夜行性昆虫微光视觉行为及其视觉适应机制
2021-12-23杨小凡路子云苑士涛李建成魏国树
杨小凡, 路子云, 苑士涛, 李建成,*, 魏国树,*
(1. 河北省农林科学院植物保护研究所, 河北省农业有害生物综合防治工程技术研究中心,农业部华北北部作物有害生物综合治理重点实验室, 河北保定 071000;2. 河北农业大学植物保护学院, 河北保定 071000)
自1891年奥地利生理学家Exner首次提出昆虫可以看到和人类相似的图像以来,昆虫视觉一直受到广泛关注(Land, 1980)。视觉作为昆虫最重要的通讯手段之一,在取食、觅偶、产卵、越冬、迁徙、回巢和避敌等行为活动中都起着重要作用。大量研究表明,昆虫的视觉非常灵敏,通过复眼等视觉器官可以准确感知外界环境中的颜色、亮度、形状、大小、对比度、偏振光和运动状态等物理特征,以做出相应的趋向或回避反应(Land, 1997; Borst, 2009; 刘军和和赵紫华, 2017)。例如,在树叶颜色变红的秋季,黄色和绿色叶片对蚜虫更具吸引力(Döringetal., 2009)。蝴蝶Papilioaegeus利用植物反射的偏振光选择合适的产卵场所,对水平偏振光选择率显著高于垂直偏振光(Kelber, 1999)。目前,有关昆虫视觉行为的研究主要局限于日行性昆虫。由于夜行性昆虫主要在光线昏暗的夜间或黄昏活动(光照强度仅为白天太阳光的1/106~1/108),周围环境中的光或色等视觉线索不可能如白天一样清晰可见(Warrant, 2004),因此,普遍认为夜行性昆虫的复眼或单眼发生了退化或功能丧失,主要依靠嗅觉和机械性感受等来探索环境(Balkeniusetal., 2006)。
早在20世纪60年代,国内外学者开始观察研究夜行性昆虫的视觉功能,但主要集中在其复眼组织结构、光学原理和光敏感性等方面,尽管基于夜行性昆虫对特定光谱和光强的趋避性研发的灯光诱捕技术已在农林害虫的监测和防控上广泛应用,但对其视觉辨别能力尚不清晰(陈元光和钦俊德, 1963; 丁岩钦等, 1974; Anton-Erxleben and Langer, 1988; Cutleretal., 1995; 魏国树等, 2000)。直至21世纪初,瑞典隆德大学Eric Warrant教授及其研究团队通过红外夜视、视网膜电位(electroretinogram, ERG)和视觉神经等观测手段和试验技术陆续发现蛾类、蜜蜂和蜣螂等夜行性昆虫具有卓越的夜视能力,夜间或低光照强度下清晰、准确地获取颜色、大小、偏振光和运动状态等有效的视觉信息,产生和日行性昆虫相似的视觉行为活动(Kelberetal., 2002; Dackeetal., 2003; Somanathanetal., 2008; Sponbergetal., 2015; Dreyeretal., 2018)。这些研究结果挑战了夜行性昆虫视觉退化的传统经典理论,开启了微光视觉(dim-light vision)这一全新的研究领域,推动了夜行性昆虫的视觉研究由单一的趋光行为扩展到了视觉生态学的各个方面,但目前这部分的研究工作尚十分薄弱。为此,本文结合最新的研究进展,重点阐述了微光视觉的概念与特性、夜行性昆虫微光视觉行为反应及其视觉适应机制,以期为推进夜行性昆虫视觉生态学研究及其在害虫防治中的应用提供参考。
1 微光视觉的概念与特性
