Critical ef fects on the photosynthetic ef ficiency and stem sap f low of poplar in the Yellow River Delta in response to soil water
2021-12-24ChangxiWangHuanyongLiuJiangbaoXiaXianshuangXingShuyongZhang
Changxi Wang · Huanyong Liu · Jiangbao Xia ·Xianshuang Xing · Shuyong Zhang
Abstract To explore the critical relationships of photosynthetic ef ficiency and stem sap f low to soil moisture, twoyear-old poplar saplings were selected and a packaged stem sap f low gauge, based on the stem-heat balance method, and a CIRAS-2 portable photosynthesis system were used. The results show that photosynthetic rates ( P n ), transpiration rates ( T r ), instantaneous water use ef ficiency ( WUE) and the stem sap f low increased initially and then decreased with decreasing soil water, but their critical values were dif ferent.The turning point of relative soil water content ( W r ) from stomatal limitation to nonstomatal limitation of P n was 42%,and the water compensation point of P n was 13%. Water saturation points of P n and T r were 64% and 56%, respectively, and the WUE was 71%. With increasing soil water, the apparent quantum yield ( AQY), light saturation point ( LSP)and maximum net photosynthetic rate ( P nmax ) increased f irst and then decreased, while the light compensation point( LCP) decreased f irst and then increased. When W r was 64%,LCP reached a lower value of 30.7 μmol m −2 s −1 , and AQY a higher value of 0.044, indicating that poplar had a strong ability to utilize weak light. When W r was 74%, LSP reached its highest point at 1138.3 μmol∙m −2 s −1 , indicating that poplar had the widest light ecological amplitude and the highest light utilization ef ficiency. Stem sap f low and daily sap f low reached the highest value (1679.7 g d −1 ) at W r values of 56%and 64%, respectively, and then declined with increasing or decreasing W r , indicating that soil moisture signif icantly af fected the transpiration water-consumption of poplar. Soil water was divided into six threshold grades by critical values to maintain photosynthetic ef ficiency at dif ferent levels, and a W r of 64–71% was classif ied to be at the level of high productivity and high ef ficiency. In this range, poplar had high photosynthetic capacity and ef ficient physiological characteristics for water consumption. The saplings had characteristics of water tolerance and were not drought resistant. Full attention should be given to the soil water environment in the Yellow River Delta when planting Populus.
Keywords Gas exchange parameters · Sap f low · Soil moisture · Water use ef ficiency · Yellow River Delta
Introduction
PopulusL. is one of the most suitable genera for industrial plantations due to rapid growth and relatively good timber (Li et al. 2018; Liang et al. 2018). The hybrid,Populus×euramericana‘Neva’ is currently the most important fast-growing and high-yielding plantation species in the Yangtze River Basin and Huang-Huai-Hai region (Zhang et al. 2018). In China, this poplar is a major timber tree, with large planting areas and wide distribution. In the Yellow River Delta, the strong inf luence of the cutof f of the Yellow River, particularly the evaporation-precipitation ratio and seasonal water shortage, has reduced the productivity of poplar plantations, and soil moisture is a key factor in the growth and development of plantations. However, studies concerning the multilevel water gradients of poplar plantations in the Yellow River Delta are lacking; thus, photosynthetic physiological processes and the quantitative relationships between soil water and growth remain unclear.Moreover, the results of recent studies on the physiological ecology of drought resistance in trees are subject to greater restrictions when applied to water management and related tree species on other sites and aspects in the Yellow River Delta.
Photosynthetic parameters are the main indicators reflecting the physiological processes underlying the response to stress in plants, and characterizing the photosynthetic response to soil water has facilitated the elucidation of the adaptability of plants to environmental changes(Jusovic et al. 2018; Liu et al. 2018; Zhang et al. 2020).Photosynthesis is not only determined by physiological functions and leaf structure dictated by the genetic characteristics of the species, but is also inf luenced by light,temperature, CO2concentration, soil water and other ecological factors (Carvajal et al. 2014; Ibrahim et al. 2017;Lockhart et al. 2017; Nama et al. 2019). Soil water is an important ecological factor af fecting plant physiological processes and plant distribution, which greatly impacts photosynthesis and water and energy use (Zhang et al.2010, 2012, 2020; Catoni and Gratani 2014). In arid regions, improving water use ef ficiency is a major adaptation strategy for the survival and growth of plants. These improvements are achieved through optimized stomata regulation (Farquhar and Sharkey 1982), the adjustment of leaf movement (Zhang et al. 2014) and other morphological and physiological pathways (Jan et al. 2019) to obtain a balance between carbon assimilation loss and water loss(Kosugi et al. 2013; Fernandes-Silva et al. 2016; Zerche et al. 2016). Based on limiting factor principles and ecological assessments, the demand on soil water in the process of photosynthesis should exist at dif ferent levels such that the photosynthetic characteristics of plants would be af fected when water content was too much or too little(Zhang et al. 2012; Xia et al. 2017). Related studies have shown that net photosynthesis, transpiration rate and water use ef ficiency did not exhibit linear relationships with soil moisture; conversely, a threshold ef fect on soil moisture was observed (Zhang et al. 2010, 2012; Xia et al. 2017).Studies concerning the physiological characteristics of drought resistance among dif ferent af forestation species have recently emerged (Zhang et al. 2012, 2014; Forni et al. 2017) evaluating anatomical structures, changes in physiological and biochemical factors and adaptations and mechanisms underlying water stress under dif ferent water stress levels (Jusovic et al. 2018; Liu et al. 2018;Zhang et al. 2018; Liang et al. 2020). However, studies concerning the physiological process of photosynthesis under dif ferent soil water conditions have primarily been restricted to a polyethylene glycol (PEG) habitat or a single pot simulation based on a few test designs, including mild, moderate, and severe water stress. In addition, little is known about the ecophysiological response of plants to soil water gradients, soil water photosynthetic productivity levels in actual habitats and the appropriate water conditions necessary for the growth of trees.
