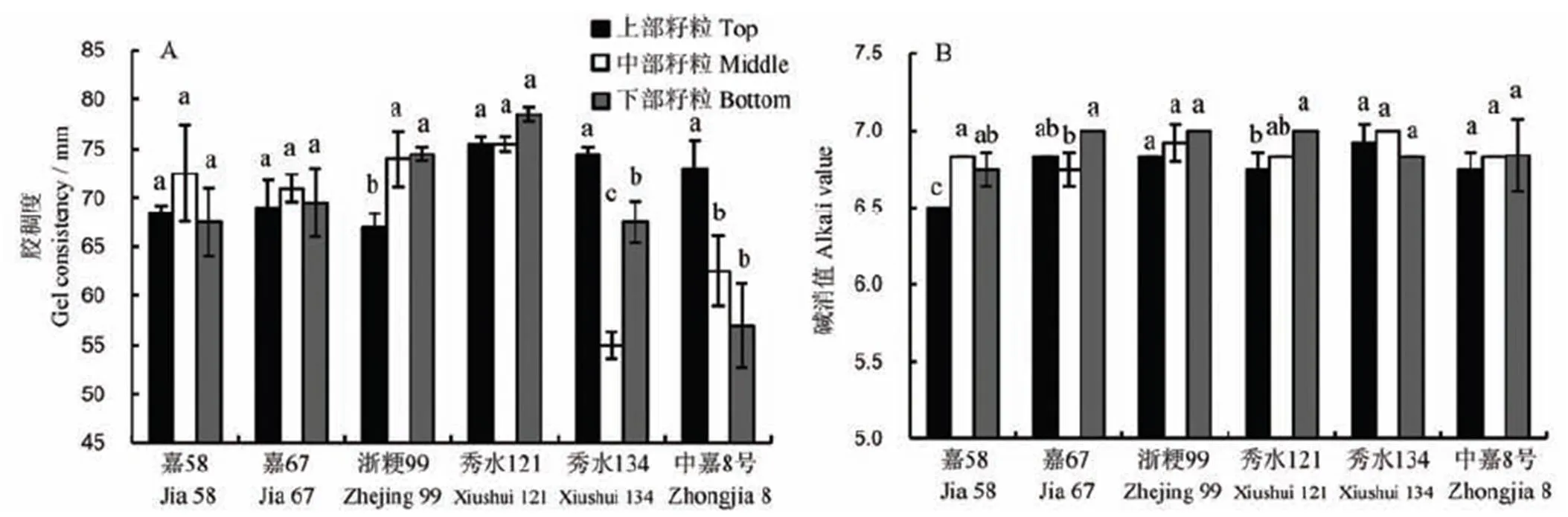
图5 稻穗不同部位籽粒胶稠度和碱消值差异
Fig. 5. Difference in gel consistency and alkali value of seeds in different parts.

表1 籽粒品质性状在不同部位和品种间的变异系数
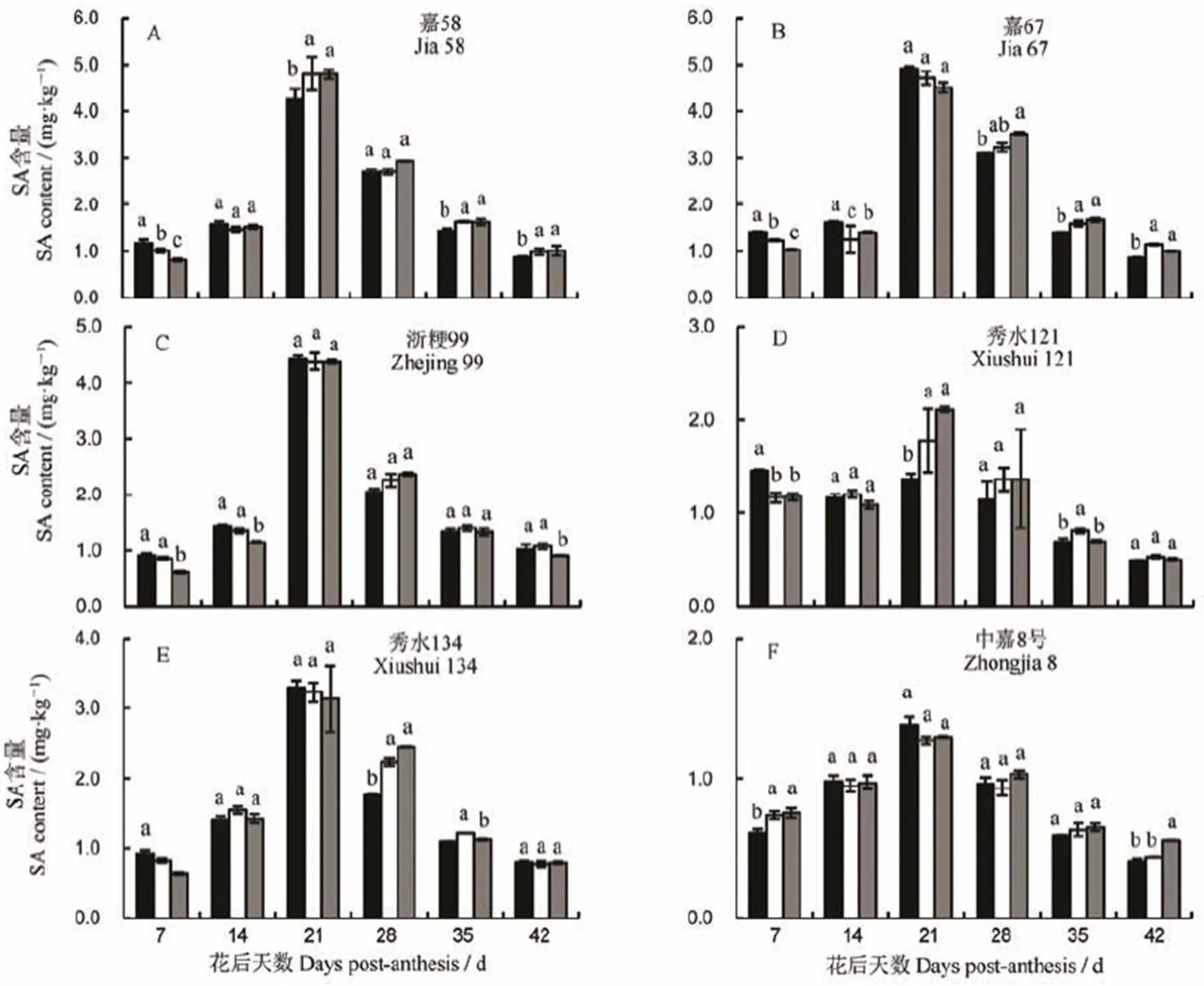
同一时段内相同小写字母表示不同部位籽粒间差异未达0.05显著水平。
Fig. 6. Dynamic changes and differences of SA content in grains of different parts.
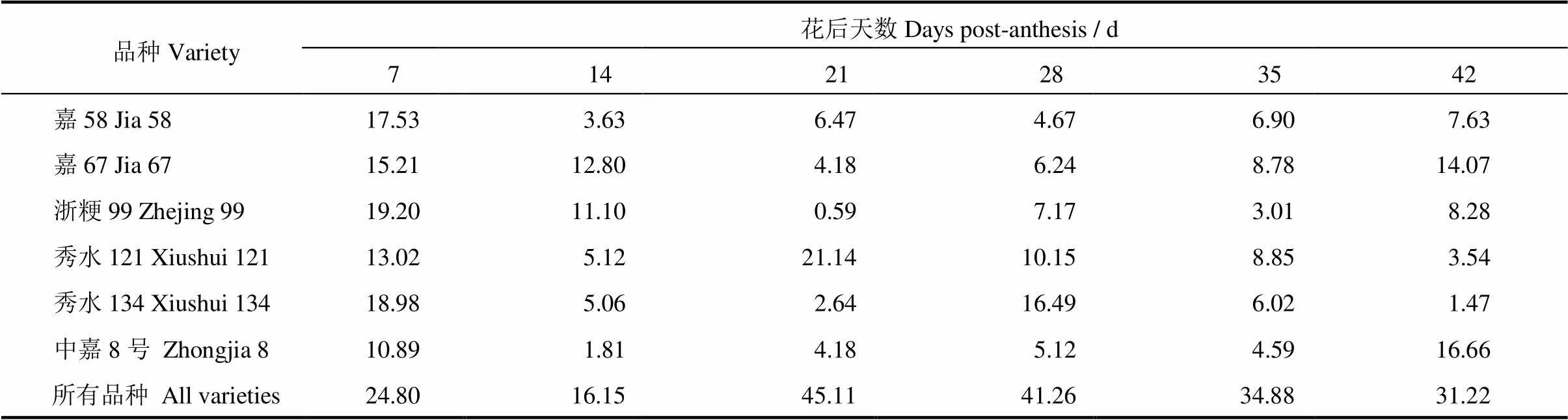
表2 籽粒SA含量在不同粒位间和品种间的变异系数
另外,从整个灌浆周期的SA动态图中可以看出,SA含量差异在灌浆初期和末期相对于中期更为显著,初期基本表现为上部>中部>下部,灌浆末期则表现为下部>中部>上部。灌浆中期多数品种SA含量差异并不显著。
2.6 不同部位籽粒脱落酸的差异及变化
脱落酸(ABA)在抗逆性方面与SA较为相似,寒冷、干旱、高温、高盐和水淹的逆境条件下都能促使植物体内的ABA迅速增加[16]。表3显示,ABA含量与SA有所不同,但也存在相似之处。不同之处表现为不同部位ABA含量差异性并不显著,虽个别品种某个阶段不同部位变异系数较大,但均未出现上中下三个部位均存在显著差异的情况。相似之处为品种间的差异仍大于多数品种部位间的差异。如同为花后7 d,品种间的变异系数为43.55%,部位间的变异系数为6.55%~45.66%,仅有浙粳99部位间的变异系数(45.66%)大于品种间的变异系数(43.55%),其余品种部位间的变异系数均小于品种间的变异系数。
2.7 不同部位籽粒吲哚乙酸含量的差异及变化
