河南省臭椿炭疽病病原鉴定
2021-12-29王树和张佳正何金鹤
王树和,张佳正,何金鹤
(河南科技大学园艺与植物保护学院,河南 洛阳 471000)
臭椿(Ailanthus altissima(Mill.)Swingle)为苦木科(Simaroubaceae)臭椿属落叶乔木,原产于中国北部及中部,全国各省区几乎都有分布[1]。臭椿树干高大挺拔,树皮光滑,树冠如伞状,极具观赏价值;而且该树种耐盐碱和干旱、抗烟尘和病虫害,是一种抗性极强的优良绿化树种[2-3]。因此,臭椿被广泛用于城市园林建设、山区植树造林和工矿区绿化,单独种植或与其它树种一起混种均可。除用于观赏绿化外,臭椿还是药用植物,其树皮、根皮及果实均可入药,具有重要的经济价值[4-5]。
2018 年9 月笔者在河南省洛阳市嵩县天池山国家森林公园进行病害调查,发现臭椿炭疽病严重发生,一些植株病叶率达到70%以上。该病主要危害叶片,病斑褐色呈圆形或近圆形,边缘颜色较深,病斑中央坏死组织常脱落形成穿孔,后期造成臭椿大量落叶,严重影响树木生长和景观价值。王教敏[6]于2009 报道了青岛地区发生的臭椿炭疽病,基于形态学和rDNA-ITS 序列,鉴定分离菌株SQD-107 为胶孢炭疽菌(Colletotrichum gloeosporioidesPenz.)。本次调查观察到的臭椿炭疽病与王教敏[6]描述的症状存在着较为明显的差异,发病后期多数病斑形成穿孔。本研究拟采用形态学与多基因系统发育分析相结合的方法,对该地区臭椿炭疽病病原菌鉴定,并经柯赫氏法则验证,以期为该病害防治策略的制定提供参考。
1 材料与方法
1.1 材料
臭椿炭疽病病叶采集于河南省洛阳市嵩县天池山国家森林公园。致病性测定的臭椿健康叶片采集于河南科技大学校园。
1.2 方法
1.2.1 病原菌分离纯化 参照Chen 等[7]病原组织分离法分离病原菌,分离的菌株单孢纯化后,保存于PDA 斜面上,置4℃冰箱中保存备用。
1.2.2 致病性测定 分离菌株CH-1 和CH-3 用于接种试验,参照Torres-Calzada 等[8]的方法进行有伤和无伤接种(图1),将采集的臭椿健康叶片用0.5% NaClO 表面消毒,无菌水冲洗3 次,叶面水分晾干后进行接种。有伤接种时用无菌接种针刺伤叶片,移液枪吸取供试菌株分生孢子悬浮液20 μL(浓度为1×106个·mL−1)滴在叶片刺伤部位,对照处理接种20 μL 无菌水;无伤接种对健叶未进行刺伤,其余操作同有伤接种。每个处理接种10 片叶子,试验重复2 次。接种后的叶片放置于加有湿润滤纸的保鲜盒内,在叶柄处包裹脱脂棉并滴加无菌水,将装有叶片保鲜盒置于培养箱内,培养条件为:温度25℃,12 h 光暗交替。逐日观察叶片的发病情况,接种叶片发病后再进行分离培养,完成柯赫氏法则的验证。
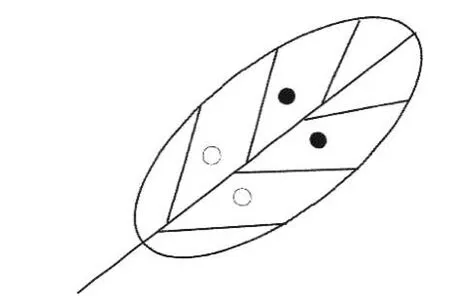
图1 叶片接种示意图Fig.1 Schematic diagram of the protocol used to leaf inoculation
1.3 病原菌的形态观察
