投喂盐富饶菌Haloferax YC-6强化卤虫对凡纳滨对虾 生长和抗逆能力的影响
2021-12-22郭子仙任北妮高美荣隋丽英
郭子仙,解 伟,任北妮,段 虎,高美荣,隋丽英
(天津科技大学海洋与环境学院,天津 300457)
凡纳滨对虾(Litopenaeus vannamei)是世界养殖产量最高的优良对虾品种,2019年养殖产量达到496.72万吨,我国凡纳滨对虾产量达到181.55万 吨[1–2].近年来,水产养殖环境恶化和种质退化等问题导致疾病频繁发生,使凡纳滨对虾养殖产业面临严峻挑战[3].对虾拥有开放循环系统,极易受到养殖水体环境的影响,而环境胁迫造成的应激对对虾机体抗氧化和免疫系统造成损伤[4].将功能性添加剂如益生菌、抗菌肽等应用于凡纳滨对虾养殖是提高对虾免疫力、促进对虾生长的重要途径[5-7].
古菌几乎能适应地球上所有极端环境,古菌细胞可积累相容性溶质、表层蛋白、类胡萝卜素等多种生物活性物质[8–9].盐富饶菌(Haloferax)是在高盐环境下生长的古菌属,具有生长快、耐盐范围广和代谢功能强等特点,因富含菌红素、单脱水菌红素和双脱水菌红素等类胡萝卜素而呈红色[10].菌红素是一种长链C50类胡萝卜素,包含13对共轭双键,且末端含有羟基,是一种有效的自由基清除剂,具有较高的DPPH自由基清除能力,其抗氧化能力高于β–胡萝卜素[11–13]. 研究表明,类胡萝卜素能提高水产动物的经济性状和品质.富含类胡萝卜素的饲料能够促进虹鳟鱼(On-corhynchus mykiss)生长,提高色素含量[14]. 向黄颡鱼(Pelteobagrus fulvidraco)投喂含β–胡萝卜素的饲料可提高其肝脏中抗氧化酶活性和热休克蛋白HSP70基因的表达,进而降低由高温胁迫和病原菌感染造成的死亡率[15].但古菌及其类胡萝卜素作为饲料添加剂在水产养殖乃至畜禽养殖中的应用未见报道.
卤虫是海水苗种培育重要的生物饵料.作为非选择性滤食生物,卤虫可高效滤食单胞藻、细菌和微小的有机碎屑等.水产育苗中常采用强化方式使卤虫肠道包裹携带营养强化剂(如鱼油)和药物等,通过投喂强化卤虫将其有效送达水产苗种体内.本研究将古菌Haloferax作为添加剂,通过卤虫强化的方式将其投喂给凡纳滨对虾仔虾,从生长性能、肠道菌群组成和抗逆性等方面探讨其对凡纳滨对虾的影响.
1 材料与方法
1.1 发酵培养
盐富饶菌菌株HaloferaxYC-6分离于天津汉沽盐场结晶池卤水,将其接种于培养基(7.5g/L酸水解酪蛋白,10g/L酵母提取物,100g/L稀释卤水),150r/min、37℃和光照(2000lx)条件下摇瓶培养至对数生长期获得种子液.添加5g/L蔗糖于上述培养基中进行发酵培养,发酵体积为3L.发酵条件:pH 5.5~8.5,温度37℃,初始搅拌速度200r/min,每24h升高100r/min直到400r/min.发酵5d后8000r/min离心10min收集菌体,4℃避光保存.
1.2 卤虫强化
将美国大盐湖卤虫卵(比利时INVE公司)在28℃和盐度30g/L稀释卤水中连续充气孵化,24h后收集卤虫无节幼体,转移至盐度50g/L稀释卤水中,以HaloferaxYC-6菌体强化卤虫.卤虫密度为200mL-1,菌体投加量(以干质量计)为0.3g/L.强化12h后收集卤虫,用相同盐度的稀释卤水冲洗,4℃充气保存,以维持卤虫存活和营养质量.
