阿拉善荒漠区两种跳鼠种群年龄结构及其季节动态特征*
2022-01-13甘红军赵云华赵永泉苏巴特尔苏佳楼付和平
甘红军,张 蓉,马 久,赵云华,赵永泉,李 伟,苏巴特尔,苏佳楼,袁 帅,付和平,*
(1.阿拉善盟草原工作站,内蒙古 巴彦浩特 730000;2.内蒙古农业大学草原与资源环境学院/内蒙古农业大学啮齿动物研究中心/草地资源教育部重点实验室/农业农村部饲草栽培、加工与高效利用重点实验室/内蒙古自治区草地管理与利用重点实验室,呼和浩特 010018;3.乌兰察布市察右后旗土牧尔台镇政府,内蒙古 土牧尔台 012406;4.鄂尔多斯市杭锦旗草原工作站,内蒙古 锡尼 017400;5.呼伦贝尔市林业和草原事业发展中心,内蒙古海拉尔 021008;6.锡林郭勒盟草原工作站,内蒙古 锡林浩特 026000)
草地啮齿动物在生态系统中具有不可替代的功能和作用,但其种群数量一旦暴发就会形成“鼠害”,因此其种群数量动态一直以来备受关注。而与其种群动态紧密联系的生物学特征之一是种群年龄结构,它一般预示着动物种群的消长,通过对种群年龄结构的分析研究,可以对动物种群的发展趋势进行较为准确的判断。国内外的有关研究表明,啮齿动物种群数量的消长与年龄结构密切相关,而且具有明显的区域特点,明显受到当地生境的生物因素和非生物因素影响[1~ 6]。本研究于2014~2016年在阿拉善荒漠区选择开垦、轮牧和禁牧三种不同典型生境,对具有冬眠习性的五趾跳鼠(Allactagasibirica)和三趾跳鼠(Dipussagitta)进行野外调查,并对两种跳鼠在不同生境的年龄结构及其季节变化特征进行分析,以期为荒漠区跳鼠类啮齿动物种群生态学和鼠害控制提供基础资料。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于内蒙古自治区阿拉善盟阿左旗南部的典型荒漠区,地处腾格里沙漠东缘,地理坐标为104°10′~105°30′E、37°24′~38°25′N,海拔1399m左右。气候为典型的高原大陆性气候,冬季严寒、干燥,夏季酷热,昼夜温差大,极端最低气温为-36℃,最高气温42℃。该地区的草地类型是典型的温性荒漠,植被稀疏,结构单调,覆盖度1%~25%。植被以旱生、超旱生和盐生的灌木、半灌木、小灌木为主;降水量为75~215mm,主要集中在7~9月份;土壤类型为灰漠土和灰棕土[7]。