参照Yan 等[9]描述的方法进行形态学鉴定。在光学显微镜(Leica DM2500,Germany)下观察分生孢子梗、分生孢子、子囊、子囊孢子等微观形态,并测量分生孢子和子囊孢子(n=50)的大小。依照Yang 等[10]描述的方法诱导分生孢子附着胞的产生,观察记录附着胞形态特征并测量其大小(n=30)。
1.4 分子生物学鉴定
采用改良CTAB 法[11]提取菌株CH-1 和CH-3 基因组DNA。使用rDNA-ITS 通用引物ITS5/ITS4[12]、肌动蛋白(actin,ACT)引物ACT-512F/ACT-783R[13]、β-微管蛋白(beta-tubulin 2,TUB2)引物TUBT1/TUB2b[14-15]、甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)引物GDF1/GDR1[16]和几丁质合成酶(chitin synthase,CHS-1)引 物CHS-79F/CHS-345R[13]对病原菌基因组DNA 进行扩增。
PCR 扩增得到的目的片段通过1%的琼脂糖凝胶电泳检测后委托北京擎科生物技术公司进行测序。测序结果在NCBI(http://www.ncbi.nlm.gov)数据库中进行BLAST 比对,根据比对结果下载参比序列(表1),使用PhyloSuite 软件中MAFFA 插件对建树序列进行多重比对[17],必要时进行手工校正。比对后的基因序列在PhyloSuite 软件中按照ACT、TUB2、CHS-1、GAPDH和ITS的顺序串联整合成一个多基因数据集[17],基因序列合并的数据集在MEGA X 软件中采用最大似然法(maximum likelihood,ML)并选用TN93+G 核苷酸替代模型构建系统进化树,各分支节点的置信值通过1000 次自举(Bootstrap)抽样进行评估[18]。

表1 本研究中用于构建系统发育树的菌株信息及基因序列登记号Table 1 Collection details and GenBank accession numbers of isolates used for phylogenetic analysis in this study
2 结果与分析
2.1 病害症状与致病性测定
该病害主要危害叶片,发病初期在叶片上可见褐色小点,扩展以后形成浅褐色圆形或近圆形病斑,病斑边缘颜色较深,发病后期在叶片病健交界处产生一圈裂纹,病斑中央组织脱落可形成穿孔。有时多个病斑相互愈合形成大斑,脱落后形成大的穿孔(图2A)。
致病性测定结果显示,有伤接种条件下供试菌株CH-1 和CH-3 均可使臭椿叶片发病(图2B,2C),接种1~2 d 后在接种部位可见褐色斑点,接种5~7 d 后一些病斑在病健交接处产生离层,进一步发展形成穿孔症状(图2B,2C),与自然发病症状完全相同。发病组织再分离,获得分离物经鉴定与接种菌株相同,由此证明分离获得的菌株为致病菌。无伤接种和对照未见发病。

图2 臭椿炭疽病症状Fig.2 Symptoms of anthracnose on Ailanthus altissima
2.2 病原菌的形态学鉴定