1.3 实验动物
实验所用凡纳滨对虾仔虾购自河北鑫海生物技术公司.将PL5仔虾在28℃、盐度20g/L稀释卤水中暂养3d,暂养过程中投喂虾片饲料.
1.4 实验设计
选出活力好且大小均匀的虾苗(平均体长0.53cm)随机分为2组,分别以饥饿12h卤虫和Haloferax菌体强化12h卤虫作为对照组和实验组,投喂凡纳滨对虾.饥饿卤虫和Haloferax强化卤虫的营养组成(以干质量计)见表1.

表1 饥饿卤虫和Haloferax强化卤虫的营养组成 Tab. 1 Nutrient composition of starved and Haloferaxenriched Artemia
养殖在循环过滤养殖系统中进行,用海绵过滤食物残渣和对虾粪便.养殖箱为长方形聚丙烯箱体(48cm×26cm×39cm),每组设3个平行,每个平行箱投放虾苗1000尾,单位养殖水体为20L.养殖期间每天投喂3次,每尾虾每次的初始投喂量为100个卤虫,投喂量按前一天的10%增加.养殖周期为10d,从第6天开始每天换1/3的水.养殖水体盐度为20g/L,水温(28±1.0)℃,溶氧量(5.5±0.5)mg/L,pH 7.6±0.2,连续充气,光/暗周期为16h/8h.
1.5 样品采集与指标分析
1.5.1 肠道菌群分析
养殖结束后测定对虾存活率,并从每个平行箱中随机取10尾虾测量体长(自对虾眼柄基部至尾节末端的长度).从每个平行箱取5尾饥饿24h的对虾,置于冰上麻醉,无菌条件下解剖取出对虾肠道,立即放入灭菌的EP管中液氮速冻,置于-80℃保存.将样品反复冻融后匀浆,采用细菌DNA提取试剂盒(天根生化科技公司)提取肠道菌群DNA,样品交予诺禾致源公司使用TruSeq®DNA PCR-Free Sample Preparation Kit建库试剂盒进行文库构建,构建好的文库经过Qubit和Q-PCR定量检测文库合格后,使用NovaSeq6000进行上机测序.使用Qiime软件(Version 1.9.1)计算Chao1指数、香农指数Shannon、辛普森多样性指数Simpson、ACE指数和覆盖率Good-coverage指数.
1.5.2 对虾粗蛋白、粗脂肪和脂肪酸测定
将收集的卤虫和对虾进行冷冻干燥(BTP-3ES型冷冻干燥机,美国SP-Scientific公司).称取0.05g冷冻干燥样品,参照GB/T 6432—2018《饲料中粗蛋白的测定·凯氏定氮法》,利用Kjeltec–8400型全自动凯式定氮仪(丹麦FOOS公司)测定对虾粗蛋白质含量.称取0.2g冷冻干燥样品,参照GB 5009.6—2016《食品安全国家标准·食品中脂肪的测定》,用索氏抽提法测定对虾粗脂肪含量.称取0.04g冷冻干燥样品,准确加入1mg内标液(C20∶2n-6,美国NUCHEK PREP公司),向混合物中加入5mL甲醇与甲苯混合物(体积比为3∶2)和5mL氯乙酰与甲醇混合物(体积比为1∶20),充分匀浆(T18型匀浆机,德国IKA公司)后,沸水浴1h.用正己烷萃取脂肪酸甲酯并浓缩[16],利用GC–2014型气相色谱仪(日本岛津公司)进行脂肪酸定性和定量测定.
1.5.3 弧菌攻毒实验
副溶血弧菌(Vibro parahaemolyticus MCCC 1A10122)购于海洋微生物菌种保藏管理中心(https:// mccc.org.cn/).菌种在28℃、150r/min条件下于2216E培养基培养至对数生长期.养殖结束后每缸随机取出20尾虾,根据预实验结果,以弧菌108mL-1剂量浸浴攻毒48h,每隔24h测定对虾存活率.
