新疆野果林的新害虫
——杏树鬃球蚧(半翅目:蚧总科:蚧科)
2021-12-29王玉丽吕昭智令狐伟高桂珍
王玉丽,吕昭智,令狐伟,高桂珍*
(1.新疆农业大学林学与园艺学院,新疆 乌鲁木齐 830052;2.青岛农业大学植物医学学院,山东 青岛 266109)
新疆天山野果林有丰富的果树种质资源基因[1]。其中,野杏(Armeniaca vulgarisLamarck)是主要的资源树种之一,被认为是世界栽培杏的原生起源种群[2]。野杏对当地生态环境健康、生物多样性维持具有重要作用,对林果业持续发展、旅游产业等具有重要经济价值[3-4]。然而,2019 年一种介壳虫在新疆天山野果林暴发成灾,大量野杏树受到危害,且危害呈扩散趋势。野杏受害后树冠枝条干枯,树势衰弱,受害严重时甚至导致整株死亡,此种介壳虫对整个野杏群落的生存构成重大威胁。
经北京林业大学武三安教授鉴定,野果林中为害野杏的介壳虫为杏树鬃球蚧(又名杏球蚧、圆球蜡 蚧)(Sphaerolecanium prunastriBoyer de Fonscolombe),其隶属于半翅目(Hemiptera)蚧总科(Coccoidea)蚧科(Coccidae)鬃球蚧属(Sphaerolecanium)。该蚧产于古北区的亚热带地区[5],目前在土耳其、俄罗斯[6]、比利时、美国[5]等多个国家均有分布,在我国主要分布在辽宁、河北、山东[7]及陕西[8]等地区。杏树鬃球蚧是多食性昆虫,其寄主包括番杏科(Aizoaceae)[9]、桑科(Moraceae)[10]、禾本科(Poaceae)[11]、鼠李科(Rhamnaceae)[12]及蔷薇科(Rosaceae)[13],这给植物资源丰富的野果林带来巨大的潜在威胁。目前,在我国杏树鬃球蚧尚未见系统研究报道。本研究于2019—2020 年通过室内观察和田间调查相结合,对杏树鬃球蚧的形态特征及生物学特性进行系统研究,旨在明确该虫的薄弱环节,以期为它的综合防治提供理论依据。
1 材料与方法
1.1 供试虫源
杏树鬃球蚧采自新疆巩留县的野杏分布区(43°14′51.10″~43°25′72.59″N,82°06′14.58″~82°51′07.34″E,1127~1329 m)。混生树种包括野苹果(Malus sieversiiLedeb.Roem.)、准噶尔鼠李(Rhamnus songoricaGontsch)、小花忍冬(Lonicera tataricaL.var.micrantha)、黑果小檗(Berberis heteropodaSchrenk)、天山绣线菊(Spiraea tianschanicaPojark)等。
1.2 试验方法