从河南省洛阳市嵩县天池山国家森林公园采集的臭椿病叶样品中分离获得11 个菌株,所有菌株在PDA 上菌落表现一致,菌落平整且边缘整齐,气生菌丝较密,棉絮状,菌落初始为白色,几天后菌落中间出现灰绿色(图3A),背面有黑色素沉积(图3B)。25℃培养5 d 的菌落直径为6.18 ± 0.67 cm。菌落上产生的分生孢子堆白色到浅黄色,显微镜检可见分生孢子为圆柱状,两端钝圆,单孢,无色(图3C),孢子大小(12.01~18.10)μm×(4.66~6.90)μm,平均长14.55 ± 1.22 μm,宽5.79 ± 0.44 μm。附着胞浅至深棕色,椭圆形或近球形(图3D,3E),大小(6.20~7.96)μm×(4.68~6.79)μm,平均长7.85 ± 1.12 μm,宽5.62 ± 0.49 μm。在PDA 上培养15 d 左右可形成子囊壳,子囊棍棒状,内含8 个子囊孢子,子囊孢子梭状,两端钝圆,稍弯曲(图3F,3G),子囊孢子大小(13.83~22.53)μm×(4.05~6.29)μm,平均长18.35 ± 1.65 μm,宽5.20 ± 0.53 μm。根据形态学和培养特征初步鉴定分离菌株为炭疽菌(Colletotrichumspp.)。[19-20]
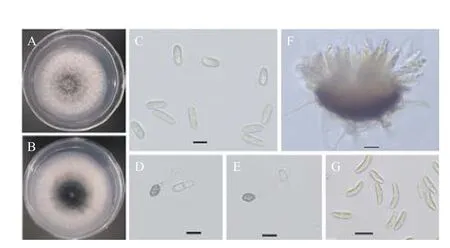
图3 臭椿炭疽病病原菌的纯培养和形态特征Fig.3 Morphology and cultural characteristics of Colletotrichum fructicola from Ailanthus altissima
2.3 病原菌的分子生物学鉴定
对菌株CH-1 和CH-3 的ITS、ACT、TUB2、GAPDH和CHS-1基因进行扩增和测序,得到大小分别为557 bp、289 bp、722 bp、280 bp 和299 bp的特异性片段。将序列提交至GenBank 数据库(表1),通过BLAST 搜索和比对,结果显示菌株CH-1 和CH-3 的ITS 和ACT基因序列与胶孢炭疽菌复合种(C.gloeosporioidesspecies complex)内的多个种序列相似性达到99%以上;菌株CH-1 和CH-3 的TUB2、GAPDH和CHS-1基因序列与果生炭疽菌(C.fructicola)序列相似性达到99%以上。从NCBI 上选取相关序列联合构建系统发育树,以C.boninense为外群,进行多基因系统发育分析,结果显示,供试菌株CH-1 和CH-3 与果生炭疽菌(C.fructicola)聚于同一分支,自展支持率为99%(图4)。结合形态学特征和多基因系统发育分析,最终鉴定在河南省洛阳市嵩县天池山国家森林公园发生的臭椿炭疽病病原菌为果生炭疽菌(C.fructicola)[21-22]。
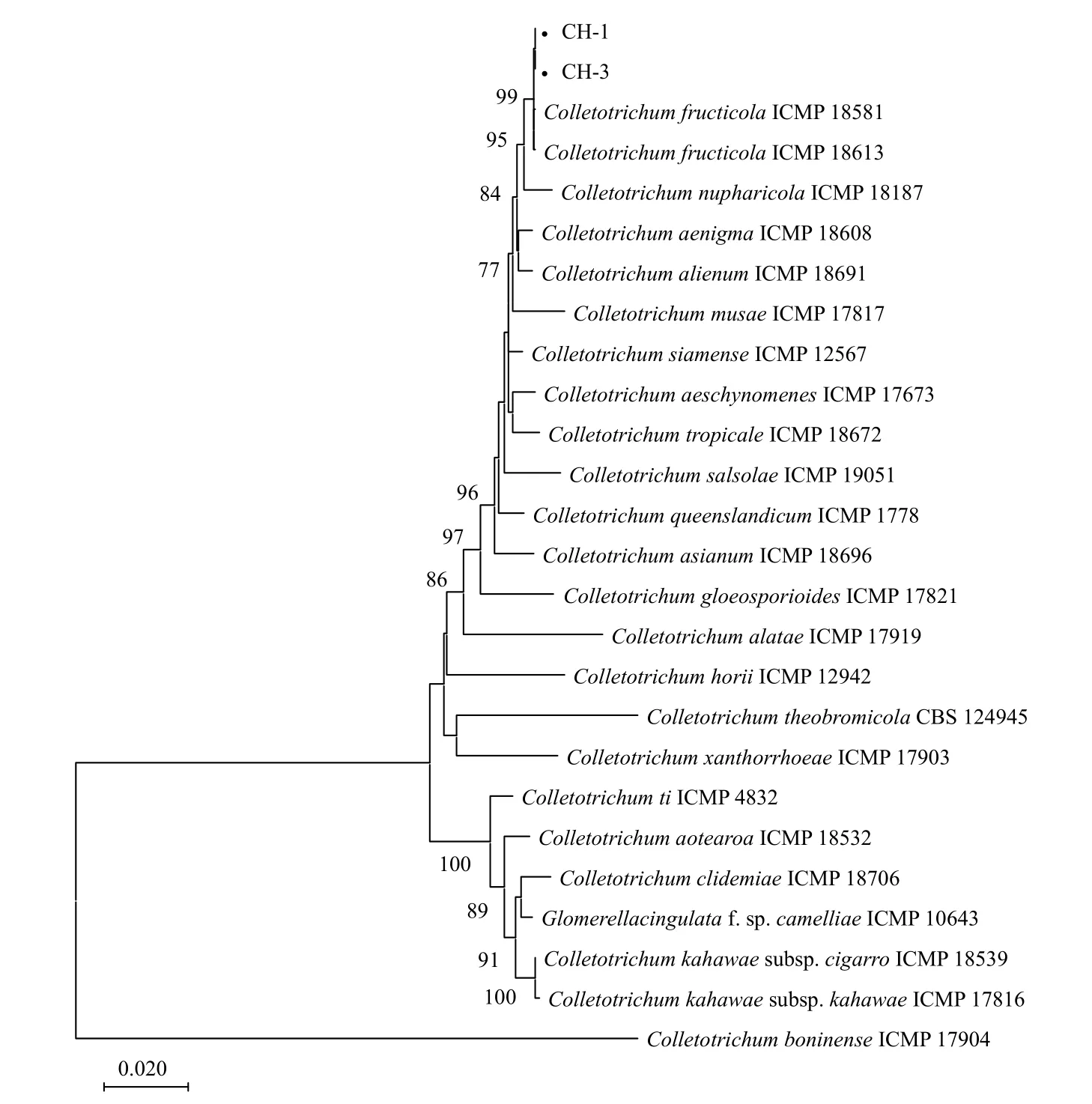
图4 基于最大似然法构建臭椿炭疽病病原菌及其相关种的多基因系统发育树Fig.4 Phylogenetic tree based on sequences of Colletotrichum isolates from Ailanthus altissima and related species using maximum likelihood method
3 讨论