1.2 研究方法
本研究于2014~2016年每年的4月、7月和10月进行。在阿拉善荒漠区依据草地利用方式,选择开垦区、轮牧区和禁牧区三种不同生境,采用夹日法对五趾跳鼠和三趾跳鼠进行调查,每种生境设置3个重复调查样地,每个样地放置300夹,共计8100夹。夹距5m,行距50m,样方间隔大于1km,每个样方面积为5hm2。以花生米作诱饵,持续布放24h。记录所捕获两种跳鼠标本的体重、体长、尾长、后足长和耳长,以及性别、繁殖状况及捕获时间和地点。三年中共记录五趾跳鼠标本135只,其中雌鼠43只,雄鼠92只;三趾跳鼠样本434只,其中雌鼠194只,雄鼠240只。
2 数据处理
利用SPSS 22.0软件进行差异性T-检验(显著水平为P<0.05,极显著水平为P<0.01),系统聚类分析划分年龄组,性别比例为雌性与雄性数量比值。
3 结果与分析
3.1 荒漠区两种跳鼠年龄组划分
本研究以两种跳鼠活体体重为划分年龄组的主要指标,以体长、尾长、耳长、后足长以及繁殖状况等作为辅助指标以提高准确性。考虑怀孕雌鼠对体重的影响,分别对两种跳鼠孕鼠和未孕雌鼠的体重指标进行独立样本T检验。结果显示,孕鼠与未孕雌鼠体重差异不显著(P>0.05,见表1),表明是否怀孕对两种跳鼠体重影响差异不显著,进行年龄组划分时可以忽略怀孕因素的干扰。因此,以体重作为主要指标,应用系统聚类分析将五趾跳鼠年龄组划分为3组:I龄组62~80g(亚成体),II龄组80~110g(成体),III龄组>110g(老年体)[7~8];应用系统聚类分析将三趾跳鼠年龄组也划分为3组:I龄组30~53g(幼体),II龄组55~78g(亚成体),III龄组79~98g(成体)[7]。
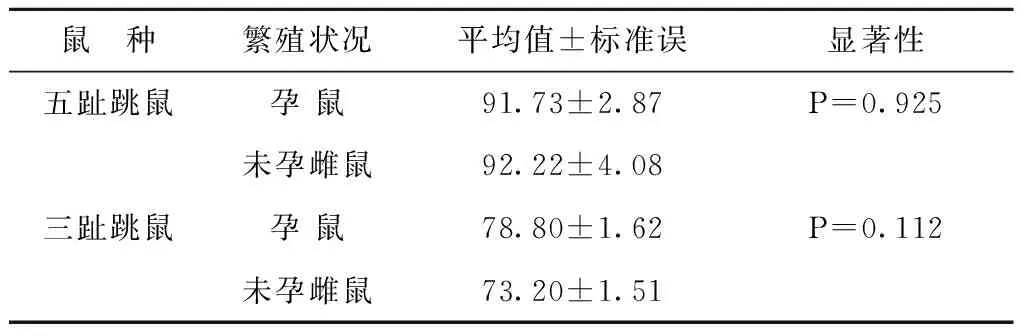
表1 两种跳鼠孕鼠与未孕雌鼠体重差异性T检验
3.2 不同生境两种跳鼠年龄结构
五趾跳鼠在开垦、轮牧和禁牧三种不同生境中的年龄结构如图1。由图1-A可知,五趾跳鼠种群在三种生境中均以II龄组数量最高。开垦生境中只有II龄组和III龄组2个年龄组,且数量差异不显著。由于无I龄组个体,开垦生境五趾跳鼠种群为下降型种群。