如图7所示,所有水稻品种的吲哚乙酸(IAA)含量在灌浆期间呈倒V字型,在花后21 d左右含量达到最高,随后逐渐降低。其含量动态表现与曹转勤等[17]研究较为相似,IAA含量在灌浆初期较少,随着灌浆过程的逐步发生,IAA含量逐渐上升至最大值后逐渐后降低。其激素含量后期下降趋势与其他内源激素有所差异,较SA与ABA更为缓慢,直至灌浆后期,其含量仍处于较高水平。部分品种含量差异较大,如同在花后21 d,嘉58 IAA含量为1.29~1.42 mg/kg,嘉67为2.29~3.08 mg/kg。部位间的差异也较为明显,尤其为嘉67与浙粳99所有时段均有不同程度的显著差异。表4对各时期部位间及品种间IAA含量差异进行了验证,数据显示从花后7 d时,不同部位间IAA含量差异最大,其变异系数高达31.73%~81.04%,在灌浆中期和末期,各品种部位间的差异较为稳定,尤其在灌浆末期,除嘉67外,其余品种不同部位籽粒IAA含量变异系数仅为3.62%~6.28%。
2.8 内源激素、产量与品质的相关性
对内源激素、产量与品质等相关参数进行相关性分析,其中SA、ABA、IAA均采用6个动态数据之和进行分析,千粒重、长宽比、整精米率、胶稠度和碱消值为籽粒完全成熟后的实验数据。
由表5显示,三类内源激素表现为显著正相关或极显著正相关。千粒重、长宽比和整精米率之间,仅千粒重和长宽比为极显著正相关,整精米率与千粒重和长宽比之间均为极显著负相关。胶稠度和碱消值与内源激素、千粒重、长宽比和整精米率间均无显著相关性。另外,SA与千粒重和长宽比呈极显著负相关,与整精米率呈极显著正相关,IAA与整精米率同呈显著正相关。

表3 不同粒位籽粒的ABA含量差异
J58−嘉58; J67−嘉67; Z99−浙粳99; X121−秀水121; X134−秀水134; Z8−中嘉8号。TS−上部籽粒;MS−中部籽粒;BS−下部籽粒。
J58, Jia 58; J67, Jia 67; Z99, Zhejing 99; X121, Xiushui 121; X134, Xiushui 134; Z8, Zhongjia 8. TS, Top spikelets; MS, Middle spikelets; BS, Bottom spikelets.

图7 不同部位籽粒IAA含量动态变化及差异
Fig. 7. Dynamic changes and differences of IAA content of grains in different parts.

