中国淡水胭脂藻属植物形态及分子系统发育分析
2021-12-24南芳茹刘旭东吕俊平谢树莲
南芳茹 李 娟 刘旭东 冯 佳 刘 琪 吕俊平 谢树莲
(山西大学生命科学学院, 太原 030006)
胭脂藻属(HildenbrandiaNardo)隶属红藻门Rhodophyta, 真红藻纲Florideophyceae, 胭脂藻目Hildenbrandiales, 胭脂藻科Hildenbrandiaceae, 藻体桔红色或暗紫色, 软骨质状, 匍匐呈扁平皮壳状, 由基层细胞和直立细胞组成[1]。该属在海洋和淡水水体中均可生长, 二者形态特征相似, 但生殖方式不同, 胭脂藻海洋类群的生殖方式为四分孢子囊(Tetrasporangia), 淡水类群的生殖方式为藻体的断裂和孢芽(Gemmae)[2]。胭脂藻属海洋类群在潮间带和潮下带区域广泛分布[3,4], 而淡水产胭脂藻对生境条件要求苛刻, 地理分布十分有限, 已有国家将其列为濒危物种[5,6]。该属植物形态特征较为简单,但却是淡水红藻中起源较早、进化地位较为高级的类群之一。
胭脂藻属(Hildenbrandia)最初由纳尔多(Nardo)于1834年在意大利发现并建立, 此后陆续报道了该属在北美洲、亚洲和欧洲等地的分布[7,8]。目前Algaebase已收录的分类单位近50个, 只包含了6个淡水种类, 分别为河生胭脂藻Hildenbrandia rivularis(Liebmann) J. Agardh、Hildenbrandia angolensisWelwitsch ex West & G. S. West、Hildenbrandia arracanaZeller、Hildenbrandia ramanaginaeM.Khan、Hildenbrandia jigongshanensisNan & Xie和Hildenbrandia japananensisF. Nanet al.[9]。根据传统的分类标准, 淡水胭脂藻种类的鉴定依据包括细胞大小、藻体壳状皮层的直径和藻体厚度, 其中细胞大小和藻体高度是2个主要的分类特征, 而这些形态特征在不同的环境条件下是可变的, 受环境因素的影响较大, 因而根据传统的形态学分类方法对淡水胭脂藻进行种类分类鉴定十分困难[10,11]。随着近年来分子生物学方法在淡水红藻分类和系统演化研究中的广泛应用, 在胭脂藻属的分类和起源研究中也加入了越来越多的分子证据。基于rbcL、18S rDNA等分子数据的支持, 建立了2个新种H. jigongshanensis和H. japananensis[12,13]。基于从北美洲采集的标本, 利用rbcL和18S rDNA序列分析, 发现淡水种H. rivularis为单系类群, 而H. angolensis在系统树中呈多系分布; 同时淡水胭脂藻与海洋类群呈多系关系, 推测淡水红藻是由海洋种类向内陆多次入侵而形成的[10,14]; 但基于从欧洲分布的胭脂藻标本构建的系统发育树, 显示淡水种是经过单次的海水种入侵形成的[15]。由此可见, 海产、淡水产胭脂藻种群的关系依然不明确, 需要更多分子数据进行深入研究。