Sap f low parameters, which accurately ref lect transpiration water consumption characteristics in plants, play an important role in revealing the contribution of stem water storage to daily transpiration (Bo et al. 2017; Yu et al. 2019),the water use ef ficiency of tree species (Xia et al. 2014a, b;Kardile et al. 2019), the precise measurement of water consumption (Xia et al. 2014a, b; Toledo et al. 2020), and other measures. Current studies have primarily focused on the characteristics of sap f low, water consumption in dif ferent species from varying regions (Chen et al. 2014; Darby et al.2016; Bo et al. 2017 ), the relationship between environmental factors and the radial distribution pattern of sap f low(Bosch et al. 2014; Xie and Wan 2018), and the response characteristics of sap f low to environmental factors, including increases in temperature (Zhao et al. 2017), changes in rainfall patterns (Tang et al. 2018), and the enrichment of atmospheric carbon dioxide (Vongcharoen et al. 2018).However, studies on the response of sap f low to soil moisture changes from saturation to wilting point, and the response of sap f low and photosynthesis to changes in soil moisture are important to determine the moisture tolerance of plants.
In the present study to determine the response characteristics of sap f low and photosynthetic ef ficiency to soil water content, and to identify the water threshold that maintains photosynthetic capacity and water use ef ficiency, the soil water gradient obtained by irrigation and natural water consumption was used to measure and analyze stem sap f low,leaf photosynthetic ef ficiency and the critical point in a water gradient availability. These results could provide a practical basis for plantation management and high water use ef ficiency of poplar in the Yellow River Delta.
Materials and methods
Study material and design
The study area was located in the experimental area of Binzhou Chenier Island and Wetland National Nature Reserve (N 38°14′30′′, E 117°52′58′′), Wudi County, Binzhou City, Shandong Province, on the southwest shore of Bohai Bay. Twoyear-oldPopulus×euramericana‘Neva’ (poplar) saplings(from cuttings) with an average height of 1.6 ± 0.1 m were used for nine experimental plots (1 m length × 0.8 m width × 0.5 m depth) were dug in the Yellow River Delta. To avoid water inf iltration, an impermeable membrane was placed in the bottom of the plots and one per plot was planted April 1, 2019.One sample was randomly selected from each plot for testing,yielding a total of nine saplings. After cultivation for 70 days with artif icial watering and natural watering using dif ferent irrigation systems until June 12, 2019, the nine experimental plots were divided into three dif ferent groups based on the initial water gradient and the relative water content (Wr) of each plant: I (comparison group) (92%), II (75%), and III (63%).Each water gradient was repeated in triplicate and the soil surface covered with plastic f ilm to prevent evaporation, followed by the continuous reduction of the soil water by natural physiological water consumption to obtain soil water content intervals of 1–2 days, until net photosynthesis reached zero.The f inal measurements were on June 28, 2019. Using a soil drill, a 10–25 cm channel with 1.2 cm diameter was created in the main root distribution zone. Soil water was monitored using the drying method (Table 1) (Zhang et al. 2018), and f ield capacity (22.7% ± 1.1%) was determined using the cutting ring method at a soil bulk density of 1.3 ± 0.2 g cm−3(Zhang et al. 2018).
Determination of gas exchange parameters
Poplar leaf gas exchange parameters were measured using artif icial light sources from 9:00 to 11:00 a.m. on a sunny day.To reduce the dif ferences in the photosynthetic physiological criterion and changes resulting from time dynamics, measurements were obtained alternating between dif ferent duplicate leaves or dif ferent water gradients, to ensure the comparability and accuracy of the data among dif ferent treatments.For each soil water gradient, poplar saplings were randomly selected in each plot and three healthy, mature leaves were selected per sapling from the upper portion of the plant for in vivo measurements. Nine water gradients from high to low were selected, with gravimetric values of 21%, 18%, 17%,15%, 13%, 11%, 9%, 7% and 4% and relative values of 92%,81%, 74%, 64%, 56%, 47%, 39%, 29%, and 19%. A LI-6400XT portable photosynthesis system (Li-COR Inc., Lincoln, NE,USA) was used to measure the net photosynthetic rate (Pn) in response to light; artif icial light was used to control the photosynthetically active radiation (PAR) at 1600–20 μmol m−2s−1, with a total of 11 light intensity gradients. The control time for each gradient was 120 s, and CO2concentration was maintained at 370 μmol mol−1using CO2controller settings(Ca). Gas exchange parameters,PAR,Pn, transpiration rate (Tr),stomata conductance (Gs) and intercellular CO2concentration (Ci) were automatically recorded. Before the data were recorded, the leaves were maintained in a leaf chamber for 5 min until a steady state of photosynthesis was reached. In addition, similar to the method used to measure water content for group I with the general light saturation point ofPnin the saplings was 1100–1300 μmol m−2s−1(González-Rodríguez et al. 2017; Xia et al. 2017), gas exchange variables were measured using a f ixed saturation intensity with aPARof 1200 μmol m−2s−1for every water treatment in groups II and III. CO2concentration was controlled at 370 umol∙mol−1with a LICOR CO2injection system and a saturating photosynthetic photon f lux density of 1200 μmol m−2s−1was supplied from a LI-COR LED irradiation source. Air temperature of the leaf chamber was maintained at approximately 31 °C, relative humidity at 43%, and the f low rate of air was 200 μmol s−1.The instantaneous water use ef ficiency (WUE) =Pn/Tr, potential water use ef ficiency (WUEi) =Pn/Gs, and stomata limitation value (Ls) = 1 −Ci/Cain the leaves were also measured.