表4 籽粒IAA含量在不同部位和品种间的变异系数
3 讨论
3.1 不同部位稻米品质的差异及其与产量性状的关系
自20世纪以来,如何提高产量与品质一直被育种家们所关注,但在水稻实际生产中,限制其品质和产量的因素较多,想要同时实现优质与高产仍存在较大的难度。因此,有关水稻产量与品质性状间关系的研究也逐步增加。我们对不同品种和不同部位间籽粒整精米率与长宽比及胶稠度与碱消值性状的差异性进行了研究,结果表明不同品种和粒位间的稻米品质均有差异,穗上部与下部籽粒的整精米率差异较大,下部籽粒的长宽比与其他部位差异较大,这一结果与王嘉宇等[18]的研究结论部分相似,王嘉宇等认为水稻下部与上中部的籽粒的长宽比差异显著,下部与中部籽粒的整精米率差异较大。本研究显示整精米率在中下部位有差异,但上部与下部籽粒也存在显著差异。品种间胶稠度的差异更甚于部位间,实验结果显示秀水134、中嘉8号和浙粳99不同粒位间有显著差异,但嘉58、嘉67和秀水121则表现为不同粒位均无显著差异。董明辉等[19]对一、二次枝梗不同粒位籽粒胶稠度作了研究,结果表明不同品种及不同部位的胶稠度均有所差异且规律有所不同,胶稠度指标差异多来源于品种及施氮量的差异。蔡一霞等[20]研究结论也表明胶稠度及RVA谱动态变化在不同部位间存在显著差异。碱消值的差异结果与胶稠度相反,表现为嘉58、嘉67和秀水121等品种粒位间存在不同程度差异,秀水134、浙粳99和中嘉8号却表现为无显著差异。在品质与产量性状关系上,其相关性如表5所示,胶稠度、碱消值与千粒重为负相关,但相关未达显著。千粒重与长宽比呈极显著正相关,与整精米率呈极显著负相关。其结论与丛琳[21]研究结论相似,其认为整精米率与产量呈负相关关系,产量越高,则稻米的加工品质越低。
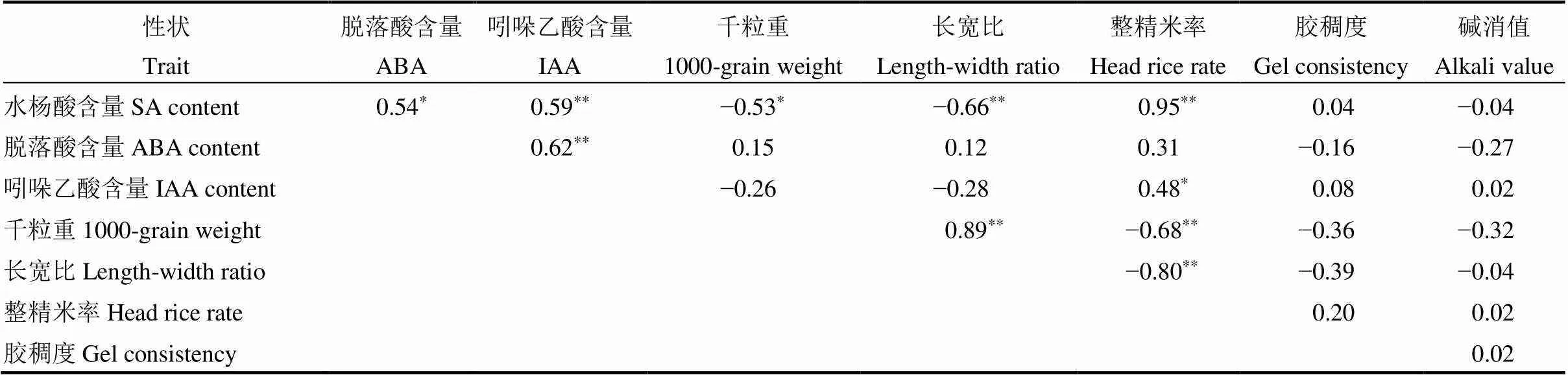
表5 内源激素、产量与品质的相关性
*在0.05水平显著相关;**在0.01水平显著相关。
*Significantly correlated at<0.05;**Significantly correlated at<0.01.