微光视觉的概念起源于20世纪初人类视觉的两重功能学说,即明视觉(photopic vision)和暗视觉(scotopic vision),其中,暗视觉强调视网膜上的视杆细胞对光的敏感度较高,能够在夜间光环境中感受微光刺激而引起视觉反应,但只能辨别目标物体的明暗,无颜色视觉(Newell, 1963)。与明亮的白天不同,夜间微光环境主要由月光、星光、大气辉光以及太阳光、月光和星光的散射光综合形成。当太阳高度角低至地平线下6°时,民用曙暮光结束,天空正式进入微光模式,此时周围环境的光照强度发生剧烈变化,由白天太阳光强度的100 000 lx下降到满月月光的0.1~0.3 lx(Croninetal., 2014)。无月时,星光和大气辉光在地球表面上所产生的光照强度仅约为0.001 lx,阴天星空的光照强度甚至下降至0.0001 lx,相当于满月时的1/103左右(图1)。
相比于人类在微光条件下只能分辨明暗,许多夜行性动物适应性进化出卓越的夜间视觉能力,能够准确辨别颜色、形状、纹理、偏振光和运动状态等特性进行捕食或避敌。Kelber等(2002)首次提出夜行性昆虫具有颜色视觉:象鹰蛾Deilephilaelpenor在夜间微光环境中依然可以准确辨别花朵颜色,这一发现为视觉生态学的研究提供了新的思路。随后,壁虎(Roth and Kelber, 2004)、指狐猴(Perryetal., 2007)、印度木蜂Xylocopatranquebarica(Somanathanetal., 2008)、鹦鹉(Lind and Kelber, 2009)、蝙蝠(Zhaoetal., 2009; Shenetal., 2010)和青蛙(Yovanovichetal., 2020)等多种夜行性动物也被证明具有颜色视觉。这些研究结果推翻了夜间无颜色视觉的传统观点,对暗视觉的基本概念提出了质疑,在视觉生态学领域引起了激烈争论。因此,为了区别于暗视觉,本文进一步明确完善了微光视觉的概念,并界定了其在视觉生态学的作用范围。基于上述夜间微光环境的光照强度范围,将微光视觉定义为在光照强度低于0.3 lx的微光条件下能够保持良好或较好视觉的能力,可以如同在明亮的白天一样清晰、准确地感知目标物体特定的视觉特性,包括明暗(亮度)、颜色、形状、大小、纹理、对比度、偏振光、空间位置和运动状态等。
以周围环境线索在夜间微光条件下作用于视觉器官引起的视觉反应为研究内容,定量评价夜行性昆虫的微光视觉能力,为丰富视觉生态学理论,阐明视觉在昆虫与植物的关系中作用提供了有效途径。目前,有关昆虫微光视觉的研究较少,仅对象鹰蛾D.elpenor(Kelberetal., 2002)、非洲蜣螂Scarabaeuszambesianus(Dackeetal., 2003)、中美洲汗蜂Megaloptagenalis(Greineretal., 2007)、印度木蜂X.tranquebarica(Somanathanetal., 2008)、烟草天蛾Manducasexta(Sponbergetal., 2015)等少数夜行性昆虫的行为反应进行了测定。这些研究结果丰富了视觉生态学理论,打破了以往夜行性昆虫的视觉不起作用的认知误区,展示了微光视觉调控昆虫行为活动的巨大潜力。
2 夜行性昆虫的微光视觉行为反应
2.1 颜色视觉