Sap f low measurements
The sap f low was continuously measured using the stem heat balance system (Dynamax Company, New York, NY, USA)while maintaining soil moisture. This system automatically and continuously measures sap f low velocity and diurnal cumulative sap f low f lux. Wraparound probes (SGA13) were installed at a height of 0.4 m from the bottom of the saplings,and f ive plants were measured. The probe was installed according to the manufacturer’s instructions, including selection and pretreatment of the stem and installation of sap f low probes,O-rings and radiation shielding. The cables of the probe for data transmission were connected with the appropriate interface on the data acquisition unit (Delta-T 80 Logger). Sap f low

Table 1 Physical characteristics of the shell sand soil
was continuously monitored for 30 min 24 h a day and data download every 2 days.
Data processing

For the three groups,PAR =1200 μmol m−2s−1was used to measure gas exchange specif ics in the soil water response characterization analysis. The rectangular hyperbolic correction model was used to simulate thePnlight response curve of group I leaves under dif ferent water gradients, and the simulation expression model (Ye 2007) was calculated as:wherePnis the net photosynthetic rate,PARthe light intensity, andαthe initial slope of the light response curve whenPAR =0, representing the apparent quantum yield (AQY);βandγare light photoinhibition and light saturation terms,respectively (Ye and Kang 2012), andLCPis the light compensation point. The light response parameters, including the quantum ef ficiency parameters (AQY, βandγ), light compensation point (LCP),LSP, maximum net photosynthetic rate (Pnmax), dark respiration rate (Rd), and other values were modeled using the nonlinear regression analysis through derivation conversion in SPSS 12.0 (Table 2). The dif ferences between the model parameters were analyzed using a one-way ANOVA (LSD) and a signif icance of dif ference level of 0.05.
Results
Light response characteristics of P n under dif ferent water conditions
ThePn(photosynthetic rate) was simulated using the rectangular hyperbolic correction model for poplar in the Yellow River Delta, and theR2model equation was 0.972 whenWr(relative soil water content) was 19%; the otherR2model equations were greater than 0.986. As shown in Fig. 1, an initial increase inPnwas observed, which subsequently decreased with increasingWr, reaching the highest level whenWrwas 74%. Photoinhibition was observed at high light intensity, withPAR> 1000 μmol∙m−2∙s−1at low waterWr< 40% and high waterWr> 80%; thePngradually changed at high light intensity under other water conditions.
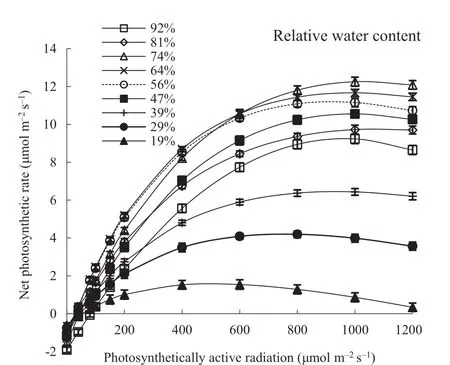
Fig. 1 Response curves for net photosynthetic rate ( P n ) f itted to the photosynthetically active radiation ( P AR ) in poplar leaves under varying relative water content; each point is the mean of at least three plants with f ifty-four replicate responses for each P AR . The lines are f itted to the response of P n to P AR under dif ferent soil water contents.Error bars represent the mean ± SE

Table 2 Light response parameters of the net photosynthetic rate in poplar leaves under varying soil water content
As shown in Table 2, whenWr <39%, theAQY(apparent quantum yield) in leaves was maintained at a low level with increasingWr. Thereafter, a signif icant increase inAQYwas observed with increasingWr, and the maximumAQYwas observed whenWrwas 64%. TheAQYsignif icantly decreased (P< 0.05) with a high waterWr >74%, indicating that the energy conversion in the leaves was reduced under low water or high water conditions, and the low-light utilization was subject to greater suppression. WhenW rincreased,LCP(light compensation point) decreased and subsequently increased, while theLSP(light saturation point) showed the opposite variation, i.e., in low waterWr< 30% and high waterWr >90%,LCPwas higher, andLSPwas lower, indicating that in this water range, the ability of poplar to use low light and strong light was diminished. These results suggest that poplar exhibits adaptation strategies under extreme water stress, showing a diminished use of light to compensate for adverse water conditions. The highestLSPand widest light ecological amplitude were observed. When theW rwas 74%, an improved ability to use strong light was demonstrated. ThePnmax(maximum net photosynthetic rate)increased with increasingWr: when theWrwas 74%, thePnmaxwas 7.8 times higher than whenWrwas 19%, suggesting that poplar has strong photosynthetic potential in high water environments. TheRdf luctuated with increasingWr, reaching a maximum when theWrwas 47%, suggesting increased respiration in the leaves upon the consumption of photosynthetic products under optimum water conditions.In high water conditions (Wr81%) and under severe drought stress (Wr< 23%), theRdwas low, indicating that leaf respiration was signif icantly reduced under water stress, and the consumption of photosynthetic products was inhibited under drought stress to maintain normal physiological activity conducive to dry matter accumulation. The photoinhibition (β) and light saturation (γ) terms showed contrasting variations with decreasingWr. When theWrwas ˂ 29% and92%, theβvalue was higher, indicating that poplar leaves are prone to photoinhibition under water stress corresponding to a lowerLSP. Under severe drought stress (Wr= 19%),a maximumγvalue was observed, indicating that the leaves easily achieved light saturation, and the ability to use strong light was reduced under drought stress.