3.2 不同部位内源激素的差异及其与产量性状的关系
植物激素是由植物自身代谢产生的一类有机物质,并从产生部位转移到作用部位。植物激素的结构虽已被人所知,但其的作用机理却非常复杂,专家学者们仍在坚持探索。水杨酸(SA)、脱落酸(ABA)和吲哚乙酸(IAA)等激素被研究得较为广泛。SA为一种内源酚类生产调节剂,在水稻体内较为常见,在水稻的生长发育、种子萌发以及叶片的光合作用等方面均发挥着重要的作用。现阶段,关于SA对水稻的研究多为SA对生长抗逆性[22]和促进种子萌发[23]的作用上,尤其为SA在高温胁迫下的作用等研究[24-26]。但实验表明,SA在促进水稻籽粒灌浆和发育上也有重大的作用,不同部位SA含量与籽粒的发育情况密切相关。周万海等[27]研究发现,外施0.25 mmol/LSA能有效提高苜蓿幼苗的株高、根长和鲜质量,王宝增等[28]认为0.6 mmol/LSA对盐胁迫下沙打旺幼苗株高和干质量的增加效果最好。ABA在水稻体内主要以游离态的形式存在,其主要作用在水稻籽粒的发育和成熟过程中[29-30],并在水稻籽粒的发育和成熟过程中发挥作用。Lee等[31]研究认为ABA既可以促进同化物向籽粒运转,又可加速成熟,与灌浆前、中期相比,后期ABA能促进籽粒成熟。张上隆等[32]认为ABA通过调节库中酸性转化酶的活性促进蔗糖分解为葡萄糖和果糖,从而促进库对蔗糖的吸收和卸载。国外研究也存在类似发现。Kato等[33]发现在水稻籽粒灌浆过程中,大的籽粒比小的籽粒ABA含量高。IAA的作用相对更为单一,其主要作用为促进细胞生长发育,其游离态的IAA是植物中唯一的具有极性运输的激素,通过增加气孔的开放作用从而增强叶片的光合作用[34]。IAA在植物生长发育过程中还能促进细胞分裂、体积增大、形态建成,例如促进植物侧根和不定根的形成[35]。
本研究对SA、ABA和IAA在水稻整个灌浆期内各时段的含量进行了比较,其中SA总体变化为先上升后下降的过程,不同部位籽粒SA差异较为明显,初期基本表现为上部>中部>下部,中期差异性逐渐减小,末期表现为下部>中部>上部。根据不同部位籽粒SA动态变化可以得出SA含量在整个周期内具有显著的规律性,由此可以看出,SA对不同部位籽粒生物量具有一定的影响。对内源激素与产量的相关性进行分析,结果如表5所示,内源SA含量与千粒重呈显著负相关。有关SA对产量影响方面,沙汉景等[36]和杨军等[37]进行了相似研究,但均为外源激素对产量的影响。杨军等[37]认为外源适量添加500 μmol/LSA,增产效果最佳,仅添加100 μmol/L,增产效果并不显著。沙汉景等[36]研究也认为外源适量水杨酸在植物生长发育中其着重要的促进作用。在内源SA对千粒重或产量的影响方面,未见相关报道。对此,结合前人的研究,我们认为,内源SA与外源添加SA的作用有所差异,外援适量添加SA可促进籽粒增重,但内源SA起抑制作用。
在ABA含量的动态研究中,段俊等[38]认为在籽粒灌浆前期,上部籽粒中的ABA含量略大于下部籽粒,在灌浆后期时正好相反,下部籽粒中的ABA含量略大于上部籽粒。但本研究结果与段俊等人研究略有差异,结果显示不同部位ABA含量在灌浆初期至中期均无明显差异,但灌浆后期基本表现为下部>中部>上部。ABA的特性是通过促进籽粒脱水和营养物质积累从而促进籽粒发黄成熟。从同一穗部籽粒的发育情况可知,穗下部籽粒的灌浆时间较上部籽粒少7 d左右。因此,我们推测灌浆后期各部位不同含量ABA的作用主要是调节不同部位籽粒的成熟度,灌浆后期ABA表现为下部>中部>上部,促进中下部位籽粒能够更快成熟,从而促进各部位籽粒能够同时从乳熟期向蜡熟期转变。IAA的动态变化虽为先上升,花后21 d左右达到顶峰后逐渐下降,但其下降的速率相对SA更为缓慢,直至灌浆后期,其含量仍为顶峰时期含量的33.60%~84.37%。但在IAA是否会对籽粒灌浆速率产生显著影响的研究上,结合IAA与鲜生物量的动态变化,我们认为灌浆初期IAA含量与生物量积累存在一定的相关性,但在灌浆中期及末期,相关性并不显著。段俊等[38]研究也认为IAA含量与水稻的灌浆速率并无显著的相关性。萧浪涛等[39]研究认为灌浆初期IAA与起始灌浆势、平均灌浆速率和籽粒的充实率呈显著相关,且中期及末期确无显著相关性。但这与陶龙兴等[40]的结论不完全一致。因此推测并非灌浆初期上部籽粒IAA含量以及籽粒灌浆的速率均高于下部籽粒,其规律同时也受品种的影响,因品种而异。而曹转勤等[17]通过突变体材料试验认为IAA含量与灌浆的速率呈正相关且IAA含量在部位间的差异并不显著。
另外,对比激素间的关系,SA含量与ABA、IAA含量分别呈显著正相关和极显著正相关,且ABA与IAA含量之间也呈极显著正相关,这一结果意味着三者之间存在一定的“激励”作用,但数据显示,ABA、IAA与产量并无显著相关性,这也验证了前文对IAA动态分析讨论中提及的IAA对籽粒灌浆的作用时间较长,其对灌浆速率可能产生影响但对产量不存在显著影响。
[1] 王慧, 方玉, 黄艳玲, 冯冲, 杨力, 周桂香, 张从合. 稻米主要食味品质基因型与环境互作分析及其相关性研究[J]. 中国稻米, 2020, 26(2): 23-26.