昆虫一般具有紫外光感受器、蓝光感受器和绿光感受器等,相当于人眼中的视杆细胞和视锥细胞,可以感知周围环境中特定光波的光谱范围(240-700 nm),从而启动昆虫的颜色视觉(color vision)(Briscoe and Chittka, 2001; Schnaitmannetal., 2020)。自1914年von Frisch首次证实西方蜜蜂Apismellifera具有颜色视觉以来,大量的研究证明大部分昆虫尤其是日行性昆虫具有良好的颜色视觉能力,可以准确分辨光源或植物反射的光谱信息,从而在取食、产卵、交配等日常行为活动中快速、准确定位(Reeves, 2011)。例如,蜜蜂、蝴蝶等访花昆虫能够辨别花朵的颜色,以便在觅食过程中采集成熟的花粉或花蜜(Kinoshitaetal., 1999; Blackistonetal., 2011)。果蝇对不同颜色的视觉反应不同,对绿光和紫外光表现出明显的趋向行为,对蓝光则躲避(Lazopuloetal., 2019)。基于昆虫的颜色视觉研发灯诱、色诱等诱捕技术,已成为农林害虫监测和防控中的重要技术,具有良好的应用前景。
由于夜间微光环境的限制,一般认为,大部分夜行性昆虫和人类一样只能看到目标物的灰色影子,无法辨别颜色。事实上,夜间自然环境中的颜色信息和白天一样丰富多彩,差异明显,可以强烈刺激某些夜行性昆虫的颜色视觉。早在21世纪之前,玉米夜蛾Heliothiszea、棉铃虫Helicoverpaarmigera和玉米螟Ostriniafurnacalis等夜行性昆虫已经被证明其复眼可以感知特定光谱范围的光刺激[如紫外线(UV) ]而产生趋光行为(Agee, 1971; 杨智勇等, 1998),但颜色视觉的概念并未明确阐述。Kelber等(2002)首次提出夜行性昆虫和日行性昆虫一样具有颜色视觉,在其取食、产卵等特定行为活动中起着至关重要的作用。通过比较象鹰蛾D.elpenor在不同光照强度下对颜色的辨别能力,发现象鹰蛾能够在昏暗的夜晚准确分辨花朵的颜色,经过训练、学习,即使在星光条件下(0.0001 cd/m2)象鹰蛾仍然可以在一系列的灰色中辨别蓝色和黄色进行觅食,选择率高达80%以上,而人类则失去辨别能力,无法区分。同时,象鹰蛾的颜色视觉具有颜色恒常性(color constancy),即使外界光照环境发生改变时,其对特定颜色的选择行为依旧保持不变(Balkenius and Kelber, 2004)。这一视觉特征对象鹰蛾能够在夜晚正确稳定地感知花朵颜色起着关键作用。此外,具有并列像眼的夜行性印度木蜂X.tranquebarica也被证明具有灵敏的微光视觉能力,在夜间微光条件下同样可以分辨颜色(Somanathanetal., 2008)。
2.2 亮度视觉
亮度视觉(brightness vision)是昆虫视觉行为中另一重要特征,是指辨别颜色明暗、深浅的能力。在自然环境中,植物因种类、生长发育阶段、营养条件等不同可导致植物叶片、茎、花和果实等的亮度特征具有明显差异,如植物叶片的亮度随叶龄的增大而逐渐降低(Farnieretal., 2014)。当植物的反射光谱曲线非常相似时,如植物叶片的光谱范围主要位于500-580 nm,昆虫仅利用颜色视觉并不能有效地辨别寄主植物,亮度视觉可能将在寄主选择过程中发挥关键作用。为了检测昆虫的亮度视觉,一般利用从黑到白的梯度变化或某种颜色的深浅表示不同亮度。已有研究表明,蝴蝶、蜜蜂、天蛾等日行性昆虫可以辨别植物颜色亮度,从而对特定的植物或部位进行准确定位(Hempel de Ibarraetal., 2001; Kinoshitaetal., 2012; Britoetal., 2015)。