1.5.4 氨氮胁迫实验
每个平行箱中随机各取出20尾虾,置于1L含30mg/L氯化铵、盐度20g/L的水体中.溶液中非离子氨(NH3-N)质量浓度为0.8mg/L[17].每隔24h测定对虾存活率.
1.5.5 对虾肝胰腺抗氧化酶活性
养殖结束后和氨氮胁迫后,在无菌条件下解剖获得对虾肝胰腺,用试剂盒(南京建成生物工程研究所)测定其过氧化氢酶(CAT)和超氧化物歧化酶(SOD)活性以及丙二醛(MDA)含量.
1.6 统计分析
数据用“平均值±标准差”表示,用SPSS statistics analysis 20.0软件进行独立样本t检验,*表示有显著差异(P<0.05).
2 结果与分析
2.1 凡纳滨对虾仔虾存活和生长
投喂Haloferax强化的卤虫对凡纳滨对虾仔虾存活和生长的影响见表2.投喂Haloferax强化卤虫的实验组对虾的存活率和体长均显著高于对照组(P<0.05).

表2 投喂Haloferax强化的卤虫对凡纳滨对虾仔虾存活和生长的影响 Tab. 2 Effect of Haloferax-enriched Artemiafeeding on the survival and growth of L. vannamei postlarvae
2.2 凡纳滨对虾营养组成
不同组别凡纳滨对虾营养组成见表3.两组对虾粗蛋白质和粗脂肪含量均无显著性差异.实验组的C14∶1n-5、C18∶0、C18∶1n-9+n-7和C18∶4含量显著低于对照组(P<0.05),其他脂肪酸含量均无显著性差异.
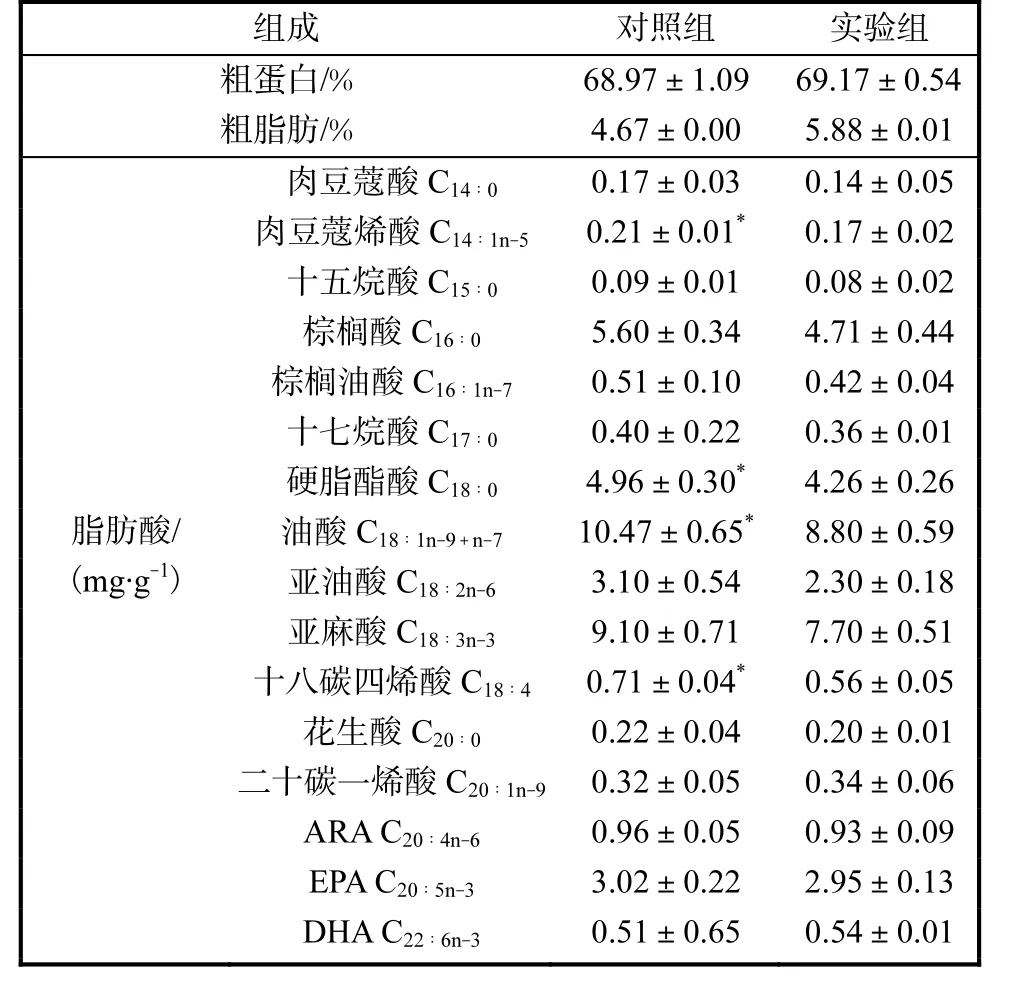
