Fine-scale activity patterns of large- and medium-sized mammals in a deciduous broadleaf forest in the Qinling Mountains, China
2021-12-24YukeZhangXiangboHeXuehuaLiuMelissaSongerHaishanDangQuanfaZhang
Yuke Zhang · Xiangbo He · Xuehua Liu ·Melissa Songer · Haishan Dang · Quanfa Zhang
Abstract The composition of animal species and interactions among them are widely known to shape ecological communities and f ine-scale (e.g., < 1 km) monitoring of animal communities is essential for understanding the relationships among animals and plants. Although the co-existence of large- and medium-sized species has been studied across dif ferent scales, research on f ine-scale interactions of herbivores in deciduous broadleaf forests is limited. Camera trapping of large- and medium-sized mammals was carried out over a 1 year period within a 25 ha deciduous broadleaf forest dynamics plot in the Qinling Mountains, China. Fourteen species of large- and medium-sized mammals, including six carnivores, six ungulates, one primate and one rodent species were found. Kernel density estimations were used to analyse the diel or 24 h activity patterns of all species with more than 40 independent detections and general linear models were developed to explore the spatial relationships among the species. The combination of overlapping diel activity patterns and spatial associations showed obvious niche separation among six species: giant panda ( Ailuropoda melanoleuca David), takin ( Budorcas taxicolor Hodgson),Reeves’s muntjac ( Muntiacus reevesi Ogilby), tufted deer( Elaphodus cephalophus Milne-Edwards), Chinese serow( Capricornis milneedwardsii David) and wild boar ( Sus scrofa Linnaeus). Long-term f ine-scale monitoring is useful for providing information about the co-existence of species and their interactions. The results demonstrate the importance for f ine-scale monitoring of animals and plants for improving understanding of species interactions and community dynamics.
Keywords Large- and medium-sized mammals · Finescale interaction · Camera trapping · Spatiotemporal patterns · Forest dynamics plots · Qinling mountains
Introduction
Understanding the distribution and interactions of animals and plants and their inf luencing factors is fundamental to ecology and crucial for ef fective conservation management(Tobler et al. 2008; Northrup and Gerber 2018; Krohner and Ausband 2019). The dynamics of animal diversity and associations inf luence the structure and function of ecological communities (Schneider et al. 2016). For example, seed and seedling mortality due to herbivory feeding, as well as animal-carry seed dispersal, af fect plant population dynamics, community structure and the maintenance of species diversity (Barnes 2001; Li and Zhang 2001; Gomez 2005;Beguin et al. 2016). Interactions between animal species also have critical impacts on population dynamics. Predators inf luence herbivore populations via top-down predation (Sinclair et al. 2003). Sympatric species using similar resources, may have intense competition and therefore achieve co-occurrence by dif ferentiating their food sources,temporal activities and/or spatial habitats (Liu et al. 2017;Haidir et al. 2018). In mammalian communities, the coexistence of carnivores and their underlying mechanisms have been studied widely (Bu et al. 2016; Gantchof f and Belant 2016; Croose et al. 2019; Santos et al. 2019).
However few studies have explored the coexistence of herbivores, especially in deciduous broadleaf forests. Farnsworth et al. ( 2002) modelled ungulate grassland ecosystems and found that coexistence between grazers depends on how they partition resources. Yang et al ( 2018) studied the dynamics of microhabitat selection and interactions of four ungulate species in coniferous forests in northern China and found that overlaps of their spatial patterns were af fected by seasonal habitat factors. Most studies on the interactions of large- and medium-sized animals were carried out at scales ranging from dozens of kilometres (Jia et al. 2014; Michalski et al. 2015) to hundreds (Santos et al. 2019). However,examination of the interactions of herbivores in deciduous broadleaf forests such as in the Qinling Mountains at a f inescale (≤ 1 km), a region of documented global biodiversity importance, is lacking. Since monitoring plots of vegetation communities is usually carried out at less than 1 km, the dif ferences between the scales of mammal and vegetation community data make their integration challenging.