淡水胭脂藻的地理分布具有一定的区域特征,欧洲分部的淡水种主要是H. rivularis[10], 北美洲分布的淡水种主要是H. angolensis[14]。目前H. ramanaginaii为印度特有种[16], 而H. arracana只报道于缅甸[17]。2003年, Sherwood和Sheath综合形态和分子系统发育构建, 对采集到的48个胭脂藻属标本进行分析, 根据形态特征归类, 将淡水胭脂藻种类分为2组, 分别对应北美、菲律宾分布和欧洲、卡纳里分布的样本[11]。另外2个淡水胭脂藻种类H. jigongshanensis为中国特有种,H. japananensis只分布于模式标本产地[12,13]。目前中国报道的淡水胭脂藻有2种, 河生胭脂藻H. rivularis分布地有山西、福建、湖北和西藏[1]; 鸡公山胭脂藻H. jigongshanensis分布地有山西和河南[12,13]。需要借助分子数据深入研究淡水胭脂藻的现代地理分布格局和祖先地理起源。
本研究基于从中国云南和河南新采集的2株胭脂藻标本, 对其进行了形态特征测量和目标基因序列测定, 综合传统形态特征和分子生物学证据对其进行了种类鉴定和系统发育构建, 同时结合目前已报道的胭脂藻地理分布信息, 对该属植物进行了地理亲缘关系和系统发生分析, 丰富了我国淡水胭脂藻的多样性, 同时为胭脂藻属植物的起源和进化提供了理论基础。
1 材料与方法
1.1 实验材料
本实验使用的胭脂藻藻株(编号: YZYUNNAN)于2019年4月16日采自云南省玉溪市新平彝族傣族自治县曼卡路 (24.07N, 101.99E), 藻株编号YZHENAN于2019年8月28日采自河南省焦作市修武县云台山不老泉(35.47N, 113.34E)。使用刀片将附着在石头表面的胭脂藻轻轻刮下, 再用无菌水将附着的杂质清洗后, 一部分立即进行分子鉴定, 另一部分用4%的甲醛溶液进行固定后用作形态学观察。
1.2 实验方法
形态学观察使用显微镜(BX-51, 奥林匹斯)对藻体进行观察, 并用其附带的相机(DP72, 奥林匹斯, 东京, 日本)进行图像信息的采集。
总DNA的提取利用Saunders提出的DNA提取方法的改良版对采集的胭脂藻进行总DNA的提取[18]。