夜行性昆虫同样具有亮度视觉。Yang等(2020)利用行为学试验测定了夜行性昆虫梨小食心虫Grapholitamolesta在4种不同光照强度(1 000, 100, 1和0.01 mW/m2)下对颜色和亮度的产卵选择行为,结果发现,梨小食心虫具有良好的微光视觉能力,雌成虫在微光条件下依然能够较准确分辨不同亮度的灰色和绿色,而对颜色的正确选择率低于25%,并由此提出了亮度依赖型产卵选择机制(Yangetal., 2020)。
2.3 空间视觉
研究表明,果蝇、蜻蜓、蜜蜂、蝗虫等日行性昆虫具有感知运动状态等空间视觉的能力,能够在复杂背景中探测和追踪运动目标并迅速做出避让、捕食等响应(O′Carrolletal., 1996; Yamaguchietal., 2008)。在探测物体运动时,昆虫能够根据目标和背景的相对运动或者目标物发出的光照强度变化进行识别。不同于日行性昆虫,夜晚微弱的光照强度对夜行性昆虫准确探测目标运动造成了巨大的挑战。尽管如此,一些夜出性昆虫具有非常灵敏的视觉系统,仍然能够在昏暗的夜晚准确探测到微小运动以对运动目标进行有效地视觉追踪。Sponberg等(2015)利用自动移动的假花模拟了黄昏或黎明时烟草天蛾M.sexta盘旋在空中取食花蜜的过程,发现在微光条件下烟草天蛾仍然能够准确探测到微风吹拂中的花朵。在取食过程中,烟草天蛾只能准确追踪到移动频率低于1.7 Hz的假花,当高于1.7 Hz时,其追踪精度显著降低。幸运的是,自然环境中94%花朵在风中的移动频率低于1.7 Hz。此外,通过比较不同光照强度下烟草天蛾对微小运动的追踪反应速度,发现烟草天蛾可以通过减缓其视觉感受速率来补偿性地适应昏暗的环境,在月光下(0.3 lx)对假花的追踪反应速度比在暮光下(300 lx)减慢17%,但其视觉追踪能力并未降低。
形状和大小是昆虫空间视觉反应的重要影响因素(Srinivasan, 2006)。Kaczorowsk等(2012)利用几何形态学定量分析了8种不同烟草属Nicotiana植物花冠形状的基本特征,并在微光条件下测定了形状对烟草天蛾M.sexta觅食行为的影响。结果发现,烟草天蛾对不同形状花冠的选择行为有明显差异,但当花冠大小相同时,则无法准确辨别形状,推测花冠的大小可能是烟草天蛾选择花朵进行觅食的关键因素。随后,Campos等(2015)研究发现,花冠曲率可以影响烟草天蛾对花朵的觅食行为,对喇叭形花朵的偏好性显著高于圆盘形,即使圆盘形花朵向上偏离一较小曲率依然会提高花朵的访问频次。这些研究结果表明,夜行性昆虫同日行性昆虫一样具有空间视觉,能够准确形状、大小和运动状态等视觉线索。
2.4 视觉导航
2.4.1利用天体导航:昆虫的复眼可以在复杂环境中精准检测到太阳、月亮和星星等有效的天体线索(celestial cues),通过接收到的图像帮助其完成觅食、回巢及迁徙等导航行为。其中,月亮是夜间天空的主要天体,其发出的光线是甲虫、蚂蚁和蛾类等夜行性昆虫视觉导航(visual navigation)的重要的定位线索(Sotthibandhu and Baker, 1979; Dackeetal., 2004)。在导航过程中,昆虫头部总是与近似平行的月光保持恒定的夹角直线飞行。由于月亮的方位随时间推移发生明显的变化,对此昆虫必须做出相应的补偿,以平均每小时14.5°的角度调整自己的飞行路线,从而达到准确定位的目的。夜行性昆虫的趋光行为正是由于其天体定向造成的。当月亮被灯光或火光等人造光源取代时,其放射状的光线将导致夜行性昆虫无法正确导航,使得其飞行轨迹发生偏移,螺旋状飞向光源(Kimetal., 2019)。