轮牧和禁牧生境中,II龄组与其他2个年龄组数量差异达到显著水平(F=1.56,P<0.05)和极显著水平(F=2.68,P<0.01),而且I龄组所占比例明显高于III龄组,五趾跳鼠种群为增长型种群。开垦生境中3个年龄组雌雄性比分别为1∶0.8、1∶1.8和1∶1.4;轮牧生境中3个年龄组性比分别为1∶5.6、1∶12和1∶1.8;禁牧生境中3个年龄组性比分别为1∶3.3、1∶2.16和2∶1.4。
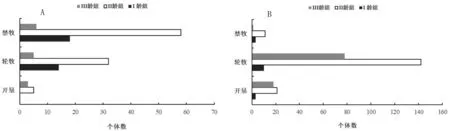
图1 不同生境五趾跳鼠(A)和三趾跳鼠(B)年龄结构
三趾跳鼠在开垦、轮牧和禁牧三种不同生境中的年龄结构如图1。由图1-B可知,三趾跳鼠种群在三种生境中同样均以II龄组数量最高,开垦生境中II龄组和III龄组数量大于I龄组,而且达到了差异显著水平(F=1.14,P<0.05;F=1.24,P<0.05),I龄组占比极小,因此三趾跳鼠种群为下降型种群。在轮牧生境中,II龄组和III龄组数量均极显著大于I龄组(F=2.63,P<0.01;F=2.47,P<0.01),三趾跳鼠种群为下降型种群。在禁牧生境中II龄组数量大于I龄组和III龄组,II龄组数量与III龄组有显著(F=1.32,P<0.05)差异,三趾跳鼠种群有增长趋势。在开垦生境中3个年龄组雌雄性比分别为1∶0.8、1∶2.7和1∶1.4;在轮牧生境中3个年龄组性比分别为1∶0.32、1∶1.7和1∶1.32;在禁牧生境中3个年龄组性比分别为1∶0.8、1∶2.22和1∶1.4。
3.3 不同生境两种跳鼠年龄结构季节动态
3.3.1五趾跳鼠年龄结构的季节动态
在4月、7月和10月份进行的研究分别对应于春季、夏季和秋季3个季节,以便分析开垦区(图2-A)、轮牧区(图2-B)和禁牧区(图2-C)三种不同生境五趾跳鼠年龄结构的季节动态。由图2-A可知,在开垦区五趾跳鼠在3个季节中II龄组种群数量明显高于III龄组,而且数量差异并不显著,三年中不同季节均未捕获到I龄组个体,说明五趾跳鼠不在开垦生境中繁殖;3个季节开垦生境中只有II龄组和III龄组个体,表明五趾跳鼠种群数量趋于下降型。由图2-B可知,在轮牧生境3个季节中3个年龄组均有捕获,而I龄组只出现在夏季(7月)五趾跳鼠的繁殖高峰期。从全年来看,II龄组数量明显高于其他年龄组,与I龄组差异显著(F=2.46,P<0.05),与III龄组数量差异极显著(F=3.98,P<0.01);I龄组数量显著高于III龄组(F=1.45,P<0.05)。