扩增目的基因及测序查阅文献, 确定PCR体系及引物, YZHENAN藻株rbcL基因引物为HILF2和rbcLr, UPA基因引物为p23SrV-f1和p23SrV-r1; YZYUNNAN藻株rbcL引物为F650和rbcLr;psbA引物为psbAF和psbAR2[13,19,20]。PCR体系为20 μL: DNA模板0.5 μL, dNTPs 2 mL, 正向、反向引物各2 μL,TaqDNA聚合酶0.2 μL, 10×Buffer(Mg2+) 2 μL, 补充ddH2O至20 μL。扩增上述基因得到PCR产物后, 通过1%琼脂糖凝胶电泳(120 V,20min)进行检测。扩增产物送至华大基因公司进行检测: 采用Sanger双脱氧终止法进行双向测序(华大基因公司, 北京)。
系统发育树构建在NCBI中运用blast工具将实验中获得的rbcL、psbA和UPA基因序列与GenBank已发表的有关序列进行在线比对后筛选出所需的基因序列。利用软件BioEdit对所选序列进行比对并切除两端不整齐的碱基[21], 在软件MAGA7.0中对下载的序列及目的序列进行序列特征分析[22], 利用Modeltest ver. 3.7软件计算分子序列集最优进化模型[23]。分别使用MEGA7.0软件构建邻接树, PHYML软件构建最大似然树, MrBayes ver. 3.1.2软件构建贝叶斯系统发育树[22,24,25], 基于1000次重复计算得到邻接树和最大似然树各节点的支持率, 贝叶斯后验概率基于1000代抽样并舍弃前25%计算获得。在建树结束后, 使用Figtree 1.4.2对各软件的计算结果进行查看并编辑 (http://tree.bio.ed.ac.uk/software/figtree/)。
地理起源分析GenBank数据库中已有的胭脂藻属各地理分布的标本中, 只有rbcL序列信息比较充足, 选取该序列进行生物地理系统分析。对GenBank中获得的rbcL序列的标本分布地进行统计, 按地理位置A-西太平洋东南亚岛国、B-拉丁美洲、C-东加勒比海、D-欧洲、E-北美洲和F-亚洲进行分类, 结合贝叶斯系统树结果, 利用RASP软件构建近源物种祖先地理分布图[26]。
2 结果
2.1 形态观察结果
采自河南焦作市的胭脂藻标本(YZHENAN)和采自云南玉溪市的胭脂藻标本(YZYUNNAN)形态观察结果分别为图 1和图 2。藻体外形为薄皮壳状,颜色呈紫红色, 附着在水流中的石块表面生长。靠近藻体的中部颜色较深, 为深紫红色, 藻体边缘区域颜色逐渐变淡。基部细胞单层, 直立细胞多层,形成整齐致密的分枝。藻体高度分别为42—50 μm,细胞直径分别为(5.0—8.0) μm×(4.0—6.7) μm和(4.8—8.0) μm×(4.8—6.4) μm, 细胞近四方形, 片状色素体周生。