偏振光模式是昆虫利用天体导航的重要策略。偏振光是太阳光和月光等自然光向地球传播时,部分光波因受到大气层中微粒的散射而产生(蒋月丽等, 2012)。借助对偏振光角度的测算,可以判断相关物体的位置。尽管月光的强度仅有太阳光的百万分之一,一些夜行性昆虫依然可以利用月光的偏振模式进行导航定位。Dacke等(2003)研究发现,夜晚活动的非洲蜣螂S.zambesianus对月光的偏振极为敏感,能够依靠月光偏振模式辨别方向,从而在外出寻找新鲜的粪便时能够保持直线前行并迅速准确返回。在有月光的夜晚,将线性滤光片放置在蜣螂直线滚动粪球路径的正上方,当偏振传输方向与月光偏转90°角时,蜣螂滚动粪球的方向向左或向右偏离90°角;当偏振传输方向与月光平行时,蜣螂继续在原来的方向滚动。随后,中美洲汗蜂M.genalis等其他夜行性昆虫也相继被发现可以利用月光的偏振光作为方向罗盘(Greineretal., 2007)。虽然月光偏振模式的强度随着月亏而下降,并不影响夜行性昆虫视觉导航的精度(Dackeetal., 2011)。
然而,在无月的晴朗夜晚,许多夜行性蜣螂仍可以沿着直线路径移动,推测认为,这些夜行性蜣螂可能和海豹等一样可以利用星星进行导航(Maucketal., 2008)。为了验证这一推测,Dacke等(2013)在天文馆进行了一系列试验,首次发现夜行性萨提洛斯蜣螂Scarabaeussatyrus主要利用像发光带的明亮银河系来定位方向而非单个星体。萨提洛斯蜣螂S.satyrus在银河的指引下,会沿着直线滚动粪球,阴天时则无法直线前进,只能转圈圈。蜣螂夜晚利用银河导航规划路线获得了2013年搞笑诺贝尔生物/天文学奖(Dackeetal., 2013)。这一结果表明了蜣螂能够利用双重天体线索来导航定位,即银河定位导航系统和月光导航系统,但当月亮和银河同时出现时,蜣螂只会利用其中一种。
2.4.2利用地面标志物(landmarks)导航:地面标志物是夜行性昆虫视觉导航另一重要的定向线索。一般而言,昆虫在夜晚利用标志物导航的能力主要依赖于标志物与其背景是否有足够的对比度而不是标志物自身亮度。例如,生活在郁闭森林中的夜行性昆虫日本朱土蝽Parastrachiajaponensis在觅食过程中可以利用树冠及其间隙形成的强烈对比度作为标志物来确定位置和分辨方向(Hironakaetal., 2008)。相比于林冠与层间隙的对比度,巢穴附近具有较低对比度的灌丛和树木等同样可以作为夜晚视觉导航的地面标志物。研究发现,夜行性的中美洲汗蜂M.genalis和印度木蜂X.tranquebarica在黑暗的森林中主要依靠低对比度的地面标志物辨别方向,从而能够在结束漫长的觅食旅行后准确返回巢穴(Warrantetal., 2004; Bairdetal., 2011)。利用地面标志物定位回巢路线是蜜蜂学习记忆的结果。通常,中美洲汗蜂M.genalis的巢穴是一空心棍,当一根蜂巢棍放置在4个空巢棍的中间时,汗蜂能够以此作为视觉标志物,并通过辨认和学习这些空心棍的空间排列来定位蜂巢的位置和方向。如果在汗蜂离开时,将蜂巢棍和空巢棍的位置调换,汗蜂仍然会直接飞进空间位置正确的空巢棍(Somanathanetal., 2008)。大型公牛蚁Myrmeciapyriformis同样具有卓越的夜间徒步回巢能力,也可以在往返巢穴和食物之间利用巢穴附近的标志物导航定位,如果将其移动到正常觅食路径的左边或右边,公牛蚁仍然能够回到正确的路径返回巢穴(Reidetal., 2011)。