The centre for tropical forest science (CTFS)–forest global earth observatory (ForestGEO) is a global network of forest research sites strategically poised for monitoring,understanding and predicting forest responses to global change (Anderson-Teixeira et al. 2015). This international partnership currently includes 59 long-term forest dynamics plots (FDPs) with a median size of 25 ha (ranging from 2 to 120 ha) around the world and studied using standardized methodologies that enable comparison across sites.Although the use of camera traps for studying terrestrial mammals has become increasingly popular in recent years,systematic monitoring specif ications for camera trapping in FDPs in areas other than the tropics are few (Tobler et al.2008; Ahumada et al. 2011; Xiao et al. 2014). However,camera trapping of large- and medium-sized mammals in FDPs (mainly 15 – 30 ha) with a density of 0.5 – 1 camera traps·ha −1 have been conducted in China since 2009, providing baseline data for exploring animal diversity and interspecies interactions (Xiao 2014). The studies have revealed species composition and activity patterns of mammals for tropical, subtropical and temperate forest areas (Li et al.2014b; Si and Ding 2014; Xie et al. 2014; Zhang et al. 2014)but the interrelationships among animal species has not been analysed.
One-year monitoring of large- and medium-sized mammals was carried out by camera trapping a 25-ha deciduous broadleaf FDP established in the temperate-subtropical transition zone in the Qinling Mountains (Xie et al. 2019).Species’ diel activity patterns were analyzed to answer the following questions: (1) what large- and medium-sized mammals live in this deciduous board-leaved forest plot;and (2) what are the spatiotemporal dynamics of activity patterns among dif ferent species on a f ine-scale?
Materials and methods
Study area
The study area was located in Foping Nature Reserve (FNR),33°33′ – 33°46′ N, 107°40′ – 107°55′ E on the southern slope of the Qinling Mountains in north-central China. The south-facing slope is in a subtropical zone with relatively wet summers and warm winters. The average annual temperature is approximately 13 °C, with the hottest temperature in July (average daily temperature 27 °C) and the coldest month is January (– 2 °C). Annual precipitation ranges from 950 – 1200 mm, most of which occurs between July and September (Yue et al. 2000). The study area is located within a permanent forest dynamic plot (FDP) established in 2014 and covering an area of 25 ha (500 m × 500 m) within a deciduous broadleaf forest inside the Reserve. The plot has a complex geographical structure, including deep valleys and steep slopes and elevations ranging from 1715 to 1836 m a.s.l. (Shi et al. 2019). A total of 119 tree species(DBH ≥ 1 cm) are distributed in the plot with deciduous species dominant (89%) and evergreen species (11%). The three most abundant species areQuercus alienavar.acutiserrataMaxim. in the canopy layer,Sorbus alnifolia(Siebold &Zucc.) K.Koch. in the sub-canopy layer andQuercus spinosaDavid in the shrub layer (Xie et al. 2019).
Camera trapping and photo analyses
Thirty camera traps were set out (Yian-weishi L710, Shenzhen, China) in the plot with a monitoring density of 1.2 cameras per ha with distances of about 100 m between cameras (Fig. 1). Monitoring ran from 18 October 2018 to 7 December 2019. All cameras were installed 60 – 140 cm above the ground, based on the topography and vegetation of the sites in order to provide a good view of the sampling area. Cameras were programmed to take two photos and a 10 s video for each trigger with no artif icial delay between consecutive triggers and were set to function continuously(24 h day −1 ) during the sample period.

Fig. 1 Distribution of camera traps in the forest dynamics plot in the Foping Nature Reserve
Consecutive detections (i.e., photographs and video) of the same individual < 30 min were considered to be an independent detection event (O’Brien et al. 2003). Independent detections of large- and medium-sized mammals were used for further analyses. Relative activity intensity (RAI) was def ined as the ratio of independent detections to the number of camera trap days (number of 24 h periods during which cameras were operating until the memory card was full or cameras were retrieved) and multiplied by 100. Thus, RAIijrefers to the relative activity intensity of speciesiat the trapping sitej(Eq. 1).

whereIdijrefers to the number of independent detections of speciesiin monitoring sitej,Ctdjthe number of camera trap days in monitoring sitej.
Species accumulation curves are commonly used to estimate the completeness of an inventory, which is especially important when looking at the changes in species diversity over space or time (Soberon and Llorente 1993; Tobler et al.2008). A species accumulation curve was produced using the package of “BiodiversityR” package in R (Kindt and Coe 2005) to assess whether our data collection ef fort was suff icient to capture species in the community. The curve portrays the cumulative number of species detected against the sampling ef fort per unit time, which can be camera days (the number of cameras used multiplied by the number of survey days) or survey days in the case of camera traps (Tobler et al.2008). We maintained standard deviations with the curve according to Kindt and Coe ( 2005) to indicate the standard deviation observed because, by chance, some combinations of sites may have few species while others may have many.