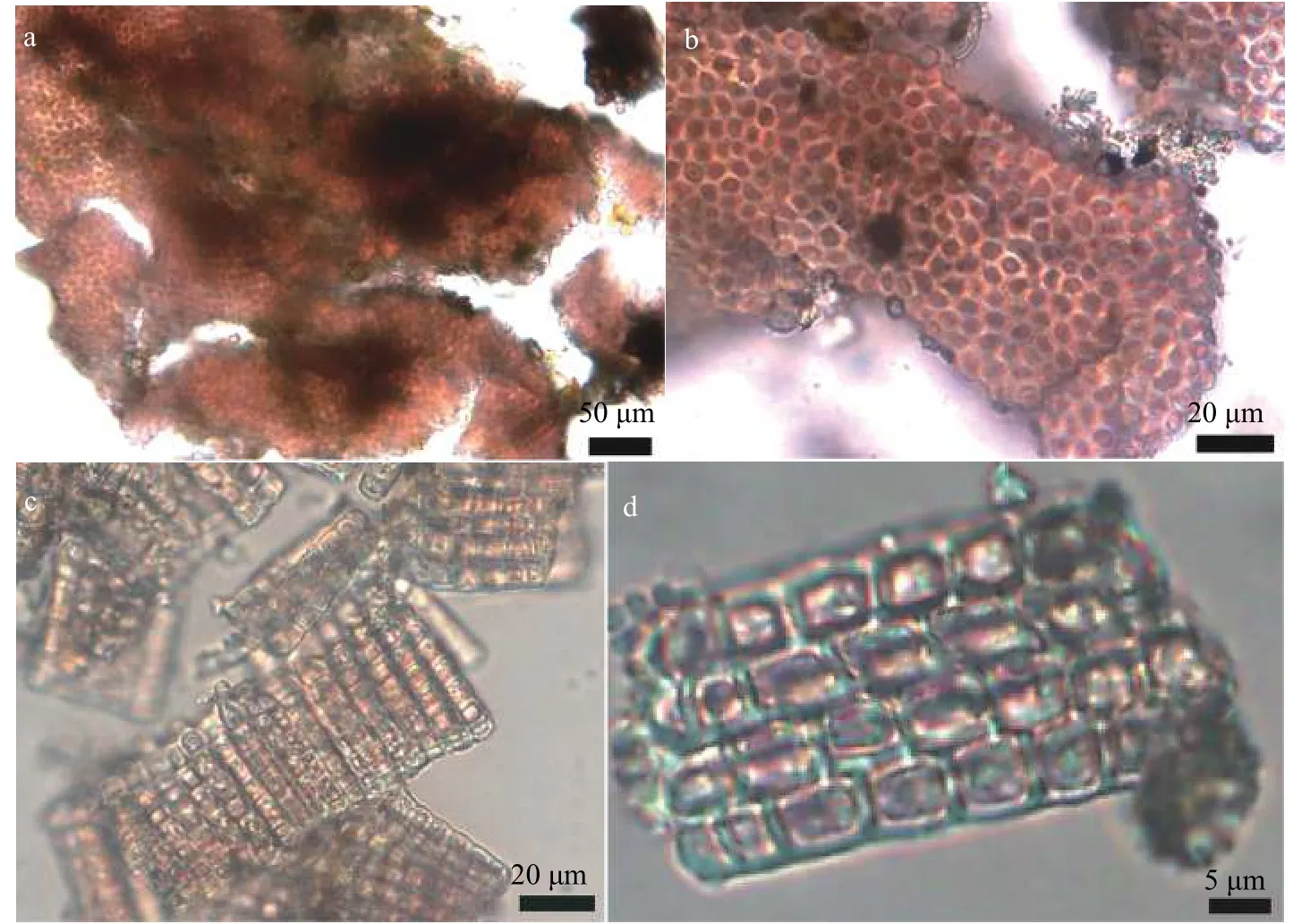
图1 采自河南焦作市的胭脂藻标本(YZHENAN)形态显微照片Fig. 1 Morphological structures of specimen collected from Jiaozuo City, Henan Province, China (YZHENAN)
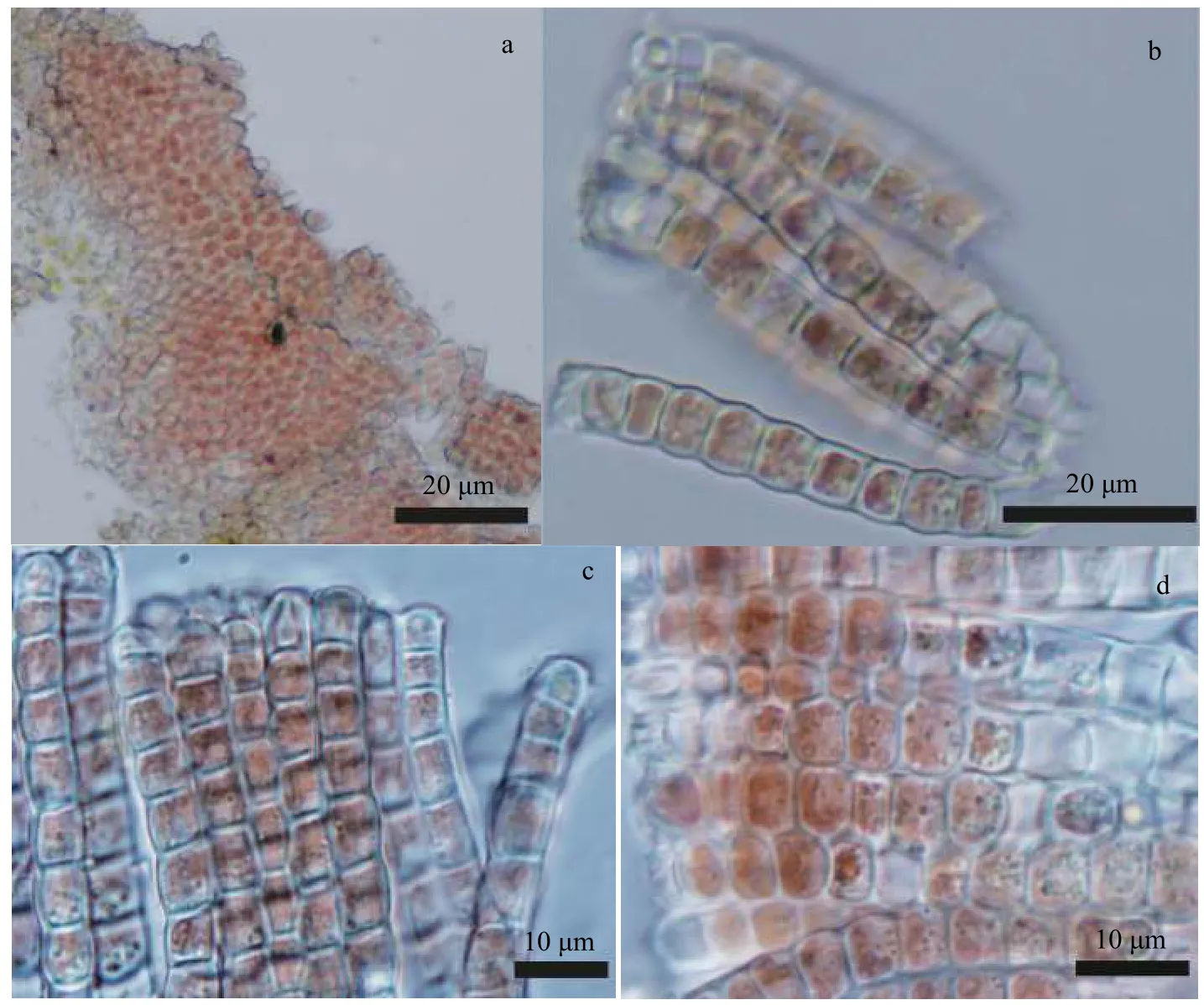
图2 采自云南玉溪的胭脂藻标本(YZYUNNAN)形态显微照片Fig. 2 Morphological structures of specimen collected from Yuxi City, Yunnan Province, China (YZYUNNAN)
2.2 系统发育分析
序列特征分析所采集的标本所测序列已上传至GenBank数据库中。基于rbcL序列计算结果, 所采标本YZHENAN、YZYUNNAN与H. jigongshanensis其他标本的遗传距离为0.001, 与其他淡水胭脂藻种类的遗传距离为0.005—0.127(与H.japananense的遗传距离为0.005, 与H. rivularis的遗传距离为0.033, 与H. angolensis的遗传距离为0.127); 本研究测得一株采自云南的胭脂藻标本(YZYUNNAN)psbA序列, 基于psbA序列计算结果,该标本与另外一株H. jigongshanensis标本的遗传距离为0.001, 与H. rivularis的遗传距离为0.086; 基于本研究中测得的采自河南的胭脂藻标本(YZHENAN)的UPA序列, 所采标本YZHENAN与另外1株H.jigongshanensis标本的遗传距离为0.023, 与H. angolensis的遗传距离为0.082, 与H. rivularis的遗传距离为0.080。各序列经过Modeltest软件分析得到的最佳进化模型及相关的模型参数列于表 1。

表1 Modeltest 3.7分析获得的rbcL、psbA和UPA基因模型及相关参数Tab. 1 Nucleotide substitution model parameter estimates of rbcL, psbA and UPA gene for Modeltest 3.7 analyses