1.2.1 形态特征 2019 年6 月—2020 年4 月在巩留县库尔德宁镇阔克塔勒村河滩地(43°25′64.81″~43°52′59.64″N,82°81′14.91″~82°82′56.41″E,海拔1127~1138 m),于杏树鬃球蚧的整个发生期分阶段采回带虫枝条。2019 年6 月20 日采集卵100粒、6 月27 日采集1 龄若虫100 头、10 月24 日采集越冬前2 龄若虫100 头,2020 年4 月11 日采集越冬后雌雄分化期的2 龄若虫各87 头,4 月18 日采集雌成虫及蛹各100 头,在体式显微镜(蔡康,XTL-3200,含测微尺)下观察测量并记录其体长和体宽。
1.2.2 生活史 试验样地同1.2.1,2019 年6 月—2020 年6 月设置3 个采样点,在每个采样点选取5 株树,且样树距离大于15 m。每株树上标记东、南、西、北4 个20 cm 小枝,观察杏树鬃球蚧发育情况并记录发生量,每5 d 调查1 次。并随机抽取10 头带回,镜检统计各虫态发育进度。
1.2.3 越冬死亡率 2019 年7 月在巩留县库尔德宁镇阿勒玛塞(43°14′35.08″~43°14′43.89″N,82°49′17.57″~82°49′43.42″E,海拔1161~1367 m)设置6 个样地,每样地各选取20 个20 cm 枝条,记录枝条上杏树鬃球蚧发生量,2020 年4 月再次调查,记录杏树鬃球蚧存活数量,并计算其越冬死亡率。

1.2.4 性比 2020 年4 月杏树鬃球蚧处于雌雄分化期,在巩留县设置野核桃沟、八连、小莫合、大莫合及萨哈5 个样地,每样地随机采集15~20 个10 cm 枝条,记录雌虫和雄虫的发生量,并计算雌雄性比。
1.2.5 雌成虫产卵量及卵的发育 试验样地同1.2.1,2020 年5 月15 日随机固定带虫枝条50 个,每隔24 h 观察枝条上杏树鬃球蚧是否产卵,从开始产卵至1 龄若虫出现,期间每天采集带虫枝条,随机在枝条上抽取10~20 头雌成虫,记录单头雌虫的产卵量。室内设置15、20、25、30 和35℃5 个温度梯度,将样地采回的初产蚧卵50 粒放置在垫有杏树叶片及加湿滤纸的培养皿中,置于黑暗、(65 ± 5)% RH 的培养箱(广州恒创,PYX-300Q-C)中,每温度10 个重复,每隔24 h 观察并记录卵孵化情况。采用线性回归的方法,即以卵的发育速率作为因变量(y 轴),不同温度作为自变量(x 轴),估算卵的发育起点温度和有效积温[14]。
1.2.6 杏树鬃球蚧在野杏不同危害部位的发生量 不同方位:2019 年7 月在巩留县野核桃沟、小莫合、大莫合、八连、萨哈、伊力格代6 个样地(海拔1219~1329 m),各选取15株树,每株树东、南、西、北4 个方位各取1 个50 cm 枝条,记录杏树鬃球蚧(若虫)发生量。不同年生枝条:试验样地同1.2.1,2020 年6 月在野杏树上各选取50 个1~4 年生10 cm 枝条,记录杏树鬃球蚧(若虫)发生量。同一枝条的阳面和阴面:试验样地同1.2.1,2019 年6 月设置3 个采样点,每采样点选取15 株树,每株树各采集4 个20 cm 枝条,记录同一枝条阴阳面的杏树鬃球蚧(雌成虫)发生量。