Spatiotemporal patterns of animal activity
The temporal patterns of animal activity in the environment form a behavioural and ecological metric that can provide an indicator of energetics, foraging ef fort and exposure to risk (Rowclif fe et al. 2014). To quantitatively describe the diel activity patterns, kernel density estimations were carried out using “overlap” packages in R (version 3.6.1), which treated the time stamp of independent detections as a random sample from continually-distributed time and plotted the changing of the probability density along the time axis(Ridout and Linkie 2009).
To describe interactions of the species, we estimated the coef ficient of overlap (Δ, ranges from 0 to 1, with 1 indicating complete overlap) for the activity patterns of any pair of species (> 40 independent detections) using “overlap”packages (Schmid and Schmidt 2006; Ridout and Linkie 2009). We used Δ1 estimator for small sample sizes (< 50 independent detections) and Δ4 estimator for large sample sizes (≥ 50 independent detections for both samples) as suggested by Ridout and Linkie ( 2009). We generated 999 times smoothed bootstrapping to estimate the conf idence interval(CI) and mean value of the Δ estimators.
The ef fects of abiotic and biotic factors on species activity patterns were explored using generalized linear models(GLMs). Three topographic variables, i.e., elevation, slope and aspect, were calculated from a 30 m resolution digital elevation model (DEM; http://www.gsclo ud.cn/, accessed Dec. 10th, 2019) using ArcGIS 10.6 (ESRI, Inc.). To represent the availability of water resources (e.g., rivers), the DEM was used to extract the potential watershed channel network in the study area, which was later ground-tested in the f ield. The Euclidean distance to the nearest river was then calculated. Considering the inf luence of vegetation on animal performance, the percentage of broadleaf trees for all the grids (a resolution of 30 m × 30 m) was calculated using the number of broadleaf trees divided by the total number of trees (DBH ≥ 1 cm, broadleaf and coniferous) in a grid. Finally, the f ive habitat variables for each of the 30 camera trap sites were extracted. The RAIs of those species with > 40 independent detections were used as biotic variables in the GLM models. We took turns using the RAI of one species to be the response variable and used the RAIs of the other species, together with the f ive habitat variables,to be predictors to build the global models.
To evaluate the correlations among the variables, pairwise correlations between all variables were examined and the variance inflation factor (VIF) calculated for the
models using the “vif” function in “nlme” package (Pinheiro et al. 2019). There was little multicollinearity in the models (|r|< 0.6 and VIF < 5) and no spatial-autocorrelation appeared after checking the semivariogram of the model using the “variogram” function in “nlme” package of R (Pinheiro et al. 2019).
All variables were centred and standardized to have zero mean and a standard deviation of 0.5 before parameter estimation in order to make them comparable within models and to eliminate the bias caused by the different detection probability of the species (Grueber et al.2011). The global models were run as model iterations using the dredge function from the MuMIn package in R(R project for Statistical Computing: https://www.r-proje ct.org/) (Bartoń 2019) and Akaike’s Information Criteria(AIC) was used to determine relative support for each sub-model. The top sub-models within 2 AIC of the best supported model were considered equally supported and were selected (Burnham and Anderson 2002). Finally,model-averaged effect estimates, 95% confidence intervals and relative variable importance for all variables included in the top sub-models were calculated.
Results
Capture frequencies and species list
Of the 30 camera traps, six were discontinued due to malfunction; the remaining accumulated 10,647 camera trapping days. A total of 1,314 independent detections were collected, including 1,127 for large- and medium-sized mammals (14 species), 167 for small mammals (< 1 kg,four species) and 20 for birds (eight species) (Table S1).The giant panda (Ailuropoda melanoleucaDavid) was the most abundant among the carnivores, followed by the yellow-throated marten (Martes flavigulaBoddaert),while the Asiatic black bear (Ursus thibetanusG. (Baron)Cuvier) was in very low numbers in the FDP. Although giant pandas are bears and have a carnivorous digestive system, over 99% of their food consists of plants (Schaller et al. 1985). Therefore, their relationships with ungulates were also examined. Reeves’s muntjac (Muntiacus reevesiOgilby) and tufted deer (Elaphodus cephalophusMilne-Edwards) had the highest relative abundance among the ungulates, while the Chinese goral (Naemorhedus griseusMilne-Edwards) had the lowest. The accumulation of species detected with increasing sampling effort was at first steep, but by 6,000 camera trap days, 13 species had been recorded, representing 92% of the total number detected (Fig. 2).