基于rbcL、psbA和UPA基因的系统进化分析基于rbcL基因数据集, 以采集到的2株胭脂藻YZHENAN和YZYUNNAN, 以及GenBank下载的胭脂藻科的28株胭脂藻标本作为研究对象, 以红索藻属Thorea为外类群, 采用邻接法, 最大似然法和贝叶斯法三种方法构建得到的系统发育树拓扑结构一致, 因此将最大似然法得到的系统树作为基准, 把3种方法所得到的支持率标注在对应的节点,得到最终的系统发育树。基于rbcL基因的系统发育树显示(图 3), 本研究中的标本YZHENAN、YZYUNNAN与其他5株鸡公山胭脂藻H. jigongshanensis标本聚为一支, 支持率为73.9/0.76/85, 日本胭脂藻H. japananense位于鸡公山胭脂藻形成的分支基部。河生胭脂藻H. rivularis所有标本聚为一个单系分支, 支持率为70.5/0.75/88。鸡公山胭脂藻H. jigongshanensis、日本胭脂藻H. japananense和河生胭脂藻H. rivularis聚类为一个大分支, 但支持率较低, 采自哥斯达黎加的2株安哥拉胭脂藻H. angolensis标本(AF107816和AF107817)形成的姐妹支位于该分支基部, 得到了很高的支持率(98.9/0.97/100)。安哥拉胭脂藻H. angolensis在系统树中形成多系分布, 但都位于其他3种淡水胭脂藻的基部位置, 淡水胭脂藻全部标本, 包括4个种类(鸡公山胭脂藻H. jigongshanensis、日本胭脂藻H. japananense、河生胭脂藻H. rivularis和安哥拉胭脂藻H. angolensis)形成一个单系分支, 但只得到了贝叶斯方法的支持度为90%。海水胭脂藻H. rubra和H. crouanii位于整个系统树的基部位置, 其中H. rubra形成多个分支。
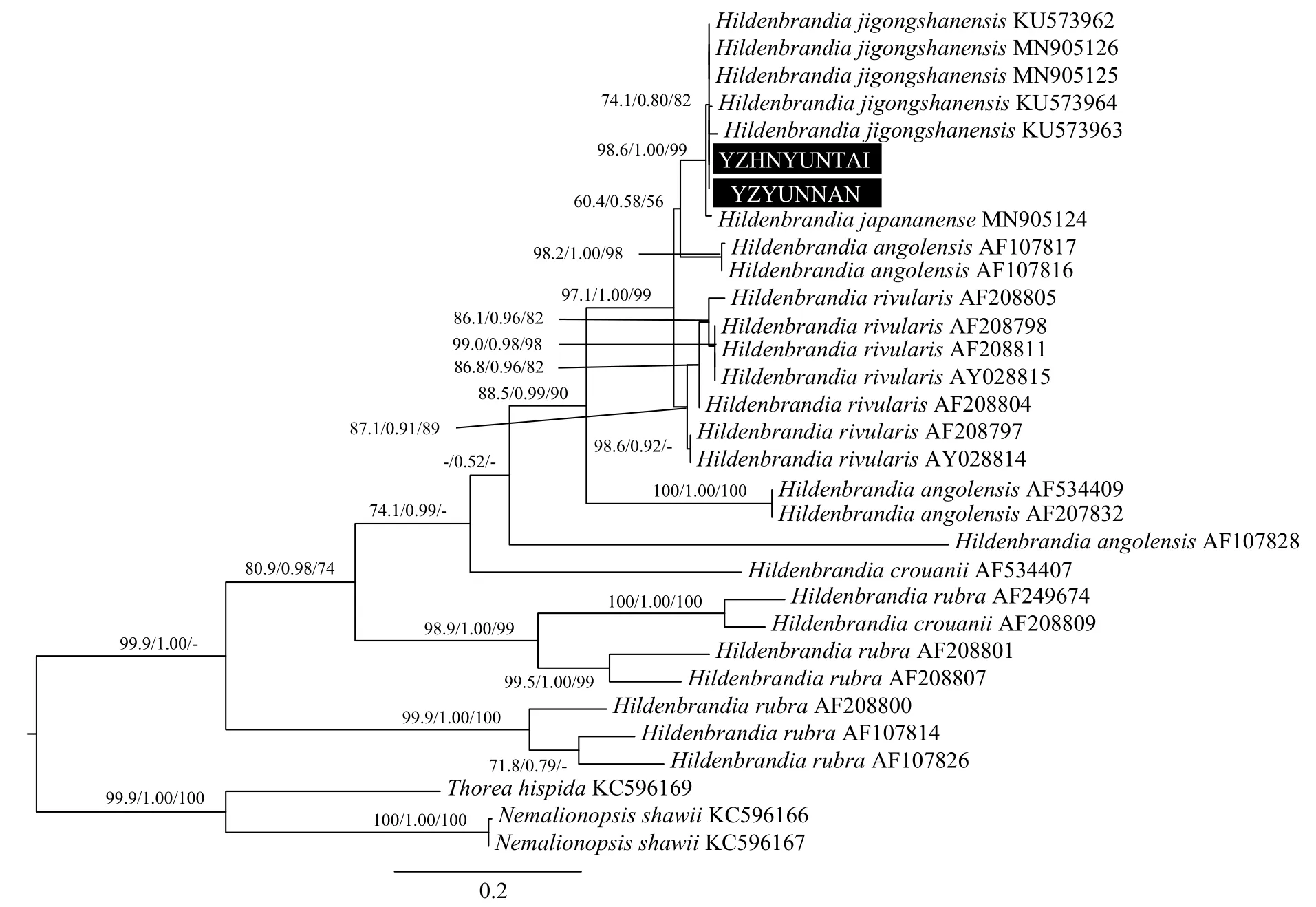
图3 基于rbcL基因序列的系统树Fig. 3 Phylogenetic tree based on rbcL gene sequence
基于psbA基因的系统发育树如图 4所示, Gen-Bank数据库中关于胭脂藻该序列的信息较少, 只有1株淡水标本(KX284723)和1株海水标本(DQ787633)的序列信息。本研究测得一株标本YZYUNNAN的psbA基因序列, 其与采自河南鸡公山的鸡公山胭脂藻H. jigongshanensis标本聚类形成姐妹支, 并得到了很高的支持率, 3种方法的支持率均为100%。河生胭脂藻H. rivularis与鸡公山胭脂藻H. jigongshanensis分支聚类, 也得到了较高的支持率(93.7/1.00/98)。海水胭脂藻H. rubra与胭脂藻科的另一个属Apoph-laea聚为一支, 支持率为90.2/1.00/65。基于UPA基因的系统发育树如图 5所示, 目前胭脂藻属在Gen-Bank数据库中有3株淡水标本(EF426600、KM 676587和KX284723)和4株海水标本(HQ421668、HQ421389、HQ421105和KX284724)的序列信息,本研究所测标本YZHENAN与鸡公山胭脂藻H. jigongshanensis聚为姐妹支, 3种方法的支持率均为100%。