Wang H, Fang Y, Huang Y L, Feng C, Yang L, Zhou G X, Zhang C H. Analysis of the genotype and environment interactions as well as the correlation research of main taste quality in rice[J]., 2020, 26(2): 23-26. (in Chinese with English abstract)
[2] Lin Z M, Zhang X H, Wang Z X, Jiang Y T, Liu Z H, Alexander D, Li G H, Wang S H, Ding Y F. Metabolomic analysis of pathways related to rice grain chalkiness by a notched-belly mutant with high occurrence of white-belly grains[J]., 2017, 17: 39.
[3] 贾小丽, 叶江华, 苗利国, 林红梅, 林文雄. 水稻籽粒灌浆速率的发育遗传机制研究[J]. 热带作物学报, 2012, 33(2): 622-626.
Jia X L, Ye J H, Miao L G, Lin H M, Lin W X. Developmental genetic mechanism research on grain- filling rate in rice[J]., 2012, 33(2): 622-626. (in Chinese with English abstract)
[4] 朱庆森, 曹显祖, 骆亦其. 水稻籽粒灌浆的生长分析[J]. 作物学报, 1988, 14(3): 182-192.
Zhu Q S, Cao X Z, Luo Y Q. Growth analysis of grain filling in rice[J]., 1988, 14(3): 182-192. (in Chinese)
[5] 户少武, 张欣, 景立权, 赖上坤, 王云霞, 朱建国, 王余龙, 杨连新. 高浓度CO2对稻穗不同位置籽粒结实和米质性状的影响[J]. 应用生态学报, 2019, 30(11): 3725-3734.
Hu S W, Zhang X, Jing L Q, Lai S K, Wang Y X, Zhu J G, Wang Y L, Yang L X. Effects of elevated CO2concentration on grain filling capacity and quality of rice grains located at different positions on a panicle[J]., 2019, 30(11): 3725-3734. (in Chinese with English abstract)
[6] Kusano H, Arisu Y, Nakajima J, Yaeshima M, Shimada H. Implications of the gene for F1–ATPase β subunit (AtpB) for the grain quality of rice matured in a high-temperature environment[J]., 2016, 33(3): 169-175
[7] 杨建昌, 刘立军, 王志琴, 郎有忠, 朱庆森.稻穗颖花开花时间对胚乳发育的影响及其生理机制[J]. 中国农业科学, 1999, 32(3): 44-51.
Yang J C, Liu L J, Wang Z Q, Lang Y Z, Zhu Q S. Effect of flowering time of spikelet on endosperm development and its physiological mechanism[J]., 1999, 32(3): 44-51. (in Chinese)
[8] 陈新红, 韩正光, 张安存, 叶玉秀, 周青, 吕宏飞. 不同施氮量与种植密度对水稻穗上不同部位结实特性的影响[J]. 浙江农业学报, 2014, 26(6): 1578-1582.
Chen X H, Han Z G, Zhang A C, Ye Y X, Zhou Q, Lv H F. Analysis on rice grain filling characteristics in different spikelet positions under different nitrogen rates and planting densities[J]., 2014, 26(6): 1578-1582. (in Chinese with English abstract)
[9] 袁莉民, 展明飞, 章星传, 王志琴, 杨建昌. 水稻穗上不同粒位籽粒胚乳结构及其结实期灌溉方式对它的调控作用[J]. 作物学报, 2018, 44(2): 245-259.