2.4.3利用光流导航:当昆虫在自然环境中飞行时,外界物体呈现在其视网膜上的图像也在运动,这种图像的运动模式称之为光流(optical flow)。光流可以基于昆虫移动的速度,感知运动的快慢,为昆虫提供丰富的空间特征信息,使其能够在障碍物之间保持等距飞行。研究表明,蜜蜂、果蝇等日行性昆虫能够利用视网膜上感受到的光流信息来执行速度测量、着陆、姿态控制、躲避障碍物等各种导航任务(Fryetal., 2009; Dyhr and Higgins, 2010)。光流的视觉感知对于控制夜行性昆虫的飞行活动同样具有重要作用。Baird等(2011)研究发现,当水平光流减少时,夜行性中美洲汗蜂M.genalis的对地速度能够显著提高,且不影响其等距飞行的行为,表明中美洲汗蜂能够在微光条件下准确感知光流信息并控制其飞行。随后,通过比较中美洲汗蜂在不同光照强度下的飞行轨迹,发现光照强度并不会影响其飞行和着陆的精确度(Bairdetal., 2015)。这一研究结果进一步证明了夜行性昆虫在微光条件下同样可以利用光流进行导航。
3 夜行性昆虫应对微光环境的视觉适应机制
昆虫具有发达和敏感的视觉器官,可以感受和识别周围环境中特定的光强或光谱刺激,并将这些光信号转变成生物电信号传输到中枢神经,是视觉信息传递的第一站。为了适应微光环境,夜行性昆虫的视觉器官进化出了一些相应的形态生理特征,以提高光子的捕获效率和吸收效率,增强视觉灵敏度,形成有效的视觉信息,使其能够在夜间微光环境中利用颜色、偏振光、标志物和运动状态等视觉线索识别配偶、食物、产卵场所和捕食者等(Honkanenetal., 2014)。揭示昆虫视觉器官应对微光环境的适应性特征不仅为全面阐明夜行性昆虫的微光视觉行为奠定必要的组织学和生理学基础,丰富和完善视觉生态学理论,而且对于发掘昆虫视觉行为调控新策略与新技术等具有重要意义。
复眼(compound eye)是昆虫最重要的视觉器官,由许多独立的小眼(ommatidium)组成镶嵌式结构,可以分为重叠像眼和并列像眼两种类型(Warrant and Dacke, 2011)。大部分夜行性昆虫为重叠像眼,其小眼极度延长,视杆与晶锥分离,呈现一段较宽的透明区,此时视小杆不仅能感受通过该小眼角膜和晶锥的光线,还能感受到邻近若干小眼折射的光线。这一独特的光学结构使得重叠像眼具有强大的折射率径向梯度,可以让通过数百个小眼的光线聚焦到视网膜上的单个光感受器上(Yacketal., 2007)。相比与并列像眼,重叠像眼的视杆较大,同时具有相对较大的瞳孔,可以提高光子的捕获效率,使得复眼在微光环境下清晰成像。研究发现,夜行性双凹蜣螂Onitisaygulus重叠像眼的瞳孔为845 μm,视杆粗大,日行性双凹蜣螂O.belial重叠像眼的瞳孔仅为309 μm且视杆短小,这些形态结构的不同导致了视觉灵敏度的巨大差异(McIntyre and Caveney, 1998)。同时,瞳孔反应是一种更快速的适应策略。复眼可以通过视网膜细胞、初级色素细胞或次级色素细胞内的色素颗粒的纵向移动来调节瞳孔大小,以增加或减少进光量(Yacketal., 2007)。棉铃虫、粘虫Mythimnaseparata、玉米螟等夜行性鳞翅目昆虫,光适应条件下,色素颗粒在视杆周围聚集;暗适应条件下,色素颗粒移动至视网膜远 端靠近晶锥(高慰曾, 1986; Lau and Meyer-Rochow, 2007; Fischeretal., 2014; Chenetal., 2019)。
