氮素添加对高寒草甸植物群落多样性和土壤生态化学计量特征的影响
2022-01-08向雪梅德科加林伟山冯廷旭魏希杰徐成体钱诗祎
向雪梅, 德科加, 林伟山, 冯廷旭, 魏希杰, 王 伟, 徐成体, 钱诗祎
(青海大学畜牧兽医科学院, 青海省畜牧兽医科学院, 青海 西宁 810016)
三江源区地处青藏高原腹地,是我国重要的水源涵养和生态功能保护区,有“中华水塔”的美誉,其主要的草地类型为高寒草甸[1-2]。高寒草甸生态系统的健康发展对维持三江源区的草-畜平衡有重要的作用[3]。
近几年来,由于矿物燃料的大量开采及化肥在农业中的大量使用,导致大气氮沉降日益增加[4]。而氮素增加会直接或间接的影响高寒草甸生态系统的稳定性。已有研究表明氮素添加会促进植物不定根数量增多,使土壤质地疏松,土壤容重降低,从而明显改善土壤结构,良好的土壤结构有利于土壤养分的释放及腐殖质的积累[5-6]。土壤是植物生长的营养库,不同植物生长所需的养分元素种类和数量是不同的[7]。当土壤养分发生改变时植物群落内组分会依据自生生物学特性最大化汲取自身生长需要的资源,导致物种种间竞争加剧,植物群落和结构的改变最终影响生态系统功能[8-10]。
土壤碳(Carbon,C)、氮(Nitrogen,N)、磷(Phosphorus,P)不仅能反映土壤肥力状况,其化学计量比也是反映土壤有机质构成和土壤质量状况及养分供给能力的重要指标[11]。生态化学计量学是研究生态系统多种化学元素平衡关系的一个新兴生态学研究领域,研究土壤C∶N∶P的生态化学计量学特征对揭示土壤养分循环和平衡机制有着重要的价值[12-13]。目前,王建林等人研究了青藏高原生态系统土壤C/N的分布特征发现,不同草地型和不同自然地带土壤C/N差异显著,而表土层与底土层土壤C/N差异也显著[14];Yue等关于全球陆地生态系统C∶N∶P对N沉降的研究发现,N添加与土壤C/N呈负相关关系,对土壤N/P没有显著影响[15];张骥等人在青藏高原高寒草甸中研究表明氮磷添加显著降低了土壤C/N和C/P[16]。
前人研究多集中于氮素添加对物种多样性、土壤养分及其化学计量特征的影响,而关于氮素添加后对生态系统植物多样性与土壤生态化学计量特征之间的相关性研究鲜见报道。已有研究表明了解植物群落特征与土壤因子间的关系对生态系统的恢复具有重要意义[17]。因此探讨氮素添加下高寒草甸生态系统中植物多样性与土壤生态化学计量特征间相关性很有必要。
本文以三江源区高寒草甸为研究对象,研究不同水平氮素添加下草场群落特征、物种多样性及土壤特征的变化,分析引起植被群落组成发生改变的关键土壤因子,为全球变化下高寒草甸植物群落与土壤特性变化过程间的内在联系提供理论依据。
1 材料与方法
1.1 样地自然概况
研究地点位于青海大学三江源生态系统教育部野外观测站(北纬33° 24′30′′,东经 97° 18′00′′),海拔高度为4 270 m,气候为典型的高原大陆性气候,年平均气温为—10.3℃~4.6℃,年均降水量为614.1 mm,年度降水主要分布在6—9月份,约占全年降水的50%。该草地类型为矮嵩草(KobresiahumilisClarke)+杂类草草甸,草场主要优势牧草是矮嵩草、异针茅(StipaalienaKeng) 等。伴生植物包括羊茅(FestucaovinaL.)、垂穗披碱草(ElymusnutansKeng) 等。不食杂类草和毒草有黄花棘豆(OxytropisochrocephalaBunge.)、青藏龙胆(GentianafuttereriDiels et Gilg)、狼毒(S.chamaejasmeL.)等。土壤pH值为6.92,有机质含量为2.36%,速效N含量为140 mg·L-1,速效P含量为7.0 mg·L-1,速效K含量为76.5 mg·L-1。
1.2 试验设计
2020年6月25日,在称多县珍秦镇选择地势平坦、植被均匀有代表性的围栏地段草地作为试验样地,试验样地面积为462 m2。以三江源区高寒草甸前期研究成果为依据[18-19],氮素水平设为0(N0),15 g N·m-2(N1),30 g N·m-2(N2),45 g N·m-2(N3),60 g N·m-2(N4),其中不施任何肥料为对照组(N0),共5个处理。随机区组设计,每处理3次重复,小区面积为20 m2(4 m×5 m)。施肥种类为尿素(CON2H4),采用喷洒方式,每小区均匀喷洒,N0处理小区喷洒等量水。
1.3 植被特征调查
2020年9月,在每小区重复取样3次,取样面积为50 cm×50 cm。调查样方内各个植物物种的高度、盖度及地上生物量。其中,植物物种的高度为样方内每个物种随机测定5株植物自然高度;植物物种的盖度用目测法测定[20];植物物种地上鲜重直接齐地剪下测定;测定后,将样方内植物种按不同经济类群(禾本科、莎草科和杂类草)分别装入布袋,带回实验室,置于65℃烘箱中烘干称重。
基于样方植物种高度、盖度和鲜重调查数据,按公式(1)~(4),分别计算群落植物经济类群重要值、植物群落Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)、Margalef(S)指数。