Yuan L M, Zhan M F, Zhang X C, Wang Z Q, Yang J C. Endosperm structure of grains at different positions of rice panicle and regulation effect of irrigation regimes on it during grain filling[J]., 2018, 44(2): 245-259. (in Chinese with English abstract)
[10] Wobus U, Weber H. Seed maturation: Genetic programmers and control signals[J]., 1999, 2(1): 33-38.
[11] Yang J C, Peng S B, Visperas R M, Sanico A L, Zhu Q S, Gu S L. Grain filling pattern and cytokinein content in the grains and roots of rice plants[J]., 2000, 30(3): 261-270.
[12] 冯志威. 灌浆期喷施植物生长调节剂对谷子产量及生理特性的影响[D]. 太原: 山西农业大学, 2017.
Feng Z W. Effects of plant growth regulator on yield and physiological mechanism of foxtail millet in filling stage[D]. Taiyuan: Shanxi Agricultural University, 2017. (in Chinese with English abstract)
[13] 杨建昌, 王志琴, 朱庆森, 苏宝林. ABA与GA对水稻籽粒灌浆的调控[J]. 作物学报, 1999, 25(3): 341-348.
Yang J C, Wang Z Q, Zhu Q S, Shu B L. Regulation of ABA and GA to the grain filling of rice[J]., 1999, 25(3): 341-348. (in Chinese with English abstract)
[14] 张浩, 罗时石, 龚荐, 葛才林, 马飞. 吲哚乙酸对水稻花后剑叶光合产物输配作用的示踪动力学研究[J]. 江苏农学院学报, 1996(1): 31-36.
Zhang H, Luo S S, Gong J, Ge C L, Ma F. Study on the isotope kinetics of the photosynthetic of flag leaf of post floral rice and IAA affection on it[J]., 1996(1): 31-36. (in Chinese with English abstract)
[15] 赵丹, 纪鹏, 何晓蕾, 景艳丽, 张涛, 王洪义. SA对盐胁迫下草本植物种子萌发及幼苗生理特性影响的研究进展[J]. 黑龙江八一农垦大学学报, 2020, 32(3): 7-12.
Zhao D, Ji P, He X L, Jing Y L, Zhang T, Wang H Y. Research progress of effect of SA on seed germination and seedling physiological characteristics of herbaceous plants under salt stress[J]., 2020, 32(3): 7-12. (in Chinese with English abstract)
[16] 任菲, 张荣佳, 陈强, 白艳波, 黄菲, 李雪梅. ABA和SA对于提高植物抗旱及抗盐性的研究进展[J]. 生物技术通报, 2012(3): 17-21.
Ren F, Zhang R J, Chen Q, Bai Y B, Huang F, Li X M. Progress in ABA and SA improving plant drought resistance and salt resistance[J]., 2012(3): 17-21. (in Chinese with English abstract)
[17] 曹转勤. 水稻几个突变体的籽粒灌浆特征及其与内源激素的关系[D]. 扬州: 扬州大学, 2014.
Cao Z Q. Grain filling characteristics and their relations with endogenous hormones in several rice mutants[D]. Yangzhou: Yangzhou University, 2014. (in Chinese with English abstract)
[18] 王嘉宇, 徐正进, 张世春, 陈温福. 水稻穗不同部位籽粒品质性状差异的比较[J]. 华北农学报, 2008, 23(1): 96-100.
Wang J Y, Xu Z J, Zhang S C, Chen W F. Comparison in quality traits at different parts within a rice panicle[J]., 2008, 23(1): 96-100. (in Chinese with English abstract)
[19] 董明辉, 桑大志, 王朋, 王学明, 杨建昌.不同施氮水平下水稻穗上不同部位籽粒的蒸煮与营养品质变化[J]. 中国水稻科学, 2006(4): 389-395.
Dong M H, Shang D Z, Wang P, Wang X M, Yang C J. Changes in cooking and nutritional qualities of grains at different positions within a rice panicle under different nitrogen levels[J]., 2006(4): 389-395. (in Chinese with English abstract)
[20] 蔡一霞, 刘春香, 王维, 张洪熙, 张祖建, 杨静, 唐汉忠. 灌浆期表观直链淀粉含量相似品种稻米胶稠度和RVA谱的动态差异[J]. 中国农业科学, 2011, 44(12): 2439-2445.
Cai Y X, Liu C X, Wang W, Zhang H X, Zhang Z J, Yang J, Tang H Z. Dynamic differences of the RVA profile and gel consistency in two rice varieties with similar apparent amylose content during grain filling[J]., 2011, 44(12): 2439-2445. (in Chinese with English abstract)
[21] 丛琳. 辽宁不同时期水稻品种产量与品质特性研究[D].沈阳: 沈阳农业大学, 2018.
Cong L. study on the yield and quality characteristics of rice varieties in Liaoning at different stage[D]. Shenyang: Shenyang Agricultural University, 2018. (in Chinese with English abstract)
[22] 张蕊, 高志明, 吕俊, 王三根. 外源水杨酸对水稻幼苗耐寒性的影响[J]. 江苏农业科学, 2012, 40(6): 62-65.
Zhang R, Gao Z M, Lv J, Wang S G. Effect of exogenous salicylic acid on cold tolerance of rice seedling[J]., 2012, 40(6): 62-65. (in Chinese with English abstract)
[23] 王俊斌, 王海凤, 刘海学. 水杨酸促进盐胁迫条件下水稻种子萌发的机理研究[J]. 华北农学报, 2012, 27(4): 223-227.