Water response characteristics of P n and T r in poplar leaves


whereWris water content;Pnis photosynthetic rate. The averagePnwas 10.8 μ mol m−2s−1at aWrof 19–92%; this corresponded toWrvalues of 41% and 87%. Using a similar method, the averageTrwas 2.6 mmol∙m-2∙s-1, and this corresponded toWrvalues of 35% and 77%. The water content required to maintain photosynthesis and transpiration above the average level ranged from 41 to 87% and 35–77%,respectively.
Diurnal response characteristics of sap f low rates to soil water conditions
There was a signif icant dif ference between the sap f low rates with decreasing soil moisture (P< 0.01), and the daily average (38.6–140.0 g h−1) and daily peak (84.4–395.4 g·h-1)sap f low rates initially increased, followed by a decrease(Fig. 3). When theWrranged from 39 to 64%, the sap f low rate remained high with averages of 97.8–140.0 g h−1.Although the maximum sap f low rate (395.4 g h−1) was observed at aWrof 56%, the maximum average (140.0 g h−1)was observed at aWrof 64%. Under drought stress, when theWrranged from 47 to 19%, the sap f low rate decreased markedly, with an average rate from 38.6 to 107.9 g h−1,and the minimum average (38.6 g h−1) at aWrof 19%. In response to soil moisture, the sap f low rate did not reach the maximum value when the soil was near saturation. A high sap f low rate was maintained when theWrranged from 56 to 64%. Water consumption decreased under drought stress but the sap f low rate was signif icantly higher under high moisture levels (Wr> 74%) than under low moisture conditions (Wr< 29%), indicating that poplar sap f low rate was sensitive to soil moisture and showed a certain adaptability.

Fig. 2 Soil water response curves of a net photosynthetic rate ( P n ),transpiration rate ( T r ), b water use ef ficiency ( WUE), intrinsic water use ef ficiency ( WUEi), c stomatal conductance ( G s ) and d intercellular CO 2 concentration ( C i ) and stomatal limitation ( L s ) in poplar leaves; points represent the mean of a minimum of f ifty-four replicate P n , T r , WUE, WUEi, G s , C i and L s responses for each soil water content; vertical bars indicate the standard error of the mean

Fig. 3 Diurnal changes in sap f low rates of poplar under different soil moisture conditions
Diurnal changes showed that poplar sap f low rates signif icantly varied light and darkness and time, showing a higher value in daylight than at night, and midday depressions were observed atWrvalues of 81–92% and 29–47%. Under all soil water conditions, the sap f low rate was lowest between 19:00 pm and 5:30 am and gradually increased thereafter.The “peak platform”, consisting of many small peaks, was observed between 9:00 and 15:00, and appeared as a broad peak. Peak rate was observed at approximately 11:00–14:00 and gradually declined.tWith decreasing soil moisture, the diurnal variation range of stem sap f low initially increased and subsequently decreased, and the maximum range was observed at aWrof 56%. The range of diurnal variation decreased to 84.4 g h−1from 270.1 g h−1under drought stress, indicating that poplar leaves inhibit water loss to some extent and show resistance under drought stress.
Response of daily sap f low rate to soil moisture
As shown in Fig. 4, variations in daily cumulative sap f low were similar to the S-type under dif ferent soil moisture conditions, and there was a signif icant dif ference between values during the day and at night. Daily cumulative sap f low was low at midnight to- 6:00 am, rapidly increased from 8:30 to 16:00 showing a large slope, and remained stable with a low sap f low in the evenings. There were signif icant dif ferences (P< 0.01) in the daily sap f low under dif ferent soil moisture conditions, and it gradually increased with increasing relative soil water content (Wr). Daily sap f low reached the highest value of 1679.7 g d−1at aWrof 64%,followed by a decline with increasingWr, indicating that water consumption of poplar decreased under waterlogged conditions. Compared with the highest daily sap f low at aWrof 64%, signif icant dif ferences (P< 0.05) were observed atWrvalues of 92%, 81%, and 74%, followed by decreases of 63.2%, 63.2%, and 33.1%, respectively, and 30.1%, 64% and 72.4% under the low water conditions, respectively (atWrvalues of 39%, 29% and 19%). Thus the water consumption of poplar declines under low moisture conditions (Wr< 39%)with drought stress or high moisture conditions (Wr> 74%)with increased waterlogging. Notably, in the Yellow River Delta, soil moisture levels signif icantly af fected transpiration water consumption of poplar, and the reduction in sap f low under drought stress was more sensitive than under waterlogged stress.