(1)
Shannon-Wiener多样性指数(H)=-∑Piln(Pi)
(2)
Margalef物种丰富度指数(S)=样方内出现的物种数
(3)
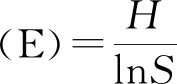
(4)
式中:Pi为群落中的种i的相对重要值,S为种i所在样方中的数目[22]。
1.4 土壤采集与养分分析

1.5 数据处理
采用SPSS 22.0软件对数据进行单因素方差分析(One-way ANOVA,LSD),数据显著性分析采用Duncan法,采用CANOCO5软件对植物群落多样性指数和土壤因子的对应关系进行冗余分析(RDA),采用Origin 2019软件制图,图表中数据为“平均值±标准误”。
2 结果与分析
2.1 氮素添加对草地经济类群重要值的影响
随着氮素水平增加,禾本科重要值呈现逐渐增加的趋势,与N0相比较,N3,N4处理分别增加了7.45%,9.63%(P<0.05);莎草科重要值随氮素水平增加呈先增加后降低的趋势,与N0相比较,N4处理降低了32.08%(P<0.05);杂类草重要值随氮素水平增加呈逐渐下降的趋势,N4处理显著低于N0处理(P<0.05)。
2.2 氮添加对高寒草甸植物群落物种多样性的影响
随着氮素添加水平增加,Shannon-Wiener指数呈下降趋势,与N0相比较,N4处理下降低了2.81%(P<0.05);氮添加处理的Pielou均匀度指数与对照无显著差异;Margalef物种丰富度指数随氮素水平增加呈下降趋势,与N0相比,N4处理下降低了32.43%(P<0.05)。

表1 不同水平氮添加下群落物种多样性的比较
2.3 不同水平氮添加对高寒草甸土壤因子特征的影响
土壤全氮含量在0~15 cm土层中随氮素水平增加呈降低的趋势,各氮添加处理组与N0差异不显著;在15~30 cm土层中全氮含量呈先增加后降低的趋势,在N2处理下显著高于N0(P<0.05)。
土壤铵态氮含量在0~15 cm和15~30 cm土层都随氮素水平增加而增加,在0~15 cm土层,与N0相比,各处理下土壤铵态氮含量分别增加了15.46%,14.54%,19.00%,33.70%(P<0.05),15~30 cm土层中,只有N4处理的土壤铵态氮含量显著高于对照(P<0.05)。
土壤硝态氮含量在0~15 cm土层中随氮素水平增加呈先降低后增加再降低的趋势,在N3处理下最高,与N0相比,增加了110.32%(P<0.05),在15~30 cm土层中,N3,N4处理下显著高于N0处理,分别高143.66%,428.17%(P<0.05)。
土壤有机碳含量在0~15 cm的土层中随氮素水平增加呈先增加后降低趋势,在N1,N2,N4处理下显著高于N0处理,分别高26.46%,23.10%,25.09%(P<0.05),在15~30 cm土层中不同处理的土壤有机碳含量差异不显著。
土壤全磷含量在不同土层中随氮素添加水平升高均呈先增加后降低的趋势。在0~15 cm土层中,N3处理下最高,与N0相比增加了202.78%,在15~30 cm土层中N2处理下最高,增加了343.73 g·kg-1(P<0.05)。
样地中土壤全氮、铵态氮、硝态氮、有机碳60%以上分布于0~15 cm土层,而土壤全磷则大多分布于15~30 cm土层。