1.3 数据分析
数据整理使用Microsoft Office Excel 2010 软件完成,应用SPSS 23.0 软件对越冬死亡率、不同温度下卵的发育历期、不同方位、不同年生枝条杏树鬃球蚧发生量数据进行单因素方差分析,以Duncan 检验比较差异显著性(P<0.05)。雌雄虫发生量和同一枝条阴、阳面发生量以独立样本T 检验比较差异显著性(P<0.05),作图、表数据为均值 ± 标准误(mean ± SE)。采用Origin 8.0 软件进行计算并作图。
2 结果与分析
2.1 形态特征
2.1.1 卵 卵椭圆形,长为0.23~0.45 mm;宽为0.11~0.31 mm,长宽比1.62,其大小随着时间的增加而变大(图1A)。初期为白色,半透明,中期转变为淡粉色,微透明,后期在解剖镜下可看清1 龄若虫轮廓。

图1 不同虫态杏树鬃球蚧Fig.1 Different insect states of Sphaerolecanium prunastri
2.1.2 若虫 1 龄若虫虫体粉红色,椭圆形,长为0.29~0.43 mm,宽为0.18~0.32 mm,长宽比1.35(图1B)。初孵时体表光滑,单眼1 对,圆形,红棕色;足发达,足基节和转节占足长1/6、腿节占足长1/3、胫节1/3、跗节1/6;体躯分节明显,9 节,长端毛1 对,腹末1 对端丝上有两根白色长丝状尾须伸出。
2 龄若虫虫体由粉红色转变为深褐色,椭圆形,长为0.66~0.82 mm,宽为0.45~0.76 mm,长宽比1.44(图1C)。越冬后的2 龄若虫,虫体黄褐色,虫体上有大量黑色条斑(图1D)。越冬后2 龄若虫雌雄两性分化,未来分化为雌性的2 龄若虫虫体比分化为雄性的小,虫体微微向上拱起,颜色呈现橘黄色;分化为雄性的2 龄若虫虫体略扁平呈椭圆形,颜色暗,呈现黑褐色(图1E)。分化为雌性的2 龄若虫长为0.95~1.64 mm,宽为0.48~1.07 mm,长宽比1.47;分化为雄性的2 龄若虫长为1.14~1.93 mm,宽为0.53~1.05 mm,长宽比1.86。
2.1.3 雌成虫 雌成虫颜色由初期的黄褐色到后期的深棕色,半球形,在膨大期产生蜜露,长为0.95~2.20 mm;宽为1.10~1.78 mm;高为0.50~1.04 mm,背部有不规则凹点,生殖期时,把大量卵产于蚧壳下(图1F)。
2.1.4 雄蛹壳 蛹壳,白色,光滑,不透明,一捏呈碎片状,有臀裂,呈120 度角,圆滑,蛹壳长1.52~2.17 mm,宽0.75~1.19 mm,长宽比1.73。(图1G)。
2.1.5 雄成虫 雄虫把蛹壳挣破,一对白色线状尾须从蛹末尾处伸出,雄成虫虫体红褐色,头胸腹分段明显。触角丝状,单眼大,胸部为倒等腰梯状,翅白色,微透明,翅脉简单,交配器呈锥状(图1H)。
2.2 生物学特性