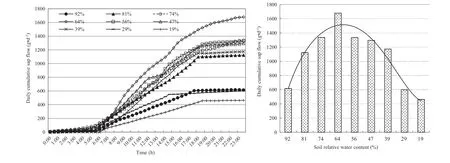
Fig. 4 Daily cumulative poplar sap f low under dif ferent soil water conditions
Water response characteristics of water use ef ficiency( WUE) in poplar leaves
TheWUEshowed an initial sharp increase and subsequently remained stable with increasingWrranging from 45 to 95% (Fig. 2 b), maintaining a high level and did not significantly change. The model for the response toWrwas:WUE =− 0.0015Wr 2+ 0.2122Wr− 2.0851, R2= 0.897.Based on this model, the highest level ofWUEwas 5.4 μmol∙ mmol-1whenWrwas 71%. According to the integral f itting equation:
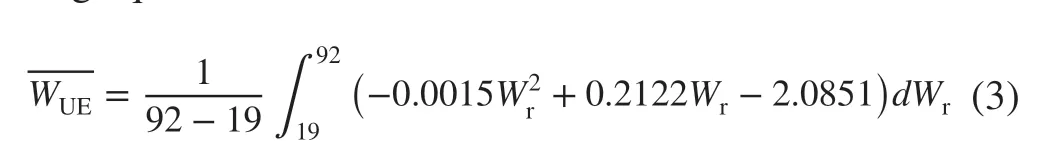
whereWris water content;WUEis water use ef ficiency. AverageWUEwas 4.4 μmol∙mmol−1on the alluvial plain in the lower Yellow River, corresponding toWrvalues of 45% and 96%, suggesting that the water content required to maintain poplar leafWUEat higher levels and exhibit the same validity ranged from 45 to 96%. In leaves under low water levels,Wr< 30%,WUEi(potential water use ef ficiency) signif icantly increased with increasingWr; when it was 35%, theWUEireached a higher value; when it was 50%, theWUEidecreased and subsequently increased; and when the relative soil water content was 78%, theWUEiwas high. Under highWr> 80%,theWUEisignif icantly decreased, indicating that the water use ef ficiency of poplar leaves decreases with both low and high water levels.
Water response characteristics of G s , C i and L s in poplar leaves
The decrease in leaf photosynthetic rates primarily ref lected variations of stomata and mesophyll cells, and based on the stomata limitation theory, this decrease was influenced through stomata or nonstomata factors (Farquhar and Sharkey 1982). Figure 2 c, d show that at 40% The soil water thresholds when photosynthetic rates (Pn)and water use ef ficiency (WUE) reached their highest, lowest and average values were indicative of soil water availability, and this is combined with the response ofPnandWUEtoWrto establish the photosynthetic ef ficiency water threshold classif ication based on the sizes ofPnandWUE(Table 2). The soil water critical ef fects ofPnandWUEin poplar leaves were adopted in this grading criteria, and physiological signif icance is shown; where photosynthetic rates were intended for production,WUEef ficiently replaced the concepts of production (biomass level) and ef ficiency(the degree of root hydrophilicity) in traditional agricultural studies. According to the light compensation and saturation points in thePnlight response curve, the point whenPnwas zero was referred to as the “Pnwater compensation point”.TheWrwas 13% and values lower than this was considered as non-productive and non-ef ficient water. WhenWrwas 42%, the main stomata limitation changed to the main nonstomata limitation, causing a decline in photosynthetic rates and this point was referred to the “Pnwater stomata limitation turning point”. WhenPnandWUEreached maximum values, referred to as the “Pnwater saturation point” and“WUEwater use ef ficiency point”,Wrvalues were 64% and 71%, respectively. Therefore, a range ofWrat 64–1% can be described as high productive and high ef ficiency water use.With high yield and high ef ficiency water use, referring to highPnandWUE, theWrwas determined by the “Pnwater saturation point” and the “WUEef ficient water point”. With average productivity and average ef ficiency water use, referring to above averagePnandWUEvalues, theWrwas determined by averaging two parameters based on the integral betweenPn,WUEand soil water. With low productivity and low ef ficiency water use, theWrwas determined by below averagePnandWUEvalues. Six photosynthetic ef ficiency water threshold classif ication types were determined: no productivity and no ef ficiency water use; low productivity and low ef ficiency water use; low productivity and middle ef ficiency water use; middle productivity and low ef ficiency water use; middle productivity and middle ef ficiency water use; and, high productivity and high ef ficiency water use. The photosynthetic potential, light energy utilization, photoinhibition levels and other characteristics of plants under stress could be ref lected through photosynthetic response parameters (Ye 2007; Xia et al. 2017; Toda et al. 2018),and changes in these parameters are closely associated with soil water content. With increasingWr, theAQY,LSP, LCPandPnmaxinitially increased and subsequently decreased,but there were differences between the critical points where peaks or valleys were observed for each indicator.The light response parameters of the shrubsHippophae rhamnoidesL. (Liu et al. 2017) andPrunus sibiricaL.,lianas such asCampsis radicansSeem. (Xia et al. 2017)and other plants showed similar variations. Under different water conditions on the alluvial plain of the lower Yellow River, the light compensation point (LCP) was 30.3–83.1 μmol m-2s−1, and the light saturation point (LSP)was 503.2–1138.3 μmol∙m−2∙s−1, with sun plant characteristics; the apparent quantum yield (AQY) was 0.01–0.04, close to the values 0.03–0.05 for optimum plant growth conditions. Under drought and waterlogged conditions, theLCPof poplar decreased to 30.7 μmol∙m−2∙s−1when theWrwas 64%, but theAQYreached its highest level at 0.04, indicating that poplar leaves had a better ability to utilize weak light.TheAQY,LSP,PnmaxandRdin the leaves were signif icantly higher under waterlogged conditions than under drought stress, and theLSPand light energy use ef ficiency reached their highest values, showing that the compensation ef fect of water to light intensity was signif icant when soil water was 