表3 不同组别凡纳滨对虾营养组成 Tab. 3 Nutritional composition of L. vannamei in different groups
2.3 凡纳滨对虾肠道菌群多样性
不同组别凡纳滨对虾肠道微生物α 多样性见表4.对照组和实验组的Good-coverage覆盖率值均为0.99,表明每个文库的16S rRNA基因代表了对虾肠道中的大多数细菌.实验组对虾肠道微生物多样性指数均有所上升,但与对照组没有显著差异(P>0.05).
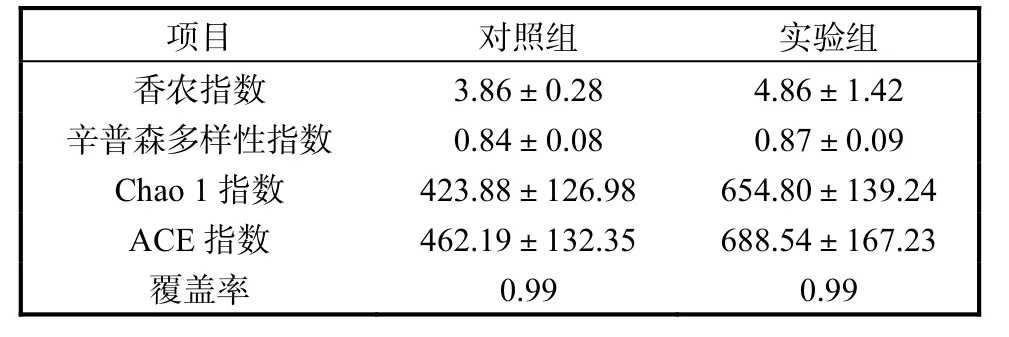
表4 不同组别凡纳滨对虾肠道微生物α 多样性 Tab. 4 Alfa-diversity of intestinal microbiota of L.vannamei postlarvae in different groups
凡纳滨对虾肠道菌群结构门水平变化如图1(a)所示,对虾肠道菌群主要以变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)和放线菌门(Actinobacteria)为主.与对照组相比,实验组变形菌门和放线菌门的相对丰度有所下降,厚壁菌门和拟杆菌门(Bacteroidetes)相对丰度提高.凡纳滨对虾肠道菌群属水平变化如图1(b)所示,实验组弧菌属(Vibro)比例下降,副球菌属(Paracoccus)、拟杆菌属(Bacteroides)、微小杆菌属(Acteroides)和乳杆菌属(Lactobacillus)的相对丰度均有所上升.