2.2.1 生活史 杏树鬃球蚧在新疆野果林1 年发生1 代,主要以2 龄若虫在枝条上越冬。3 月下旬至4 月上旬越冬2 龄若虫出蛰,随后固定并雌雄开始分化。4 月下旬至5 月上旬雄性若虫发育为预蛹,再化蛹至羽化为成虫,羽化后立即寻找雌成虫交配,雄成虫寿命短,交配完不久死亡。雌成虫膨大并分泌大量蜜露,但体壁尚未硬化。5 月下旬至6 月上旬雌成虫蜡质壳高度硬化且开始产卵。6 月上旬至中旬卵孵化,1 龄若虫出现,并在枝条、叶片、果实上活动。杏树鬃球蚧在6 月中旬至7 月上旬多在枝条分布,随后蜕皮为2 龄若虫,虫体表面逐渐分泌蜡丝,直至把虫体完全覆盖,10 月开始越冬(表1)。

表1 新疆杏树鬃球蚧生活史Table 1 Life history of Sphaerolecanium prunastri in Xinjiang(2019—2020)
2.2.2 越冬死亡率 不同海拔样地(1161~1367 m)杏树鬃球蚧越冬死亡率分别为77.31%、72.31%、67.96%、84.87%、82.68%和90.40%,不同样地杏树鬃球蚧的越冬死亡率均大于65%,其中只有海拔1367 m 样地的杏树鬃球蚧越冬死亡率显著大于1203 m海拔样地,其它样地之间无显著差异(F=2.148,df=5,P>0.05)(图2)。

图2 杏树鬃球蚧越冬死亡率(2020 年4 月)Fig.2 Overwintering mortality of Sphaerolecanium prunastri (April 2020)
2.2.3 性比 通过对杏树鬃球蚧雌雄若虫发生量的调查,结果表明:雌性杏树鬃球蚧发生量为108.95头·10 cm−1枝条,雄性杏树鬃球蚧发生量为104.76头·10 cm−1枝条,雌性若虫和雄性若虫发生量无显著差异(F=0.126,df=1,P>0.05)(图3),雌雄性比为1.02∶1。

图3 杏树鬃球蚧雌雄虫发生量(2020 年4 月)Fig.3 Number of female and male insects of Sphaerolecanium prunastri(April 2020)
2.2.4 雌成虫产卵量及卵的发育 2020 年5 月22日在样地初次发现雌成虫产卵,从5 月22 日—5月30 日卵量开始不断增加,由5 月22 日的190 粒不断上升到5 月30 日的527 粒(图4),5 月31 日1 龄若虫出现。

图4 雌成虫的产卵情况(2020 年5 月)Fig.4 Spawning of female adults (May 2020)
在15~35℃条件下,杏树鬃球蚧卵的发育历期随着温度升高先缩短后延长,在30℃时卵的发育历期最短,为3.86 d。杏树鬃球蚧卵的发育速率与温度的关系表达式为y=0.012 3x−0.117 1,相关系数0.992 1(F=251.5593,P<0.05),相关性检验达到显著水平,说明卵发育速率与温度之间具有极好的相关性。35℃卵的发育速率在直线段外,因此没有包含在线性回归中(图5)。卵的发育起点温度为9.50℃,有效积温81.17 DD。在20~35℃恒温下卵的存活率均高于98%,而15℃时卵的存活率仅为36.6%(表2)。