74%. Maximum photosynthetic capacity occurred atWrof 74%, and aPnhigher than the water saturation point was observed when theWrwas 64%, as determined through simulations under saturation light intensity. These results show that there are some dif ferences between simulated and measured values. Under this water state, increased photosynthetic capability in poplar leaves might be associated with an increase inGs(stomatal conductance) and improvements in plant water status and inorganic ion absorption and transportation (Liu et al. 2018; Jan et al. 2019; Zhang et al.2020). These results suggest that light utilization capacity was enhanced under improved water conditions.Under drought conditions (Wr19% and 29%) and waterlogging (Wr92%), dif ferences in the photoinhibitionβwas high, indicating that poplar PSII antenna pigment molecules might have a larger photon absorption cross-section, or the average life expectancy of these molecules was longer in the excited state and photoinhibition was evident (Fig. 1).The longer average life expectancy of PSII antenna pigment molecules in the excited state exerts a protective ef fect on plants with a reducedPnand lower quantum advantage (Ye and Kang 2012). Under severe drought conditions,Wrvalues were 19%, and 29%, with a high light saturationγ, indicating that the leaves easily reached light saturation under severe water def iciency, and excessive absorption photons could be dissipated through f luorescence and heat consumption (Liu et al. 2017). These results indicate the adaptation characteristics of tolerating high light intensity as a survival strategy to protect the photosynthetic organs from damage under extreme drought conditions. Photosynthesis and the growth and development of plants are greatly af fected under water stress conditions, and studies by Zhang et al. ( 2014) and Xia et al. ( 2017) have shown that xerophytes and mesophytes under too little or too much water could easily exhibit reduced photosynthesis, ref lecting a block in the transportation of photosynthetic products and their accumulation in the leaves could lead to feedback inhibition forPn(Zhang et al. 2020). Photosynthesis is optimumin plants under suitable water conditions, such as in the desert saxaul (Haloxylon ammodendron(C.A. Mey.) Bunge),and the photosynthetic capacity of the leaves and population levels are strongest at aWrof 50%. Photosynthetic capacity is reduced in response to either increased or decreased soil water (Kosugi et al. 2013). Poplar leaf photosynthetic ef ficiency parameters showed a signif icant threshold ef fect to soil water levels in the Yellow River Delta, and the water conditions are key factors limiting stomatal changes. In range 40% The photosynthetic capacity in poplar leaves was strongest when soil water level was 64% but signif icantly lower than photosynthetic rates under drought or waterlogged stress (Fig. 1), and the poplar exhibited wet tolerant features.The maximumTrandWUEwere observed whenWrvalues were 56% and 71%, respectively, indicating that poplar leaves initially show strong transpiration with increasedWr,Gs, andPn, eventually obtaining their highest level. However, with increasingWr, normal metabolism of roots might be blocked (Zhang et al. 2013), or plant consumption might be limited to obtaining suf ficient carbon for the maximum amount of CO2assimilation with a limited loss of water,and optimal behavior was exhibited (Farquhar and Sharkey 1982). Stomata closure gradually reduces transpiration water loss and reduces the absorption of CO2to increase photosynthetic capacity; however, the rate-limiting process of photosynthesis is carboxylation (Zhang et al. 2020).Thus, under waterlogged conditions, poplar leaves exhibit reducedGs,TrandCito reduce water loss, while enhancing CO2assimilation, resulting in higherWUEandWUEilevels(Fig. 2). Poplar trees show a positive response strategy under waterlogged stress. This conclusion was not consistent with other studies, showing that in the early mild drought stages,water-def icient plants increase their water use ef ficiency to adapt to arid habitats (Zhang et al. 2010; Kosugi et al.2013), likely ref lecting the dif ferent synchronous responses ofPnandTrto soil water, resulting in a dif ferentiation of the poplar leafWUEto water stress.Pnwas more reduced than transpiration under drought stress in poplar (Fig. 2 a), resulting in lowerWUElevels (Fig. 2 b). The opposite ef fects were observed under waterlogged conditions, indicating that, in the Yellow River Delta, reductions in photosynthetic capability and water use ef ficiency in poplar were more associated with drought than waterlogging, indicating that drought tolerance of poplar is poor. Recent studies (Lang et al. 2013;Xia et al. 2017; Liu et al. 2017) have also shown that gas exchange parameters such asPn,TrandWUE, did not show a linear relationship with soil water, as these indicators were reduced when water levels are too high or too low. However,the dif ferences in plant species and habitat conditions contributed to greater dif ferences in the water critical point of photosynthetic ef ficiency parameters. In fact, the “Pnwater saturation point” (Wr64%) was higher in poplar leaves than inHaloxylon ammodendrondesert plants (Wr50%) (Kosugi et al. 2013), and this feature was lower than inPrunus sibiricain the loess hill area (Wr68%) (Xia et al. 2011), inWisteria sinensis(Wr71%) (Lang et al. 2013), inCampsis radicans(Wr71%) (Xia et al. 2017) in the rocky mountain area, inMalus pumila(Wr76%) (Zhang et al. 2010), and inPlatycladus orientalis(L.) Franco (Wr76%) andRobinia pseudoacaciaL. (Wr81%) (Catoni