Fig. 2 Species accumulation curve with sampling ef fort for the community of large- and medium-sized mammals detected by camera trapping; the bars indicate + 2 and − 2 standard deviations
Diel activity pattern
Only six species of large- and medium-sized mammals, i.e.,giant panda, takin, Reeves’s muntjac, tufted deer, Chinese serow and wild boar had > 40 independent detections over the monitoring period. The giant panda was highly active in the morning and in the late afternoon until midnight, with two valleys around 300 and 1300 h (Fig. 3). Three of the ungulates, the takin, tufted deer and Reeves’s muntjac, had obvious crepuscular activity patterns, i.e., they were active in twilight conditions. Therefore, the coef ficients of overlap among the three were higher than 0.8 (Table 1, Fig. S1).In contrast, the Chinese serow showed a typical nocturnal pattern while wild boar showed peak activity between 1000 and 1800 h. These two species had the lowest overlap of diel activity patterns to each other (Δ = 0.4664).
Spatial relationships of activities among species
The giant panda models indicated that the intensity of wild boar activity was one of the dominant predictive variables,showing a positive association with giant panda activity(Table 2). In addition, the intensity of giant panda activity was higher in microhabitats with less broadleaf vegetation and at lower elevations. The percentage of broadleaf trees also ranked as the f irst dominant predictive variable of takin activity and the third for wild boar activity. Only the RAI of wild boar and tufted deer and broadleaved vegetation remained in the average top sub-models predicting the activity of Reeves’s muntjac, but only the former had a signif icant positive association. Aspect had a signif icant positive ef fect on the RAI of the tufted deer. Besides the RAI of the giant panda, the intensity of the activity of wild boar also can be explained by the activities of Reeves’s muntjac and takin.
*Pb, the percentage of broadleaf trees, was calculated using the number of broadleaf trees divided by the total number of trees (DBH ≥ 1 cm, broadleaf and coniferous)in a grid.

Fig. 3 Diel activity patterns of the six large- and medium-sized mammal species in the forest dynamics plot; letters “a” to “f”indicate giant panda ( Ailuropoda melanoleuca David), takin( Budorcas taxicolor Hodgson),tufted deer ( Elaphodus cephalophus Milne-Edwards), Reeves’s muntjac ( Muntiacus reevesi Ogilby), Chinese serow ( Capricornis milneedwardsii David)and wild boar ( Sus scrofa Linnaeus) respectively
Discussion
Species composition in the forest dynamics plots

Table 1 Coef ficient of overlap of species pairs in the forest dynamics plot in the Foping Nature Reserve
Due to its protected status, our study site within the Foping Nature Reserve has had little human-caused disturbance over the last century, making it ideal for examining the dynamics of mammal populations and their interactions(Liu et al. 2017). Six species of carnivores and six ungulates were recorded. Compared to previous camera trap studies in the low-elevation primary forests of the Qinling Mountains, four species were not observed in this study:the masked palm civet (Paguma larvataSmith), the Siberian weasel (Mustela sibiricaPallas), the Eurasian otter(Lutra lutraLinnaeus) and the forest musk deer (Moschus berezovskiiFlerov) (Zhang et al. 2019). This may be because the previous study spanned a wider elevation range, covered three years and the undetected species were represented by small populations (Zhang et al. 2019). The population of Chinese goral (Naemorhedus griseusMilne-Edwards) in this area experienced a steep decrease after 2009. Prior to 2009, the species was just less than the takin and wild boar populations, but it became the least detectedungulate in 2014 (Jia et al. 2014; Li et al. 2014a; Zhang et al. 2019). The decline is suspected to be a result of an infectious skin disease which causes high mortality. The recovery of this population may be slow since there were no macro carnivores predating diseased individuals and slowing the spread of the disease (Zhou 2017). However,the abundance of tufted deer and Reeves’s muntjac have increased since 2008 (Li et al. 2014a).