本研究在河南省洛阳市嵩县天池山国家森林公园进行病害调查,发现臭椿炭疽病严重发生,对发病叶片进行病原菌分离,共获得到11 个分离物,它们在PDA 上菌落特征表现一致,正面菌落先白色后灰色,背面黑色。菌落上产生的分生孢子堆白色到浅黄色,分生孢子圆柱状,两端钝圆,单孢,无色。由于炭疽菌属(Colletotrichum)内种类繁多、形态简单,能够用于形态鉴定的特征较少,造成其种间界限不明确、分类鉴定困难[19,23]。通过形态学难以确认11 个分离物确切的种,初步鉴定为炭疽菌(Colletotrichumspp.)。
近年来,分子生物学技术在真菌分类鉴定中的应用,对炭疽菌属的分类鉴定方法产生了深刻影响[19,24]。rDNA-ITS 序列分析是炭疽菌分子鉴定中应用最早、最多的方法,为许多物种的鉴定及系统进化分析提供了有力工具[23-24]。研究发现,仅基于rDNA-ITS 序列并结合形态特征进行系统进化分析或物种鉴定存在一定的局限性,对于一些近缘相似种和复合种,仍不能准确反映和有效识别其亲缘关系[19,24]。目前,基于形态学和多基因序列的系统发育分析方法被众多研究者接受[25-27]。随着炭疽菌多基因系统学研究的深入,将炭疽菌属真菌划分为14 个复合种(C.acutatum、C.boninense、C.caudatum、C.dematium、C.destructivum、C.dracaenophilu、C.gigasporum、C.gloeosporioides、C.graminicola、C.magnum、C.orbiculare、C.orchidearum、C.spaethianum和C.truncatum)和部分独立种[19,28-29]。Weir 等采用ITS、ACT、CAL、CHS-1和GAPDH基因对大量炭疽菌株进行多基因序列分析和形态学鉴定,明确了C.gloeosporioides复合种内包含了22 个种和1 个亚种[21]。目前,C.gloeosporioides复合种已接受41 个合格种,其中大多数为植物病原菌[21,25,28,30]。本研究中对分离菌株进行形态学观察和多基因(ITS、ACT、TUB2、GAPDH和CHS-1)系统发育分析的结果显示,它们均为C.gloeosporioides复合种内的果生炭疽菌(C.fructicola)。
果生炭疽菌(C.fructicola)首次在泰国咖啡果实上发现[20],之后陆续报道该病菌可侵染多种经济作物,如梨[21]、苹果[22]、柑橘[31]、烟草[32]、木薯[33]和芒果[34]等,引起叶片坏死和果实腐烂。Weir等[21]研究发现果生炭疽菌具有明显的地域多样性和生物多样性,仅依赖形态学特征或rDNA-ITS 序列均很难对其准确鉴定。目前,推荐使用多基因序列的系统发育分析对其进行准确鉴定[21,27]。
4 结论
本研究对采自河南省洛阳市嵩县天池山国家森林公园的臭椿炭疽病样品进行了病原菌分离与纯化,致病性测定证明分离物可侵染臭椿叶片,并引起与林间症状一致的炭疽病;病原菌形态特征观察及多位点基因(ITS、ACT、TUB2、CHS-1和GAPDH)系统发育分析表明,引起该地区臭椿炭疽病的病原菌为果生炭疽菌(C.fructicola)。该研究结果可为深入研究臭椿炭疽病的发生流行规律以及制定防治策略奠定基础。