and Gratani 2014) on the semi-arid Loess Plateau. The “WUEwater point” (Wr71%) in poplar leaves was signif icantly higher than those inPrunus sibirica(Wr68%) (Xia et al. 2017),Malus pumila(Wr67%) (Zhang et al. 2010),Wisteria sinensis(Wr56%)(Lang et al. 2013),Robinia pseudoacacia(Wr64%) andPlatycladus orientalis(Wr52%) (Catoni and Gratani 2014),and this lower than inCampsis radicans(Wr85%) (Xia et al.2017). These results further indicate that poplar trees easily achieve photosynthetic capacity and water use ef ficiency in high water environments. Soil moisture was the main source of transpiration and sap f low was sensitive to soil moisture. The sap f low ofSchima superbaGardn under varying soil moisture levels during dry or rainy seasons was signif icantly dif ferent,and sap f low density declined signif icantly with decreasing soil moisture (Zhu and Zhao 2013). An increase of soil moisture signif icantly improved sap f low rates ofTamarix ramosissimaLcdcb (Zelelew et al. 2018) andAlbizia Kalkora(Roxb.) Pain. Simulated drought stress experiments also showed that precipitation was reduced by half,leading to a 41–44% decrease in stand transpiration, which further decreased to 80% under extreme drought conditions(Lapa et al. 2017). However, other studies have shown that drought stress does not lead to a reduction in the sap f low of some deciduous species but rather advances leaf senescence and leaf fall in the late growing season.A decline in sap f low only occurred when soil moisture was reduced to a particular level, and the moisture threshold and rate of sap f low decline were determined by the depth of the plant roots (Leuschner et al. 2019; Li et al. 2019). In the present study, the sap f low rate of poplar increased initially and subsequently fell with decreasing soil moisture. The highest transpiration and net photosynthetic rates did not occur under high moisture conditions (Fig. 2), potentially associated with oxygen limitation in the roots, moisture imbalance, stomatal closure, and increased CO2dif fusion resistance through brief waterlogged stress (Das and Das 2014). When soil moisture was suf ficient, transpiration was limited to some extent in the poplar, but the desertTamarix ramosissimamakes full use of water resources(Zelelew et al. 2018), suggesting that the poplar did not adapt to waterlogged soil. The sap flow was reduced with decreasing soil moisture under drought conditions,consistent with observations inSchima superbaandA.kalkora(Zhu and Zhao 2013; González-Rodríguez et al.2017), possibly ref lecting an increase in hydraulic resistance between woody components and soil, and the limitation of root water absorption through low soil hydraulic conductance (Huber et al. 2015). More than 99.8% of sap f low was used for transpiration and the daily cumulative f low directly ref lected the daily transpiration capacity (Dang et al. 2019). Daily water consumption ofHaloxylon ammodendronplants was signif icantly correlated with soil moisture, and the daily cumulative sap f low increased with increasing moisture (Cao et al. 2013). Daily transpiration showed a threshold to soil moisture. The daily transpiration was not the largest when water was suf ficient, but was the highest under suitable water conditions, and this factor was signif icantly reduced under severe drought. In poplar under dif ferent water conditions, daily cumulative sap f low exhibited an “S” shape,and dif ferences between day and night were signif icant, in contrast to the ef fects observed inHaloxylon ammodendronunder high soil moisture, but similar to the variation observed in these plants in drought conditions (Cao et al.2013), indicating that the consumption of water through transpiration was closely associated with soil moisture,but with a change of species and environmental factors,the variations in daily transpiration to soil moisture were signif icantly dif ferent. Diurnal changes in sap f low rates could ref lect the plant responses to water consumption, serving as a key parameter to determine the contribution of tree-stored water to transpiration (Yu et al. 2019). This study was carried out in the f ield environment where light, temperature, air relative humidity were constant. Therefore these were not considered and only soil moisture was used to analyze changes in sap f low. Notably, although diurnal changes in the sap f low rate might be ef fectively coordinated through soil moisture and the other environmental factors, a comprehensive measurement and analysis were not carried out. The diurnal changes primarily exhibited single, double and multimodal peaks,displaying high variation during the day and low variation at night (Barbeta et al. 2012; Cao et al. 2013; Zelelew et al.2018; Gholami and Zahedi 2020). It has been suggested that meteorological factors such as solar radiation, vapor pressure def icit, relative humidity, air temperature and wind speed, and soil moisture, may directly af fect diurnal changes in sap f low rates. The diurnal curve of sap f low rate of poplar in the Yellow River Delta varied over a 24-hour period, similar toZiziphus jujubaMill. andAlbizia kalkorain the rocky mountainous area of north China (González-Rodríguez et al.2017; Ceciliato et al. 2019) which showed a single peak of a particular width forming a “peak platform” from 9:00 to 15:00, consistent with the results obtained withHaloxylon ammodendronandA. kalkorain the dry, hot valley in Jinshajiang River (Cao et al. 2013; Ceciliato et al. 2019). The range of diurnal variation in the sap f low decreased under drought stress, suggesting an adaptation