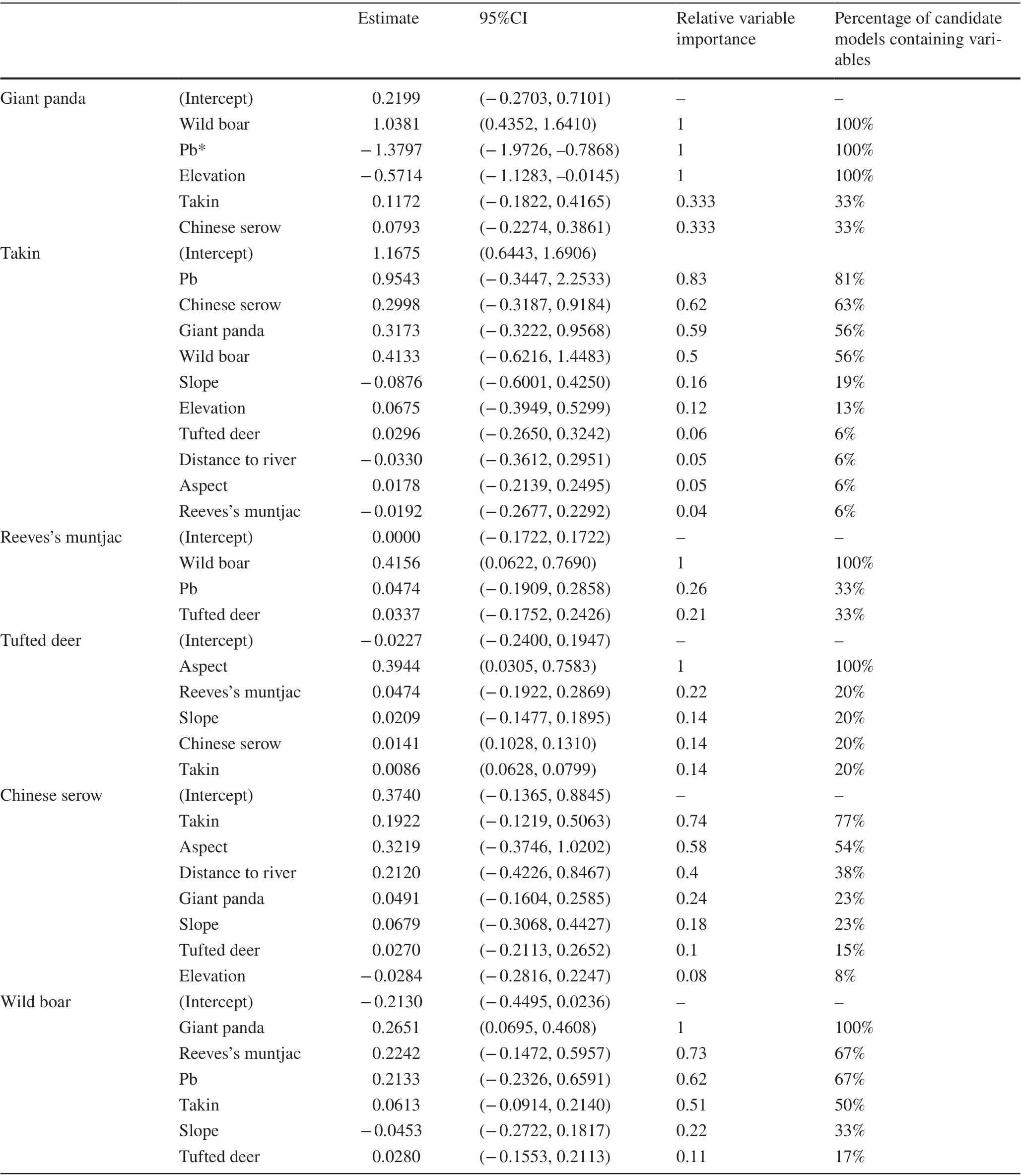
Table 2 Model-averaged mean ef fect estimates and conf idence intervals for the inf luence of predictors on relative activity intensity of the six species of large- and medium-sized mammals in the forest dynamics plot
Spatiotemporal associations of the species
Diel activity patterns, adaptations to environmental variability throughout the day, of giant pandas have been studied in the Qinling Mountains, in the Minshan Mountains(Wanglang Nature Reserve) and in the Qionglai Mountains(Wolong Nature Reserve), where they showed two activity peaks during the day, with one between 1500 and 1800 h which is consistent with our study (Hu 1987; Pan et al. 2001;Duan 2014). However, there are large variations across space and time: giant pandas in this study and in Wanglang showed the f irst activity peak between 800 and 1000 h and a depression between 300 and 400 h, while the two earlier studies in Qinling and Wolong showed lower activity levels during the morning, with an activity depression at 500−900 h(Hu 1987; Pan et al. 2001; Duan 2014). The diel activity patterns of takin, tufted deer, Reeves’s muntjac, Chinese serow and wild boar were consistent with previous studies in the Guanyinshan Nature Reserve on the southern slope of the Qinling Mountains, indicating stable patterns for those species across dif ferent scales (Jia et al. 2014; Wang et al.2015). However, the diel activity patterns of Chinese serow in our study area were dif ferent compared with the population in the Gaoligong Mountains in western Yunnan, where the species tended to be active in the afternoon and middle of the night (Chen et al. 2019). The reason for such dif ferences is unclear, but it may be a result of the wet climate(annual precipitation of 1200−3900 mm) in Gaoligong that drives the individuals to be active in order to keep warm, or the release of competition due to lower relative abundance of other sympatric ungulates in Gaoligong (Chen et al. 2016).