复眼的光学灵敏度是评估昆虫对光子捕获效率的重要手段,光学灵敏度越高,复眼在特定光环境中吸收的光子数量越多。计算公式为S=(π/4)2A2(d/f)2[kl/(2.3+kl)],其中A为小眼面直径,f为小眼焦距,d和l分别为视杆直径和长度,k为吸收系数。因此,瞳孔直径越大、焦距越短和视杆越宽越长在一定程度上均可以提高昆虫复眼的光学灵敏度,使夜行性昆虫能够在微光环境中清晰准确分辨目标物。一般而言,重叠像眼的光学灵敏度是相同大小并列像眼的100~1 000倍(Warrant and Dacke, 2011)。尽管如此,仍有一部分夜行性昆虫如汗蜂M.genalis、纸黄蜂Apoicapallens等具有并列像眼(Greineretal., 2004; Greiner, 2006)。但理论上,并列像眼并不能在微光条件下清晰成像。为了解释这一矛盾现象,国外研究学者利用组织解剖学和电子显微镜技术等探究了一些夜行性昆虫并列像眼的组织结构,揭示了并列像眼应对微光环境的适应特性。这些夜行性昆虫的并列复眼与日行性昆虫相比在小眼面、小眼焦距、视杆和色素颗粒移动等结构特性方面发生了优化,提高光学灵敏度,使其更适应夜间微光环境。例如,与亲缘关系相近的日行性西方蜜蜂A.mellifera相比,夜行性的中美洲汗蜂M.genalis具有较大的小眼面直径(即瞳孔直径)(A=36 μmvs20 μm),视杆直径是意大利蜜蜂的5倍(d=8 μmvs1.6 μm),导致汗蜂的光学灵敏度提高了27倍,使其表现出良好的微光视觉能力(Greineretal., 2004)。不同于中美洲汗蜂M.genalis,夜行性纸黄蜂A.pallens复眼较高的光学灵敏度则是通过视杆直径和小眼数量来实现的,其视杆直径是日行性长足胡蜂Polistesoccidentalis的4倍(d=8 μmvs2 μm),且小眼数量较多(9 176个vs7 112个)(Greiner, 2006)(表1)。色素颗粒移动同样是并列像眼适应环境的重要调节机制,主要通过视网膜细胞或初级色素细胞内色素颗粒的横向移动来调节视觉灵敏度。光适应条件下,色素颗粒紧紧包围视杆或晶锥;暗适应条件下,色素颗粒则远离视杆或晶锥以提高光吸收效率(Somanathanetal., 2009; Jia and Liang, 2017)。此外,为克服夜间微光环境,夜行性昆虫的并列像眼还特化出了较大角膜内曲率和晶锥等适应结构,以便增加复眼在较低光照强度的灵敏度(Narendraetal., 2013)。
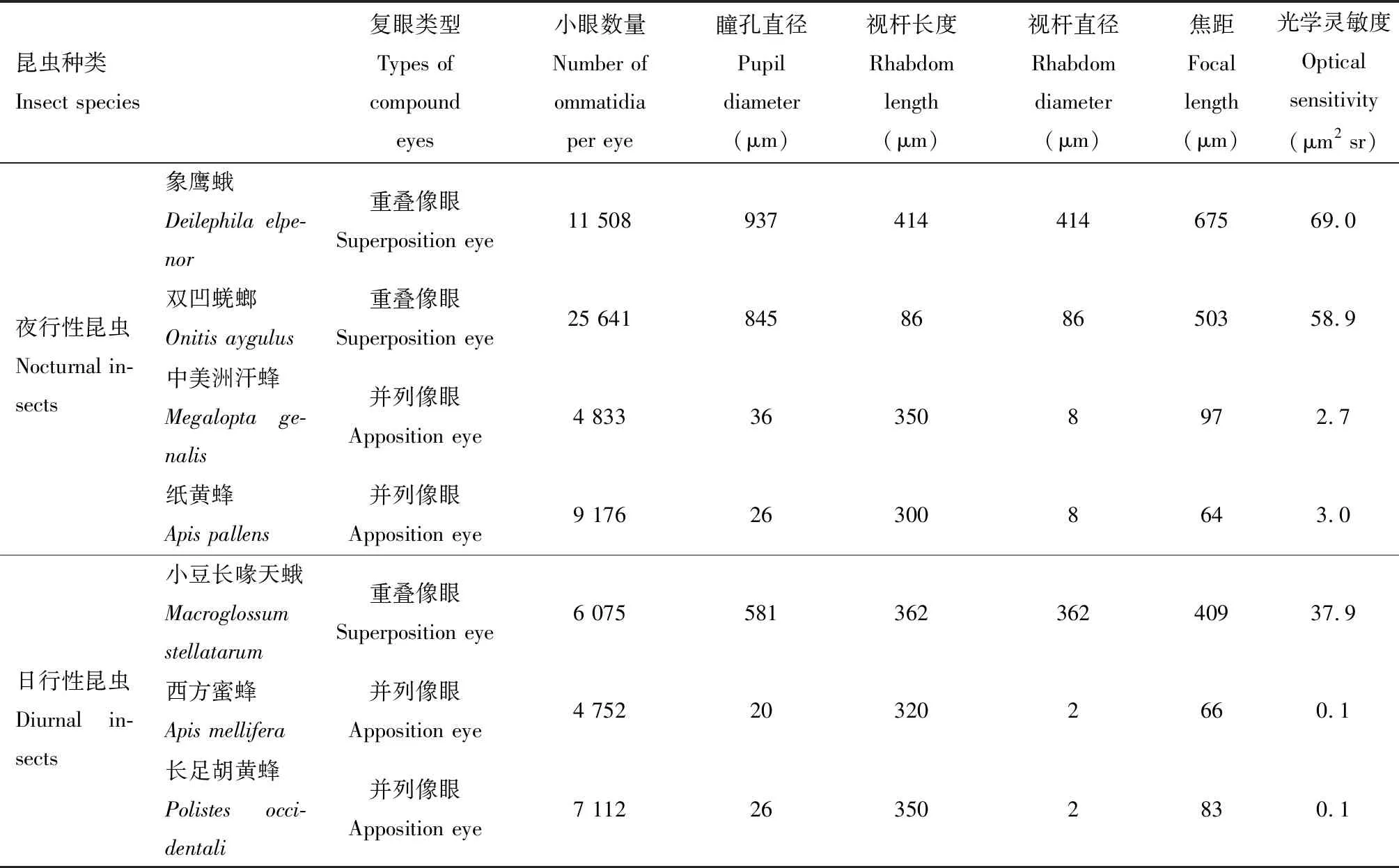
表1 夜行性和日行性昆虫复眼形态结构特征的差异比较Table 1 Comparison of morphological and structural characteristics of compound eyes of nocturnal and diurnal insects