strategy when the lack of soil moisture af fects root water uptake, resulting in a marked decline in transpiration water consumption (Barbeta et al. 2012). Sap f lows ofQuercus palustris(Moon et al. 2016) andPinus elliottii(Will et al. 2001) in humid and subhumid regions were predominantly terminated in the evening,potentially associated with water stored in the trees (Zhu and Zhao 2013; Osada et al. 2014). However, weak f lows were observed by various shrubs in the arid desert at night under drought conditions (Barbeta et al. 2012; Gholami and Zahedi 2020). This is consistent with the conclusions in this study, ref lecting the weak f low resulting from water potential dif ferences and pressure formed by the roots, crown and leaves (Gholami and Zahedi 2020 ) or the darkness, drought,high temperatures and moderate wind speeds (Barbeta et al.2012; Zelelew et al. 2018). There are two major classes of soil water availability based on physiological indices. Class I adopts photosynthetic and transpiration rates, water use ef ficiency and solar energy utilization indices for the cluster analysis of a series of soil water points (typically 7–8) based on the average class size (Xia et al. 2017). However, there was a def iciency in the precise division of soil water ef fectiveness with increased randomness of the soil water point. Class II is a limit value solving method based on the quantitative relationship between the physiological indices and the soil water, where the soil water availability is divided by solving the low and high limits of photosynthetic physiological parameters. Using the Class II method, the low and upper limit values of soil water that maintain photosynthetic ef ficiency are determined, but a moderate level of photosynthetic ef ficiency is not quantitatively def ined. In the present study, an approach was introduced based on the Class II classif ication, and mathematical models were generated for the integral calculation of the mean of photosynthetic rate and water use ef ficiency, i.e., threshold values,based on the low and high limits and the medium that maintainedPnandWUElevels to obtain a water threshold grade and to evaluate the photosynthetic ef ficiency. Notably, this method can be described as critical value taxonomy. Using this approach, it was determined that poplar on the Yellow River Delta was not productive and inef ficient water use with soil water < 13%, low productivity and low ef ficiency water use at 13–41% with soil water > 96%, low productivity and average water use ef ficiency at 87–96%, average productivity and low water use ef ficiency at 41–45%, and average productivity and average water use ef ficiency at 45–64%and 71–87% (Table 3). In the soil water range of 64–71%(Table 3), high productivity and high ef ficiency water use was observed. Poplar had a high photosynthetic capacity and inef ficient water consumption, ref lecting inhibited transpiration and the ef ficient physiological water use in poplar leaves was ensured. Comparing the soil water range at a high rates of photosynthesis and water use ef ficiency with those of other plants, high productivity and high ef ficiency ranges were 46.9–74.5% forPrunus sibirica(Xia et al.2011), 49.5–71.1% forCampsis radicans(Xia et al. 2017),and 47.5–64.0% and 40.5–52.0% forRobinia pseudoacaciaandPlatycladus orientalis, respectively (Zhang et al. 2012).These results show that poplar resistance to drought is poor and the hybrid better adapted to wet habitats (Table 4). In summary, in the Yellow River Delta, too high or too low water levels reduce the photosynthetic ef ficiency in poplar,but with high soil water environments, these hybrids exhibit photosynthetic capacity and water use characteristics of high water levels. The soil water critical ef fects of Poplar photosynthetic ef ficiency are obvious, with high productivity and high ef ficiency water use ranging from 73% Table 3 Critical point of photosynthetic ef ficiency and its threshold grade in poplar Table 4 Results of grading criteria of soil water productivity and availability The diurnal changes in sap f low rates exhibited a single peak of a certain width, and the dif ferences between day and night f low rates were signif icant. Under insuf ficient moisture conditions in the Yellow River Delta, poplars could reduce daily transpiration time and the amount of water consumed through the delayed initiation and early termination of stem sap f low, indicating that poplars regulate water physiology and adaptability to drought stress.Graded water threshold of leaf photosynthetic ef ficiency
Discussion
Inf luence of soil water on photosynthetic light response
Critical ef fect of soil moisture on photosynthetic ef ficiency
Inf luence of soil moisture on transpiration water consumption
Impact of soil moisture on diurnal changes of sap f low rate
Grading and evaluation of photosynthetic ef ficiency related soil moisture
Conclusions

杂志排行

Journal of Forestry Research的其它文章
- Genome-wide identif ication and cold stress-induced expression analysis of the CBF gene family in Liriodendron chinense
- Decay rate of Larix gmelinii coarse woody debris on burned patches in the Greater Khingan Mountains
- Characterizing conservative and protective needs of the aridland forests of Sudan
- Point-cloud segmentation of individual trees in complex natural forest scenes based on a trunk-growth method
- Accuracy of common stem volume formulae using terrestrial photogrammetric point clouds: a case study with savanna trees in Benin
- Appropriate search techniques to estimate Weibull function parameters in a Pinus spp. plantation