Wang J B, Wang H F, Liu H X. Study on mechanism of salicylic acid on the promotion of rice seeds germination under salt stress[J]., 2012, 27(4): 223-227. (in Chinese with English abstract)
[24] Feng B H, Zhang C X, Chen T T, Zhang X F, Tao L X, Fu G F. Salicylic acid reverses pollen abortion of rice caused by heat stress[J]., 2018, 18(1): 1-16
[25] 符冠富, 张彩霞, 杨雪芹, 杨永杰, 陈婷婷, 赵霞, 符卫蒙, 奉保华, 章秀福, 陶龙兴, 金千瑜. 水杨酸减轻高温抑制水稻颖花分化的作用机理研究[J]. 中国水稻科学, 2015, 29(6): 637-647.
Fu G F, Zhang C X, Yang X Q, Yang Y J, Chen T T, Zhao X, Fu W M, Feng B H, Zhang X F, Tao L X, Jin Q Y. Action mechanism by which SA alleviates high temperature-induced inhibition to spikelet differentiation[J]., 2015, 29(6): 637-647. (in Chinese with English abstract)
[26] Mohammed A R, Tarpley L. Effects of night temperature, spikelet position and salicylic acid on yield and yield‐related parameters of rice plants[J]., 2011, 197(1): 40-49
[27] 周万海, 师尚礼, 寇江涛. 外源水杨酸对苜蓿幼苗盐胁迫的缓解效应[J]. 草业学报, 2012, 21(3): 171-176.
Zhou W H, Shi S L, Kou J T. Exogenous salicylic acid on alleviating salt stress in alfalfa seedlings[J]., 2012, 21(3): 171-176. (in Chinese with English abstract)
[28] 王宝增, 张一名, 张江丽. 水杨酸对盐胁迫下沙打旺幼苗生长的影响[J]. 草业学报, 2016, 25(8): 74-80.
Wang B Z, Zhang Y M, Zhang J L, Bi S Q, Kong H. Effects of salicylic acid on growth ofseedlings under salt stress[J]., 2016, 25(8): 74-80. (in Chinese with English abstract)
[29] Wang Y F, Hou Y X, Qiu J H, Wang H M, Wang S, Tang L Q, Tong X H, Zhang J. Abscisic acid promotes jasmonic acid biosynthesis via a 'SAPK10-bZIP72-AOC' pathway to synergistically inhibit seed germination in rice [J]., 2020, 228(4): 1-47
[30] 俞乐, 高彬, 李烨林, 谭伟健, 刘拥海. 外源ABA对GalLDH超表达转基因水稻种子萌发和生理指标的影响[J]. 种子, 2019, 38(12): 13-19
Yu L, Gao B, Li Y L, Tang W J, Liu Y H. Effects of exogenous ABA on seeds germination and physiological indicators of GalLDH-overexpressing rice[J]., 2019, 38(12): 13-19. (in Chinese with English abstract)
[31] Lee B T, Martin P, Bangerth F. Phytohormone levels in the florets of a single wheat spikelet during preanthesis development relationships to grain set[J]., 1988, 39(204): 933-937.
[32] 张上隆, 陈昆松, 叶庆富, 陈大明, 刘春荣. 柑桔授粉处理和单性结实子房(幼果)内源IAA、ABA和ZT含量的变化[J]. 园艺学报, 1994, 21(2): 117-123.
Zhang S L, Chen K S, Ye Q F. Changes of endogenous IAA, ABA and ZT in pollinated, non-pollinated and parthenocarpic ovary (fruitlet) of citrus[J].a, 1994, 21(2): 117-123. (in Chinese)
[33] Kato T, Sakurai N, Kuraishi S. The changes of endogenous abscisic acid in developing grains of two rice cultivars with different grain size[J]., 1993, 62(3): 456-461.
[34] 范晓荣, 沈其荣. ABA、IAA对旱作水稻叶片气孔的调节作用[J]. 中国农业科学, 2003, 36(12): 1450-1455.
Fang X R, Shen Q R. Effects of ABA and IAA on the behavior of stomata of rice crop cultivated in aerobic soil condition[J]., 2003, 36(12): 1450-1455. (in Chinese with English abstract)
[35] 段娜, 贾玉奎, 徐军, 陈海玲, 孙鹏. 植物内源激素研究进展[J]. 中国农学通报, 2015, 31(2): 159-165.
Duan N, Jia Y K, Xu J, Chen H L, Sun P. Research progress on plant endogenous hormones[J]., 2015, 31(2):159-165. (in Chinese with English abstract)
[36] 沙汉景, 胡文成, 贾琰, 王新鹏, 田雪飞, 于美芳, 赵宏伟. 外源水杨酸、脯氨酸和γ-氨基丁酸对盐胁迫下水稻产量的影响[J]. 作物学报, 2017, 43(11): 1677-1688.
Sha H J, Hu W C, Jia Y, Wang X P, Tian X F, Yu M F. Effect of exogenous salicylic acid, proline, and γ-aminobutyric acid on yield of rice under salt stress[J]., 2017, 43(11): 1677-1688. (in Chinese with English abstract)
[37] 杨军, 蔡哲, 刘丹, 胡犁月, 曲文波, 张崇华, 王尚明, 田俊. 高温下喷施水杨酸和磷酸二氢钾对中稻生理特征和产量的影响[J]. 应用生态学报, 2019, 30(12): 4202-4210.