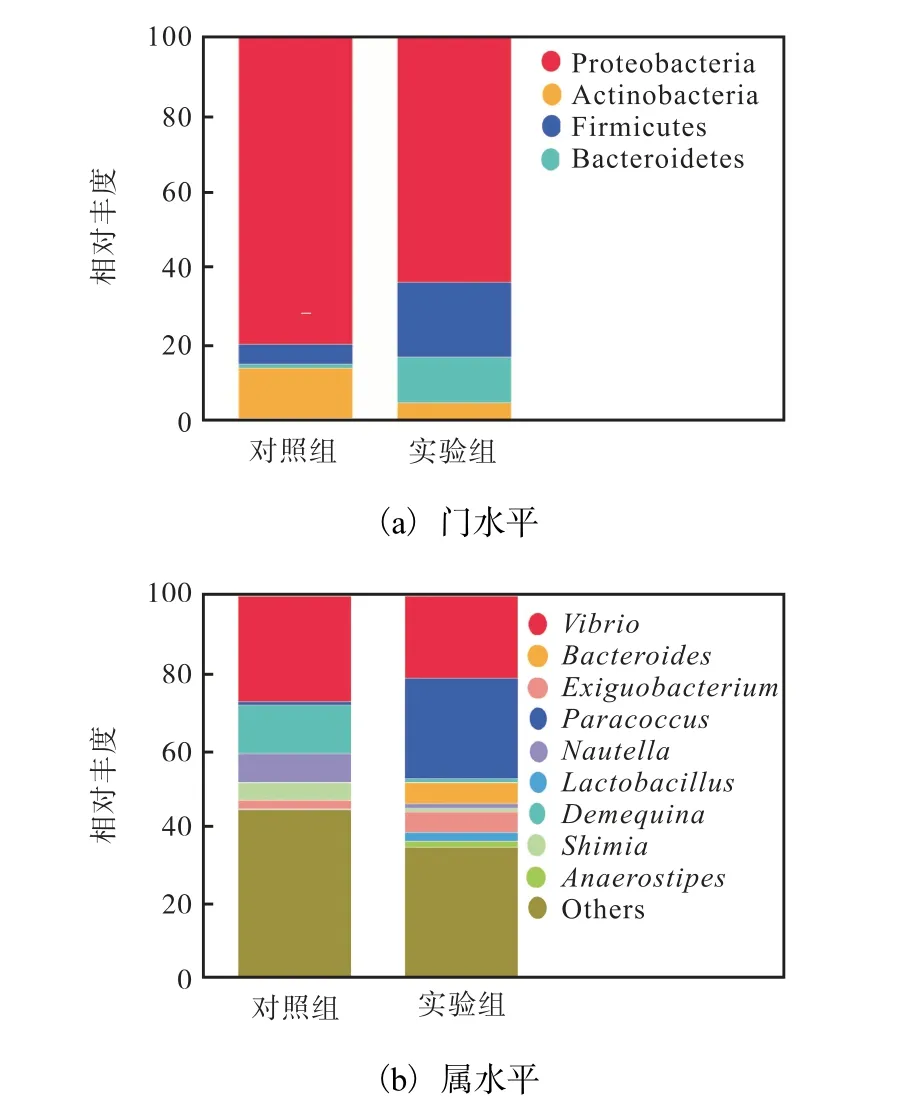
图1 投喂Haloferax强化卤虫对凡纳滨对虾肠道菌群门和属水平的影响 Fig. 1 Effect of Haloferax-enriched Artemiafeeding on the intestinal microbiota of L. vannameipostlarvae in phylum level and in genus level
2.4 凡纳滨对虾肝胰腺抗氧化能力
投喂Haloferax强化卤虫对对虾肝胰腺抗氧化能力的影响结果见表5.与对照组相比,养殖结束后实验组对虾肝胰腺SOD活性有所上升,但没有显著性差异.实验组对虾肝胰腺CAT活性显著高于对照组(P<0.05),MDA含量显著低于对照组(P<0.05).

表5 投喂Haloferax强化卤虫对对虾肝胰腺抗氧化能力的影响 Tab. 5 Effect of Haloferax-enriched Artemiafeeding on antioxidative capacity of L. vannamei postlarvae
2.5 凡纳滨对虾抗胁迫能力
2.5.1 氨氮胁迫
氨氮胁迫对凡纳滨对虾存活率的影响如图2所示.24h 对照组和实验组对虾存活率没有显著差异,但氨氮胁迫48h后,实验组存活率显著高于对照组(P<0.05).