表2 不同温度下杏树鬃球蚧卵的存活率Table 2 The survival rate of the eggs of Sphaerolecanium prunastri at different temperatures

图5 杏树鬃球蚧卵发育速率与温度的相关性Fig.5 The correlation between egg development rate and temperature of Sphaerolecanium prunastri
2.2.5 杏树鬃球蚧在野杏不同危害部位的发生量 杏树鬃球蚧1 龄若虫在东、南、西、北4 个不同方位的发生量无显著性差异(F=0.484,df=3,P>0.05)(图6A)。杏树鬃球蚧1 龄若虫在1~4 年生野杏枝条发生量呈现3 年生>4 年生>2 年生>1 年生枝条的规律,存在显著差异(F=40.208,df=3,P<0.05)(图6B)。杏树鬃球蚧雌成虫在同一枝条阴、阳面的发生量分别为22.03 和6.23头·20 cm−1枝条,即同一枝条阴面发生量显著多于阳面(F=85.58,df=1,P<0.05)(图6C)。
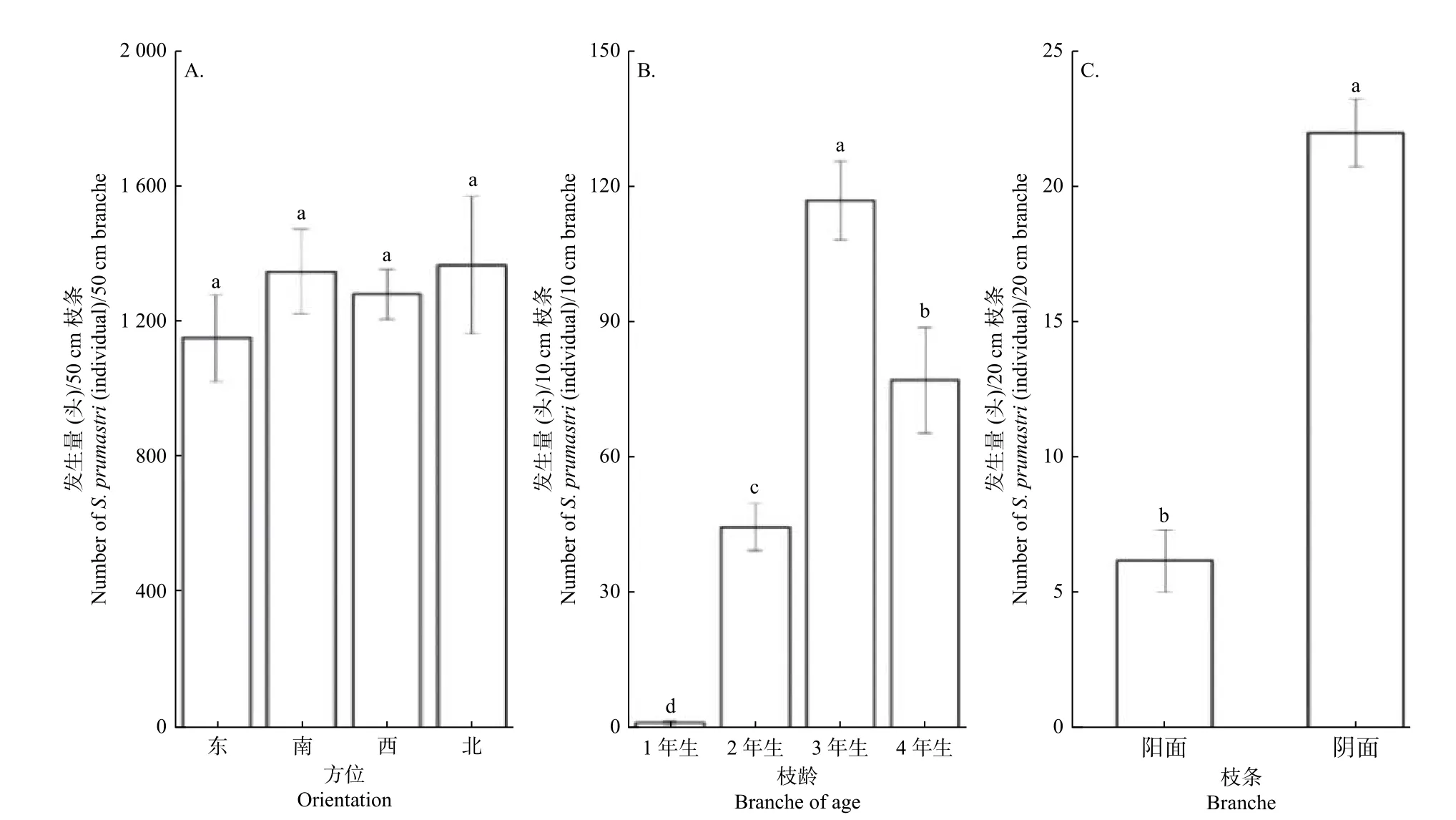
图6 杏树鬃球蚧在野杏不同危害部位的发生量Fig.6 Number of Sphaerolecanium prunastri in different damaged parts of wild apricot
3 讨论
杏树鬃球蚧在新疆野果林1 年发生1 代,以2 龄若虫在枝条上越冬,该蚧发育和野杏物候期关系密切。2020 年4 月野杏处于现蕾及花期,杏树鬃球蚧越冬2 龄若虫处于出蛰期,能短距离活动并重新选择刺吸部位固定;5 月杏树鬃球蚧雌虫膨大期时营养需求大,此时野杏处于坐果及果实膨大期,养分充足,可供雌虫完成发育。在杏树鬃球蚧发育过程中,2 龄若虫出蛰期、膨大期(3 月下旬至4 月下旬)和1 龄若虫涌散期(6 月上、中旬)是该蚧虫体柔软且裸露在外的薄弱环节,同时也是化学防治该蚧的最佳时期。
介壳虫产卵量受多种因素的影响,其中包括交配行为[15]、寄主[16]、体型大小[17]、温度[18]等。杏树鬃球蚧在比利时产卵量达3000 粒[5],在土耳其产卵量为400~800 粒[6],而在中国新疆野果林产卵量为527 粒,这可能与本实验中对产卵量的监测到出现1 龄若虫即停止调查、海拔[19]、地理环境不同[20]等有关。不同介壳虫雌雄比不同,同属蚧科的朝鲜球坚蚧(Didesmococcus koreanusBorchsenius)雌雄比为1∶1.25[21]、吐伦球坚蚧(Rhodococcus turanicusArch.)雌雄比约为1∶2[22],本试验调查结果杏树鬃球蚧雌雄比为1.02∶1。