图2 氮素添加对土壤养分的影响
2.4 氮素添加对土壤C∶N∶P化学计量比的影响
氮素添加引起土壤C∶N∶P生态化学计量特征不同程度的变化,0~15 cm土层土壤C/N随氮素水平升高而增加,与N0相比,N1,N4处理C/N增加了27.72%,38.04%(P<0.05);15~30 cm土层土壤C/N随氮素添加呈先增加后降低的趋势,与N0相比,N2,N3,N4处理C/N显著降低,分别降低了43.18%,35.12%,25.63%(P<0.05)。土壤C/P在 0~15 cm土层随氮素水平升高而降低,与N0相比,各处理分别降低46.97%,55.18%,66.01%,49.79%(P<0.05);在15~30 cm土层土壤C/P也随氮素水平升高而降低,与N0相比,各处理分别降低了65.19%,71.31%,74.49%,66.27%(P<0.05)。在0~15 cm土层,与N0相比,N1,N2处理下土壤N/P增加了44.39%,47.80%(P<0.05),在15~30 cm土层中,与N0相比,土壤N/P在N2,N3,N4处理下分别增加了119.89%,72.33%,81.34%(P<0.05)。

图3 氮添加对土壤C∶N∶P生态化学计量特征的影响
2.5 植物群落特征与土壤因子的关系
由表2可知,第一轴和第二轴共解释87.88%的植物群落特征变化和99.99%的群落结构特征与土壤因子关系,结果表明土壤因子对植物经济类群的分布具有显著影响(P=0.018<0.05)。

表2 高寒草甸植物群落特征与土壤因子的RDA二维排序结果
根据图4中箭头的长度可以看出,对植物群落多样性影响最大的土壤因子分别是土壤有机碳、铵态氮含量。根据实线与虚线的夹角可以看出,Shannon-Wiener多样性指数与土壤N/P,C/P、全氮含量呈正相关关系,与土壤有机碳含量、C/N、全磷含量、硝态氮含量、铵态氮含量呈负相关关系。Margalef物种丰富度指数与土壤有机碳含量、全氮含量、C/P,N/P呈正相关关系,与C/N、全磷含量、硝态氮含量、铵态氮含量负相关。Pielou均匀度指数与土壤C/N、全磷含量、硝态氮含量、铵态氮含量呈正相关关系,与有机碳含量、全氮含量、C/P,N/P呈负相关关系。