图2 氨氮胁迫对凡纳滨对虾存活率的影响 Fig. 2 Effect of ammonia nitrogen on survival rate of L. vannamei
投喂Haloferax强化的卤虫对氨氮胁迫48h后对虾肝胰腺抗氧化能力的影响结果见表6.氨氮胁迫48h后,实验组对虾肝胰腺SOD活性有所降低,但差异不显著(P>0.05),CAT活性和MDA含量显著降低(P<0.05).

表6 投喂Haloferax强化卤虫对氨氮胁迫48h后对虾肝胰腺抗氧化能力的影响 Tab. 6 Effect of Haloferax-enriched Artemia feeding on antioxidative capacity of L.vannamei postlarvae after being exposed to ammonia nitrogen for 48 h
2.5.2 副溶血弧菌攻毒
副溶血弧菌攻毒对凡纳滨对虾存活率的影响如图3所示.攻毒24h的对照组和实验组对虾存活率没有显著差异,在91.67%~93.33%之间.攻毒48h后,实验组对虾存活率显著高于对照组(P<0.05).
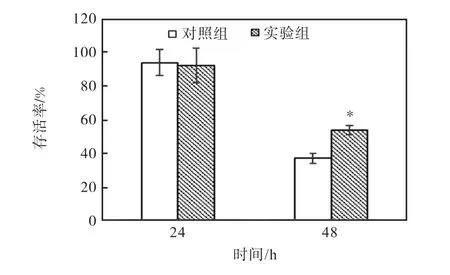
图3 副溶血弧菌攻毒对凡纳滨对虾存活率的影响 Fig. 3 Effect of challenge with V. parahaemolyticus on survival rate of L. vannamei
3 讨 论
3.1 Haloferax对凡纳滨对虾生长和营养组成的影响
本文研究结果表明,投喂Haloferax强化卤虫能显著提高对虾仔虾的存活率,促进对虾生长.有研 究[15,18]表明,饲料中类胡萝卜素的添加起到促进水产动物生长和提高抗逆能力的作用.Haloferax对对虾的促进作用可能与细胞膜上大量积累的类胡萝卜素有关.
本研究中强化卤虫的各种脂肪酸含量较饥饿卤虫高,与卤虫在饥饿状态下代谢脂肪酸等营养物质为其提供生长存活必需的能量有关.一般而言,n-3多不饱和脂肪酸特别是高不饱和脂肪酸(如EPA、ARA和DHA)对海洋水产动物细胞膜的流动性和渗透性有较大影响[16,19].对照组对虾中C14∶1n-5、C18∶0、 C18∶1n-9+n-7和C18∶4含量显著高于实验组,可能与对虾本身的脂肪酸组成和对饵料中脂肪酸的选择性代谢有关.
3.2 Haloferax对凡纳滨对虾肠道菌群的影响
肠道菌群对对虾维持机体健康和免疫稳态非常重要[20].本研究中Haloferax的添加在一定程度上增加了对虾肠道菌群的多样性,同时Haloferax的摄入降低了变形菌门丰度,主要表现在弧菌属的丰度降低. 变形菌门广泛存在于水生无脊椎动物肠道微生物群中,通常是甲壳类动物肠道菌群的主要组成部分[21]. 弧菌是对虾肠道的优势菌属,部分弧菌产生的几丁质酶有助于其在肠道菌群形成优势菌,与宿主共存[22]. 但弧菌属于条件致病菌,其数量和毒力与环境因子有关,环境胁迫造成弧菌的数量和毒力增加,进而致病性增强,导致对虾的死亡率较高[23-24].本研究中Haloferax的添加在一定程度上降低了弧菌属的丰度,提高了乳杆菌属的丰度,对对虾肠道健康起到有益作用.