温度在介壳虫的生长发育过程中起重要作用,对介壳虫种群的发生具有显著影响[22-24]。本研究调查发现,海拔最高的样地内杏树鬃球蚧的越冬死亡率最高,与何雪香和刘清浪[25]、王光旭等[19]研究结果一致,这是由于海拔升高,温度降低所致。本研究中2020 年开春调查的杏树鬃球蚧越冬死亡率及其卵在不同温度下的存活率结果,均表明低温不利于该蚧的生存。袁福香等[26]研究也表明,冬季气温越低,介壳虫若虫的越冬死亡率越高。尽管杏树鬃球蚧在寒冷冬季具有较高的死亡率,但冬季积雪在一定程度上有利于介壳虫的越冬存活,而其余温度较高野果林区所存活下来的杏树鬃球蚧也可能会在夏季重新传入并继续生存发展[27]。因此,如何通过温控试验确定杏树鬃球蚧冬季所能忍受的极端最低气温指标,或夏季发育所需的积温指标,结合野果林不同林区气候条件,可以为进一步科学防控该蚧在野果林的扩散提供理论依据。
昆虫属于变温动物,发生期受环境影响大[28-29]。陈宏灏等研究甘草胭珠蚧(Porphyrophora sophoraeArch.)卵的发育起点温度和有效积温分别为9.27℃和574.98 DD,为该蚧若虫发生提供预测并进行关键期的有效防治[30]。因此,明确杏树鬃球蚧的发育速率和温度之间的关系是预测预报的关键。本实验采用直线拟合温度和发育速率的关系得到相关系数为0.9413,杏树鬃球蚧的发育起点温度为9.50℃,有效积温为81.17 DD。因此可结合当地气温条件对该蚧进行发生期的预报,可为防控该蚧提前做好准备,以免错过最佳防治期,对该虫的防治进行更有效地指导。今后可对自然条件下杏树鬃球蚧卵的发育起点温度和有效积温展开进一步研究。
杏树鬃球蚧对野杏的威胁大。伊犁地区天山野杏林是我国野杏的重要集中分布地,面积为660 hm2,西起霍城县,伊宁县,向东延伸至新源县、巩留县及察布查尔县[31]。杏树鬃球蚧的寄主植物广泛,目前调查发现野果林中野苹果、准噶尔鼠李、天山绣线菊等野生植物和周边栽培果园中已有杏树鬃球蚧寄生并危害,该蚧对野果林中大量珍贵树种和栽培果树产生威胁。同时,野果林中病虫害的发生具有复杂性。杏树鬃球蚧在野杏上的大量发生,导致野杏树势衰弱,野杏上也存在病原菌侵入,其中有杏树穿孔病[32-33]、疫霉菌等[34],目前病虫的交互作用不明确,可进一步开展研究。同时,调查中发现有朝鲜毛球蚧与杏树鬃球蚧在局部地区混合发生危害野杏,两种蚧都属于蜡蚧科,目前两种蚧的种间关系并不明确,为后期防治工作加大难度。
4 结论
杏树鬃球蚧属于性二型昆虫,主要分布在野杏3 年生枝条,在树冠4 个不同方位发生量无显著差异。杏树鬃球蚧在新疆野果林1 年发生1 代,主要以2 龄若虫在枝条上越冬,越冬死亡率高。2 龄若虫出蛰期、膨大期(3 月下旬至4 月下旬)和1龄若虫涌散期(6 月上、中旬)是杏树鬃球蚧的薄弱环节,建议抓住以上两个关键时期开展综合防治。