图4 植物群落多样性指数与土壤因子关系的RDA排序图
3 讨论
3.1 氮素添加对植物群落组成及物种多样性的影响
氮素添加通过增加草地可利用竞争性资源的方式影响草地植物种群间的竞争,从而影响草地的物种组成、草地生产力和草地的稳定性[24-25]。本研究发现在同一氮素水平下莎草科植物重要值高于禾本科及杂类草,说明莎草科植物在群落中占据主导地位,影响着该植物群落生态小环境[26]。随着氮素添加水平增加,种群结构和主要的牧草种类发生了变化,在添加60 g·m-2纯氮处理禾本科重要值达到最大值,类杂草重要值则最小,同时在该氮素添加量下植物多样性明显降低。这是因为纯氮量添加到30~45 g·m-2时,禾本科植物、莎草科、杂类草植物等的盖度、高度差异不明显,高寒草甸生境变化很小,植物多样性变化不大。在添加60 g·m-2纯氮时,植物多样性呈降低的趋势,主要因为氮素添加下禾本科植株密度增加,个体数加大,而杂类草和莎草科植株相对减少,不同功能群植物对某些营养物质利用效率的高低存在差异[27]。并且氮素添加后植物从地下部分对养分的竞争转变为地上部分对光照的竞争,而矮小物种在光竞争中处于劣势,使得群落中的一些物种被淘汰[28],从而导致物种多样性降低。另外,群落多样性的变化除了植被本身变化,还受到气候条件等其他非生物条件的影响[29]。
3.2 不同水平氮素添加对土壤养分及其化学计量比的影响
土壤养分状况是土壤物理、化学、生物等因素耦合的结果,为植物的生长提供营养元素,并有效地调节植物的功能属性[30]。
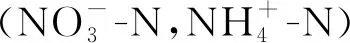
土壤有机碳是土壤供氮的重要物质基础[36]。本研究中,氮素添加能增加土壤有机碳含量。这是因为外源氮素添加在一定程度上缓解了土壤氮素缺乏对植物生长的抑制,促进植物生长和光合作用,增加了植物对碳的同化,从而增加以凋落物的形式进入土壤的有机质的量,归还到土壤表层中的养分也随之增加[37]。总体来看土壤有机碳随着氮素添加水平升高呈先增加后降低的趋势,并在添加45 g·m-2纯氮时,土壤有机碳含量呈下降趋势,可能是土壤微生物在这个处理下活性较高,消耗有机质所致[38]。
土壤磷有比较固定的来源,主要是岩石风化,只有很小有部分是来自于有机质降解[39]。全磷含量随着氮素添加水平的升高先增加后降低,这说明适宜的氮素添加能增加土壤全磷含量,而过高的氮素会降低土壤的全磷含量,因为土壤全磷主要源于成土母质,且由于氮素添加使植物群落组成发生变化,而导致土壤全磷含量分布上的差异[40]。土壤全磷受到气候、母质等多种因素的影响,加之施氮年限较短,并不能得出一致的结论。
此外,施氮对不同土层的养分含量影响也不尽相同。在本试验中研究发现土壤全磷、硝态氮在不同土层均表现出显著差异,但土壤有机碳含量在0~15 cm土层中变化明显,而在15~30 cm土层中总氮和铵态氮含量变化明显。这表明施氮后土壤中元素变化并不相同,这主要是由土壤养分空间异质性造成的[41]。氮素添加后土壤养分大都会向表层聚集,这与韩发等人的研究一致[42]。主要是因为植被以浅根系植物为主,故表层的凋落物及根系较多,养分含量也较高。
土壤C,N,P化学计量比变化也是土壤-植被生态系统养分循环的重要环节,有利于判定土壤限制性元素及养分调控因素[43]。土壤C/P是衡量磷有效性的指标之一,C/P越低则磷的有效性越高[44]。本研究发现随着氮素添加土壤C/P呈降低趋势。因为氮素添加会改变土壤中氮素和磷素的相对平衡关系,促进氮素与有机碳的结合,加速有机物分解,从而降低了土壤C/P[45]。
土壤C/N是土壤氮素矿化能力的标志[46]。在15~30 cm土层中随氮素添加土壤C/N呈下降趋势,这与陈磊等对黄土高原油松林土壤短期氮添加的研究结果一致[47]。土壤氮素含量增加会加速土壤微生物对有机质的分解矿化,从而导致土壤C/N降低。不同土层中土壤C/N变化不同,这更大程度上反映出土壤深层剖面腐殖质的年代和分层化[48]。
土壤N/P反映土壤肥沃程度,直接影响土壤养分的生物利用有效性[49]。低的土壤N/P意味着生态系统主要受氮限制[50]。在本研究中发现氮添加会提高土壤N/P,这与刘红梅等人的研究一致[51]。表明氮添加减缓了草原植物氮限制,但是磷限制性增强[52]。与土壤C/N相比,随着氮素添加水平增加,土壤C/P和N/P的变化幅度较大,这表明土壤氮和碳之间有较为紧密的耦合关系[53]
3.3 植物群落多样性与土壤C∶N∶P生态化学计量特征及其他关键因子的关系
草地植物多样性与土壤环境因子间的相关性是草地生态系统维持相对稳定的基础[54],因此探讨植物多样性与土壤营养元素定量关系具有重要意义。有研究表明物种多样性指数与土壤养分及其化学计量比间存在显著相关性[55]。本研究发现高寒草甸植物多样性与土壤养分间的相关性随着氮素添加而发生改变。氮素添加下Shannon-Wiener多样性指数与全氮含量呈正相关关系,物种丰富度会随着土壤有机质和全氮含量的增加而增加,这与孙小丽等人在荒漠草原的研究一致[56]。氮素添加下均匀度指数与土壤硝态氮、铵态氮、全氮含量呈正相关关系。土壤因子是影响研究区植物种类变化的重要因素,当土壤养分含量之间存在差异时植物多样性会发生变化,这与Kristina得出的结论吻合[57]。
此外,土壤C∶N∶P平衡关系在维持植物群落结构方面发挥着重要的作用[58]。有研究表明植物群落多样性随土壤C/N和N/P增加而增加[59]。而本研究发现随着氮素添加下土壤C/P和N/P与物种多样性呈正相关关系,与土壤C/N呈负相关关系,表明氮素添加调节了土壤元素化学计量平衡关系,从而影响了植物群落多样性。冗余分析结果表明,在不同水平氮素添加下对植物群落多样性影响最大的土壤因子是土壤有机碳、铵态氮含量。不同水平氮素添加下植物多样性与土壤养分并不是简单的线性关系,还需进一步研究。
4 结论
在高寒草甸中,氮素添加能提高土壤有机碳、全氮、铵态氮、硝态氮、全磷含量,影响土壤生态化学计量比。当添加量为60 gN·m-2时禾本科重要值最高,杂类草重要值最低,植物多样性指数显著降低。氮素添加能影响植物多样性与土壤养分及化学计量比相关性,土壤有机碳和铵态氮含量是影响植物多样性的关键因子。