鸡公山胭脂藻H. jigongshanensis分支与2株安哥拉胭脂藻H. angolensis标本形成的姐妹支聚类,基部为河生胭脂藻H. rivularis, 共同形成一个单系分支。4株海水胭脂藻H. rubra标本与胭脂藻科的另一个属Apophlaea聚类。以红藻属Bangiopsis为外类群, 3株紫球藻标本形成的分枝位于系统树基部, 其他胭脂藻科的所有类群聚为一个单系分支,得到了3种方法100%的支持率。
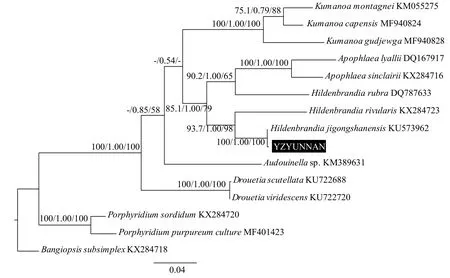
图4 基于psbA基因序列的系统树Fig. 4 Phylogenetic tree based on psbA gene sequence
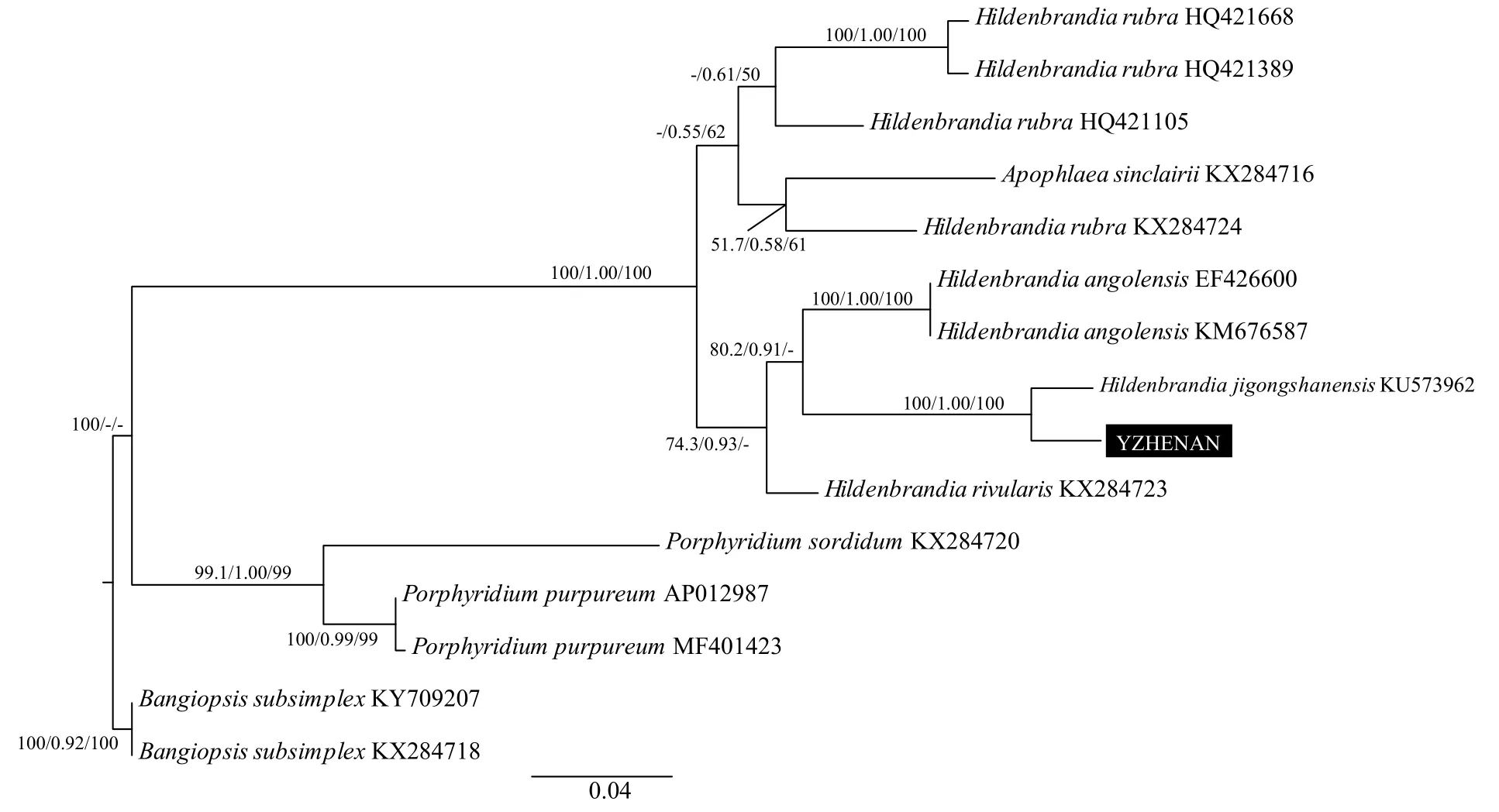
图5 基于UPA基因序列的系统树Fig. 5 Phylogenetic tree based on UPA gene sequence
基于可靠的系统发育关系和各个标本的现代地理分布信息, 得到胭脂藻属植物的祖先地理分布图(图 6)。目前所有鸡公山胭脂藻H. jigongshanensis标本的分布地均为中国,H. japananense的分布地仅在日本, 河生胭脂藻H. rivularis的分布地主要在欧洲, 安哥拉胭脂藻H. angolensis的分布地比较广泛, 包括西太平洋东南亚岛国、拉丁美洲和东加勒比海区域和北美洲。通过祖先地理分布区重建,淡水胭脂藻的祖先地理分布地追溯至北美洲(节点59), 且得到100%的支持。鸡公山胭脂藻H. jigongshanensis各标本的聚类关系中, 采自河南的4株标本(KU573962、KU573963、KU573964和YZHENAN)聚为一枝, 而采自云南的YZYUNNAN与采自山西的MN905125聚为一枝, 采自山西的另一株标本MN905126形成独立的分枝。
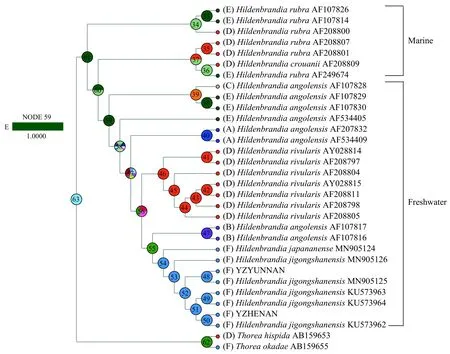
图6 基于rbcL序列的胭脂藻属祖先地理分布图Fig. 6 Reconstruction of ancestral geographical distribution of genus Hildenbrandia based on rbcL gene sequence
3 讨论