4 小结与展望
近十几年来,昆虫视觉生态学的研究取得了重大进展,随着对视觉行为、生理、神经和基因等特性的深入了解,特别是夜行性昆虫微光视觉的正确解析,加深了视觉调控昆虫行为活动的认识,为昆虫视觉的应用奠定了坚实的基础。但不可否认的是,在昆虫视觉功能和应用方面仍存在许多问题亟待解决,今后应在以下几个方面加强研究:
(1)目前,关于夜行性昆虫微光视觉行为的研究尚处于起步阶段,仅见于少数访花昆虫或粪食性昆虫,而很多农业上的重大害虫具有夜行性。虽然,已有研究学者利用行为学、电生理等测定了一些夜行性农业昆虫如油松球果小卷蛾Gravitarmatamargarotana、小菜蛾Plutellaxylostella、铜绿丽金龟Anomalacorpulenta等的视觉行为反应(李新岗等, 2006; Colaresetal., 2013; Jiangetal., 2015),但多在强光照强度下进行测定,缺乏夜间微光环境下的视觉行为表现。加之,棉铃虫H.armigera、甜菜夜蛾Spodopteraexigua、粘虫M.separata、草地贪夜蛾Spodopterafrugiperda和松毛虫Dendrolimuspunctatus等夜行性农业害虫已初步证明具有UV、蓝光和长波敏感视蛋白基因(Xuetal., 2013; Yanetal., 2014; Liuetal., 2018, 2021; Zhangetal., 2020)。因此,重大农业害虫微光视觉的研究将会是未来视觉生态学重要的发展方向之一。
(2)针对夜行性昆虫复眼对微光环境的视觉适应机制的研究主要集中在模式昆虫和体型较大昆虫的并列复眼或重叠复眼。理论预测认为,由于光学结构的限制,昆虫重叠复眼的半径不能小于250 μm(Meyer-Rochow and Gál, 2004)。最近已有研究证实,潜叶蛾Camerariaohridella、卷叶蛾Adoxophyesorana和微蛾Ectoedemiaargyropeza等微小型蛾类的复眼为非典型重叠复眼,兼具并列复眼和重叠复眼的一些特征,其中小眼中的晶锥和视杆直接相连,透明区消失(Fischeretal., 2012, 2014; Satohetal., 2017)。然而,这些研究仅仅分析了非典型重叠复眼的光学结构特征,关于非典型重叠复眼应对微光或变化光环境的适应机制尚不清楚。此外,非典型重叠复眼与重叠复眼和并列复眼的光环境适应机制有何不同?这些关键问题仍未得到答案。这对于全面深入解析夜行性昆虫的微光视觉行为至关重要。
(3)视蛋白是昆虫视觉信息传递过程中的一类重要分子。日行性昆虫和夜行性昆虫在长期进化过程中受不同光环境的影响,其视蛋白数量、种类、序列结构和生物学功能等方面可能存在差异。利用基因组学、转录组学和蛋白组学等分子生物学相关方法和技术,从遗传进化角度探究日行性昆虫和夜行性昆虫视蛋白基因的差异变化,对进一步系统揭示昆虫对微光环境的视觉适应机制具有重要意义。
(4)基于夜行性昆虫视觉行为研发高效、绿色的害虫监测和防控技术已成为我国农药减量和替代的战略需求,其中,害虫灯光诱控技术已在全国多个省市的农林生产中推广应用,但主要局限于趋光反应(桑文等, 2019)。因此,在深入解析夜行性昆虫的颜色视觉、空间视觉、运动视觉、视觉导航等微光视觉行为及功能的同时,应充分考虑各种视觉线索的协调调控作用,筛选新型的行为调控技术,为害虫的可持续防控提供支撑。