由图2-C可知,在禁牧生境3个季节中3个年龄组同样均有捕获,而I龄组同样只出现在夏季(7月)五趾跳鼠的繁殖高峰期;从全年来看,II龄组数量明显高于其他年龄组,与I龄组差异显著(F=1.96,P<0.05),与III龄组数量差异极显著(F=2.87,P<0.01);I龄组数量显著高于III龄组(F=1.82,P<0.05)。
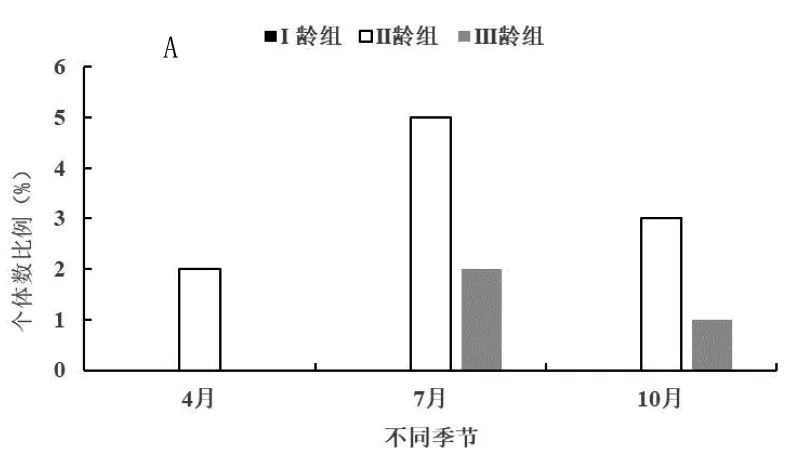
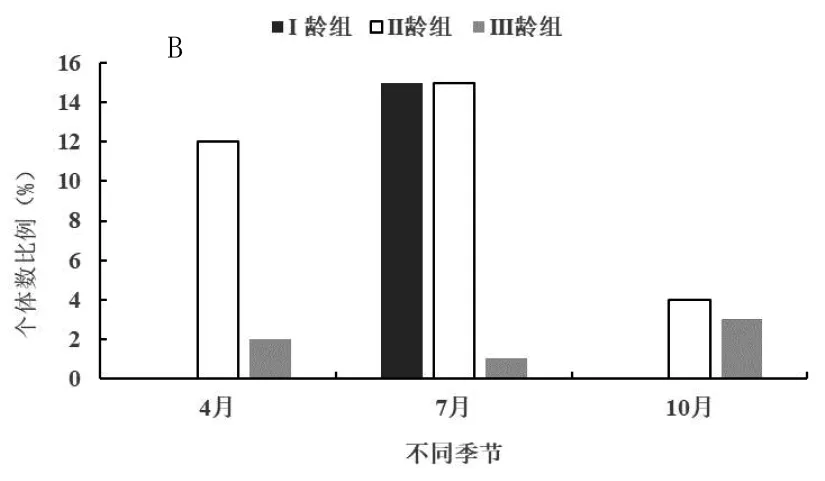
A为开垦区;B为轮牧区;C为禁牧区;下图同图2 不同生境五趾跳鼠年龄结构季节动态
由图2-B和2-C可知,在轮牧和禁牧生境,由于五趾跳鼠种群在夏季I龄组数量显著高于III龄组,因此夏季为增长型种群,春季和秋季为下降型种群。
3.3.2三趾跳鼠年龄结构的季节动态
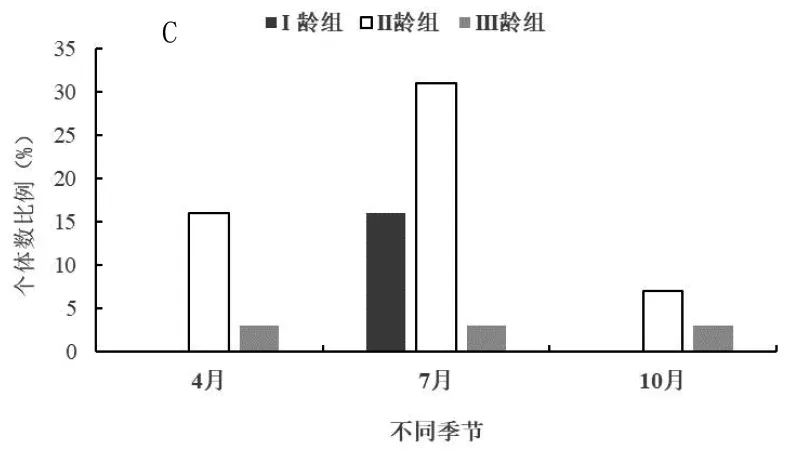
三趾跳鼠在开垦、轮牧和禁牧三种生境中不同季节(4月为春季、7月为夏季、10月为秋季)年龄结构动态如图3。由图3-A可知,开垦生境中3个季节三趾跳鼠II龄组数量均最高,在春季3个年龄组均有捕获,I龄组少量出现说明三趾跳鼠在开垦区进行繁殖,而且I龄组数量显著小于II龄组和III龄组(F=3.11,P<0.01;F=2.52,P<0.05);夏季以II龄组和III龄组为主,秋季只有II龄组个体,III龄组个体已经进入冬眠期,因此在开垦区3个季节三趾跳鼠种群均为下降型种群。由图3-B可知,轮牧生境中3个季节同样II龄组数量最高,整体来看II龄组数量显著高于I龄组和III龄组(F=3.41,P<0.01;F=2.24,P<0.05),III龄组数量也显著高于I龄组(F=2.95,P<0.01),因此在3个季节三趾跳鼠种群均为下降型种群;夏季I龄组个体出现较多,说明在轮牧生境中三趾跳鼠主要在夏季进行繁殖,而在秋季进入冬眠较开垦生境中的个体要晚。由图3-C可知,禁牧生境中3个季节同样II龄组数量最高,而且与I龄组和III龄组具有极显著的差异(F=3.13,P<0.01;F=3.59,P<0.01);春季出现I龄组和II龄组,说明三趾跳鼠在禁牧生境中进行繁殖,而夏季只有II龄组和III龄组个体,秋季3个年龄组均没有捕获,说明在禁牧生境中三趾跳鼠较开垦和轮牧生境提前进入冬眠;由于春季I龄组数量明显高于III龄组,因此三趾跳鼠种群为增长型种群,夏季为下降型种群。