rbcL序列已广泛用于淡水胭脂藻属的分子生物学研究中, 而psbA和UPA序列也在其他淡水红藻的系统分类研究中比较常见, 它们都包含有较多的系统发育信号, 能够被用于进行种类鉴定和系统发生分析[13,27,28]。基于rbcL序列的计算结果, 河生胭脂藻H. rivularis的种间遗传距离为0—1.9%[10]。而本研究中YZHENAN、YZYUNNAN与H. jigongshanensis的rbcL序列遗传距离为0.1%, 介于种间遗传差异范围内, 该差异也远小于YZHENAN、YZYUNNAN与其他淡水胭脂藻种类的遗传距离,同时基于3个基因序列的系统发育树也表明,YZHENAN、YZYUNNAN与鸡公山胭脂藻H. jigongshanensis的标本聚合为一个独立分枝, 并得到了很高的支持率, 根据分子生物学证据, 可以将YZHENAN和YZYUNNAN定种为鸡公山胭脂藻H.jigongshanensis。目前鸡公山胭脂藻H. jigongshanensis为中国特有种, 其模式标本产地为河南省信阳市, 之后在山西省平定县也报道了该种的分布[12,13],本研究中该种类的标本采集自云南省玉溪市和河南省焦作市, 拓宽了该中国特有种的地理分布范围。
基于传统形态特征和现代分子生物学证据, 胭脂藻属目前被接受的4个淡水种类的关键分类特征列于表 2, 其中日本胭脂藻与鸡公山胭脂藻H. jigongshanensis的形态特征参数重叠, 主要依据分子生物学证据建立该种[13]。本研究中所采标本YZHENAN和YZYUNNAN藻体高度为42—50 μm,细胞直径分别为(5.0—8.0)μm×(4.0—6.7)μm和(4.8—8.0)μm×(4.8—6.4)μm, 与河生胭脂藻的形态参数是相吻合的, 但分子系统发育分析显示它们之间存在很大的遗传差异, 包含YZHENAN和YZYUNNAN的鸡公山胭脂藻与河生胭脂藻各自形成独立的分枝, 本研究对YZHENAN和YZYUNNAN的定种主要依据分子生物学证据, 与传统的形态特征分类存在不一致。随着分子系统发育分析的普遍应用, 淡水红藻中利用分子生物学证据进行种类鉴定, 从而否定了一些传统形态分类特征的可靠性[20,29,30]。通过本研究中所采集2株鸡公山胭脂藻的形态特征比较, 发现传统的营养结构形态分类特征包括藻体高度和细胞直径, 在不同的环境条件下是可变的, 不适合作为胭脂藻属植物种类鉴定的依据。对于胭脂藻属这一形态结构较为简单的类群, 借助分子生物学证据和系统发育分析手段对其进行种类鉴定和亲缘关系重建是必要的。

表2 淡水胭脂藻种类的关键分类特征Tab. 2 Key taxonomic characters of freshwater Hildenbrandia