Theoretical studies show that species with completely overlapping niches cannot coexist and all but one of the species is eventually doomed to extinction (Levin 1970; Farnsworth et al. 2002). Spatial and temporal interactions between the six species were explicitly tested and niche separations were found among the giant panda and the f ive ungulates.Signif icant associations of spatial activity patterns appear among wild boar and giant panda and Reeves’s muntjac,but interspecif ic competition was low due to their temporal separation and, to some degree, dif ferent food resources.Takin, tufted deer and Reeves’s muntjac had very similar diel activity patterns but showed different microhabitat preferences which may relate to dif ferent use of resources.Chinese serow had a closer association with takin compared to the other three ungulates in terms of intensity of their spatial activities, but had the lowest overlapping diel activity patterns.
The percentage of broadleaf trees was an important habitat variable that inf luenced the intensity of activity of most of the species. In our study area, lower broadleaf composition and low-lying areas indicated a relative higher cover of bamboo,which partly explains the increased giant panda activity. The distance to the river and the slope in our models did not signif icantly inf luence the activity intensity of the species because the gradient of these environmental factors in 25 ha was small and thus their ef fects on microhabitat preference of the species were limited. However, aspect was more af fected by the local topography. The variation of aspect may cause dif ferences in microclimate, soils and the composition of vegetation, which inf luences the spatial activity patterns of mammals (Desta et al. 2004).
Limitations and prospect
The community structure (i.e., composition of species) af fects food, shelter and movements of mammals, thus inf luencing their perference of microhabitats. For example, the availability and quality of food resources as well as predation risks inf luence the spatial and/or temporal activity patterns of herbivores(Bakker et al. 2005; Wirsing et al. 2007). In addition, it was the vegetation diversity and growth, such as herbaceous plants and lianas rather than the canopy cover, that signif icantly inf luence habitat selection of some forest antelope species (Bowkett et al. 2008). Microhabitat factors such as food resources and vegetation growth may change with plant phenology and thus need to be surveyed along a time axis.
However, high-precision growth measurements in forest dynamics plots provide f ine-scale data of temporal and spatial variation in vegetation growth and productivity, making it possible to identify interactions among mammals and their habitat (Anderson-Teixeira et al. 2015). Conducting mammal monitoring in FDPs can also provide important information for understanding critical processes within plant communities. Therefore, future research should systematically combine the f ine-scale monitoring of mammals and plants together, along with microhabitat factors from ground surveys, to accurately predict mammal activity patterns.
In addition, in order to understand the relationships among mammals, future studies should focus on the segregation of food niches among species and the interactions among predators and prey.
Finally, our methods of camera trapping and modelling provide a reference for global ecosystem observations based on a network of forest dynamics plots. The results can also be used to assess ecosystem health and integrity and inform conservation strategies for rare species.
AcknowledgementsWe appreciate Foping Nature Reserve administration for support for the f ield work.
杂志排行

Journal of Forestry Research的其它文章
- Genome-wide identif ication and cold stress-induced expression analysis of the CBF gene family in Liriodendron chinense
- Decay rate of Larix gmelinii coarse woody debris on burned patches in the Greater Khingan Mountains
- Characterizing conservative and protective needs of the aridland forests of Sudan
- Point-cloud segmentation of individual trees in complex natural forest scenes based on a trunk-growth method
- Accuracy of common stem volume formulae using terrestrial photogrammetric point clouds: a case study with savanna trees in Benin
- Appropriate search techniques to estimate Weibull function parameters in a Pinus spp. plantation