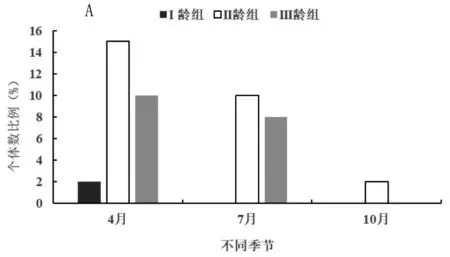
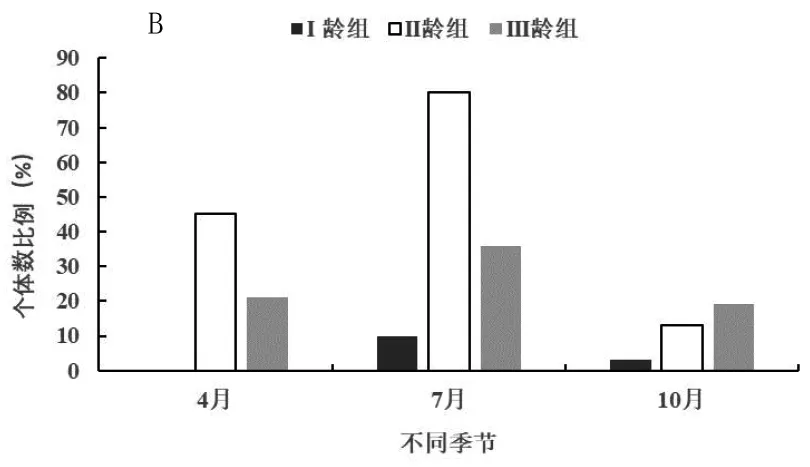
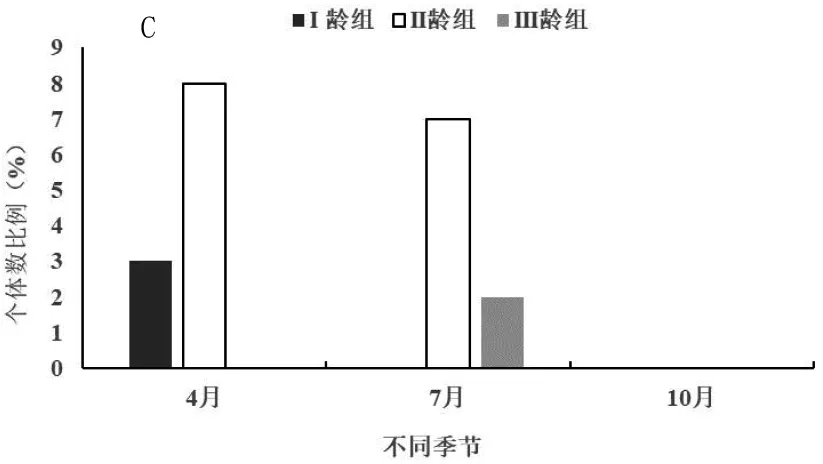
图3 不同生境三趾跳鼠年龄结构季节动态
4 讨论与结论
在草地生态系统中,生物灾害的防控一直以来都是草地生态与管理的难点和重点[9],特别是草地鼠害的治理,需要长期对不同种类的啮齿动物种群数量动态特征进行研究才能提出较为合理的防控措施。而某种啮齿动物种群数量消长的精确判断,离不开对其年龄结构的分析。一般来说,啮齿动物年龄结构与其繁殖开始的时间、每年繁殖次数以及季节变化、环境因子干扰等有密切关系。有关研究表明,棕色田鼠(Lasiopodomysmandarinus)种群在春季以III龄组、IV龄组占比较高,初夏I龄组、II龄组个体数明显上升,冬季以II龄组、III龄组为主[6,10]。红尾沙鼠(Merioneslibycus)在春季时主要为成体,夏季时幼年数量开始上升,亚成体和成体仍然占比较高,秋季亚成体数量下降,成体和老年体占比增加[11]。本研究中五趾跳鼠和三趾跳鼠的年龄结构随季节的变化与上述两种均有所不同,五趾跳鼠在春季时以成体为主,且存在少量老年个体;夏季时以亚成体和成体为主,老年个体占比较少;进入秋季则以成体和老年体为主,无亚成体[7,8],此时也是五趾跳鼠进入冬眠的时期。三趾跳鼠在春季时以亚成体和成体为主,幼体占比较少;夏季幼体数量虽然上升,但仍然以亚成体和成体为主;秋季亚成体和成体数量明显下降,这是由于其进入冬眠期。根据我们连续多年的野外调查,三趾跳鼠在阿拉善荒漠区一般在10月上旬开始进入冬眠期,到10月中旬全部冬眠。
综上所述,在开垦、轮牧和禁牧三种不同类型生境中,五趾跳鼠和三趾跳鼠均以II龄组数量为最高。在开垦生境中,五趾跳鼠种群为下降型种群;在轮牧和禁牧生境中,夏季为增长型种群,春季和秋季为下降型种群。三趾跳鼠种群在开垦和轮牧生境中,春季、夏季与秋季均为下降型种群;在禁牧生境中,春季为增长型种群,夏季为下降型种群。