Yang J, Cai Z, Liu D, Hu L Y, Qu W B, Zhang C H, Wang S M, Tian J. Effects of spraying salicylic acid and potassium dihydrogen phosphate on physiological characteristics and grain yield of single-season rice under high temperature condition[J]., 2019, 30(12): 4202-4210. (in Chinese with English abstract)
[38] 段俊, 田长恩, 梁承邺, 黄毓文, 刘鸿先. 水稻结实过程中穗不同部位谷粒中内源激素的动态变化[J]. 植物学报, 1999(1): 3-5.
Duan J, Tian C E, Liang C Y, Huang Y W, Liu H X. Dynamic changes of endogenous hormones in grains in different parts of the panicle during rice setting[J]., 1999(1): 3-5. (in Chinese)
[39] 萧浪涛, 王若仲, 丁君辉, 严钦泉. 内源激素与亚种间杂交稻籽粒灌浆的关系[J]. 湖南农业大学学报: 自然科学版, 2002, 28(4): 269-273.
Xiao L T, Wang R Z, Ding J H. Relationship between endogenous hormones and grain filling of inter- subspecies hybrid rice[J]., 2002, 28(4): 269-273. (in Chinese with English abstract)
[40] 陶龙兴, 王熹, 黄效林. 内源IAA 对杂交稻强、弱势粒灌浆增重的影响[J]. 中国水稻科学, 2003, 17(2): 56-62.
Tao L X, Wang X, Huang X L. Effects of endogenous IAA on grain filling of hybrid rice[J]., 2003, 17(2): 56-62. (in Chinese with English abstract)
Difference in Yield-related Traits of Grains in Various Parts of Panicle inRice and Its Correlation with Endogenous Hormones
ZHENG Xiaolong1, 2, ZHOU Jingqing3, TENG Ying4, ZHANG Linping2, SHAO Yafang2, HU Peisong1, 2, WEI Xiangjin2, *
(1College of Agriculture, Jiangxi Agricultural University, Nanchang 330045, China;2China National Rice Research Institute, Hangzhou 314001, China;3Zhejiang Environmental Monitoring Engineering Co. Ltd, Hangzhou 310015, China;4Plantation Technology Extension Center of Jintan District, Changzhou 213200, China;*Corresponding author, E-mail: weixiangjin@caas.cn)
【】The purpose of this study is to analyze the differences of grain yield in different parts of panicle given the levels of endogenous hormones in rice at grain filling stage. 【】 Six conventionalrice varieties from Zhejiang Province, including Jia58, Jia 67, Zhejing 99,Xiushui 121, Xiushui 134and Zhongjia 8, were used as materials. All the rice materials were grown in the same conditions. According to the internodes of the primary rachis branch on the main panicle, the rice paniclewas divided into the top, middle andbottom parts. The dynamic changes of grain salicylic acid (SA), abscisic acid (ABA), indole-3-acetic acid (IAA), fresh biomass, thousand-grain weight, head rice yield, length-width ratio, gel consistency, alkali spreading value during six rice development stages were measured. 【】 The thousand-grain weightofgrains from different panicle parts increased steadily during the filling stages(0-35 days) with the top parts being the heaviest and the bottom part being the lightest. The variation coefficient of grain biomass in different panicle parts ranged from 1.66% to 10.26% on the 35th day. Then the gaps between different panicle parts were narrowed, but the differences could be still observed. After maturity, thethousand-grain weight still followed the trend of the top (25.08-33.88g) > the middle (24.04-32.49g) > the bottom (23.46-31.78g).There was a slight difference in head rice yield and length-width ratio among different panicle parts. The correlation analysis among all the parameters showed that thousand-grain weight was positively correlated with length-width ratio (= 0.89,< 0.01), and negatively correlated with head rice yield (= –0.68,< 0.01).For the gel consistency and alkali spreading value, the differencesamong the six varieties was significantly greater than those among the three panicle parts. Significant differences were observed among the three endogenous phytohormones fromdifferent panicle parts. The variation coefficients of SA, ABA and IAA fromdifferent panicle parts were 0.59%–25.90%, 1.35%–66.25% and 0.16%–81.04%, respectively. At the earlyfilling stage, theSA content showed a tendency top > middle > bottom, while at the late filling stage, bottom > middle > top. 【】 It was found that the endogenous phytohormones and grain yield parameters of different panicle parts were significantly different. SA was positively correlated with ABA (=0.54,< 0.05), IAA (=0.59,< 0.01) and head rice yield (= 0.95,< 0.01), and was negatively correlated with thousand-grain weight (= –0.53,< 0.05) and length-width ratio(= –0.66,< 0.01).
grain position; grain biomass; endogenous phytohormone; difference; correlation
10.16819/j.1001-7216.2022.210112
2021-01-12;
2021-04-28。
国家农产品质量安全风险评估重大专项课题(GJFP2019043);中国农业科学院农业科技创新工程资助项目(CAASZDRW202011);中央级公益性科研院所基本科研业务费专项(CPSIBRF-CNRRI-202125)。