淡水胭脂藻的起源和系统演化研究方面, Sherwood和Sheath基于rbcL序列对北美洲和欧洲的胭脂藻属植物进行了系统发育分析, 表明淡水胭脂藻种类H. rivularis形成一个单系分枝, 与海洋种类相分离, 且种间遗传距离很小, 认为其是海洋种的单次入侵形成; 而H. angolensis与海水种类呈多系聚类的关系, 认为淡水的胭脂藻类群是经过海洋种多次入侵形成的[11,14,15]。根据本研究中胭脂藻属植物分子系统发育结果, 支持河生胭脂藻H. rivularis是一个独立的单系类群, 安哥拉胭脂藻H. angolensis是多系起源, 而鸡公山胭脂藻H. jigongshanensis也是一个独立的单系类群, 其与分布于日本的日本胭脂藻H. japananense亲缘关系较近, 与其他2个淡水种H. rivularis和H. angolensis形成一个独立的分枝, 但与海洋种类的遗传差异较大。基于3个基因的系统发育关系, 都表明淡水胭脂藻种类形成一个独立的聚类群, 与海洋种类遗传距离较远,推测该类群是在内陆水体中独立起源和演化形成的。与之前淡水胭脂藻通过海洋近缘种入侵内陆水体的起源推测不同, 本研究通过增加胭脂藻属植物样本取样分析, 佐证了之前基于细胞器基因组系统演化的分析结果[11,14,31,32]。胭脂藻淡水与海洋类群营养细胞形态相似, 主要区别是生殖方式不同[2],这与红藻门另一个属红毛菜属Bangia是相似的, 但在红毛菜分类体系中仍将海洋和淡水类群归为同一属[33], 因此我们建议依然保留胭脂藻属现在的分类体系, 将海洋和淡水类群的遗传距离归属为种间差异。
淡水胭脂藻具有明显的地理分布特点, 根据报道,欧洲分布的淡水胭脂藻种类为H. rivularis, 北美的淡水胭脂藻种类主要为H. angolensis,H. jigongshanensis和H. japananense为中国和日本特有种[11,12—14]。胭脂藻目属于红藻纲中起源时间最早的一个谱系,在大约943百万年前发生分化, 经历了较长的演化过程[34], 这可能是该属植物各地理区域演化形成不同种类的原因之一。祖先地理重建的分析结果表明, 淡水胭脂藻的祖先起源地位于北美洲, 然后逐渐向东加勒比海区域、拉丁美洲和欧亚大陆扩散,经过种群扩散和各地理分布区类群的独立演化后,形成现代的种类地理分布模式。淡水胭脂藻在北美洲区域起源时间较早, 进化历史漫长, 遗传多样性大, 这与该地理区域分布的种类H. angolensis为多系类群是相一致的。淡水红藻的典型代表类群串珠藻目Batrachospermales, 其祖先起源地位于古地中海沿岸, 大洋洲和北美洲是两个次级分布中心[35]。在红藻的现代分类系统中, 胭脂藻属隶属于胭脂藻亚纲Hildenbrandiophycidae, 与串珠藻目植物隶属2个不同的亚纲分类单元, 亲缘关系较远, 且属于红藻中起源比较古老的谱系[34], 因此二者的地理起源和分布扩散模型不同, 也进一步表明了淡水红藻的古老起源和进化多样性。