3.3 Haloferax对凡纳滨对虾抗氨氮胁迫能力的影响
古菌细胞膜中含有大量菌红素等长链C50类胡萝卜素[10].与β–类胡萝卜素相比,Haloferax类胡萝卜素提取物可更有效地清除DPPH自由基,并对H2O2介导的红细胞溶血具有更好的保护作用[25].研究[15,26]表明,类胡萝卜素的添加对水生动物产生积极的效果,改善由养殖密度过高、高温胁迫或病原菌感染造成的不良影响.在脊椎动物中,类胡萝卜素发挥了其抗氧化和免疫作用,可以清除过量自由基从而降低自我损伤[27].类胡萝卜素也被证实能够提高甲壳动物免疫力和抗氧化能力[28-29].
氨氮是影响对虾存活和生长的重要环境因子. 当暴露于高浓度氨氮时,对虾机体发生一系列改变,包括肝胰腺损伤、细胞凋亡以及免疫损伤等[30-31].转录组分析[32]表明,氨氮暴露破坏对虾氧化和抗氧化平衡,造成机体的氧化应激.机体的抗氧化系统包括两类:酶类(如SOD和CAT等)和小分子非酶类(如谷胱甘肽、类胡萝卜素和维生素等).研究[33]表明,对虾CAT、SOD、谷胱甘肽过氧化物酶(GSH-Px)等抗氧化酶在对虾氨氮防御中发挥重要作用.本研究发现,养殖过程中Haloferax的添加使对虾肝胰腺抗氧化酶活力上升,其中CAT活性显著增加.氨氮胁迫48h后,实验组对虾的存活率显著高于对照组,表明古菌提高了对虾抵抗氨氮胁迫的能力,与古菌的摄入提高了对虾抗氧化酶活性进而发挥了氨氮防御功能有关.值得一提的是,氨氮胁迫后实验组对虾肝胰腺SOD活性有所下降,CAT活性和MDA含量显著降低,可能与机体抗氧化系统的反应机理有关.在抗氧化系统中,SOD作为清除活性氧和自由基的第一道防线,能将活性氧转化为过氧化氢和氧气,然后过氧化氢通过CAT作用转化为水和氧气.MDA是脂质过氧化的终产物,其含量可以作为细胞膜氧化损伤的标志[34].本研究中氨氮胁迫后实验组MDA含量显著降低,表明实验组对虾机体氧化损伤程度较轻,而CAT活性降低可能是因为Haloferax提供的类胡萝卜素代替抗氧化酶发挥了清除活性氧和自由基的功能,进而保护对虾免受氧化应激损伤.
3.4 Haloferax对凡纳滨对虾抗弧菌能力的影响
副溶血弧菌是对虾急性肝胰腺坏死综合征(acute hepatopancreatic necrosis syndrome,AHPNS)的主要病原体.除损伤肝胰腺外,弧菌感染还可损伤对虾肠道的屏障功能,造成对虾感染和死亡[35].本研究中Haloferax的添加提升了凡纳滨对虾对弧菌的抵抗能力,显著提高了其存活率.其原因一方面可能是Haloferax的摄入降低了对虾肠道内弧菌属的丰度,缓解了弧菌感染的程度;另一方面可能与Haloferax含有的活性物质有关.由于对高盐环境的适应性,嗜盐古菌产生具有生物活性的初级和次级代谢产物,如酶类、类胡萝卜素、PHB以及嗜盐菌素等[9,36-38],这些物质可能成为对虾抗弧菌感染的贡献者.
4 结 论
将古菌Haloferax作为饲料添加剂通过卤虫强化方式投喂给凡纳滨对虾仔虾,可以促进对虾的存活和生长,改善对虾肠道菌群,增强对虾抵抗弧菌和抗氨氮胁迫能力.Haloferax菌体富含大量高抗氧化活性的C50类胡萝卜素,从而提高对虾机体抗氧化能力,增强对虾对弧菌的抵抗力.本研究结果为进一步研究古菌在水产养殖中的应用提供了依据.
