钙化与非钙化株系颗石藻Emiliania huxleyi胞内 元素组成对氮限制的响应比较
2021-12-22冯媛媛侯丹丹李美琪
郭 佳,冯媛媛,侯丹丹,李美琪
(天津科技大学海洋与环境学院,天津 300457)
海洋颗石藻属于定鞭藻门,是一种可通过生物钙化作用生产颗石粒的单细胞浮游植物[1].颗石藻大概贡献了全球海洋碳酸钙生产总量的50%[2],是海洋碳循环的重要组成部分.一方面,颗石藻可通过光合作用吸收大气中的二氧化碳(CO2)进行有机碳的生产;另一方面,颗石藻通过钙化作用进行无机碳生产,合成其钙质化外壳,该过程释放CO2并改变海水总碱度[3].颗石藻在除极地及热带海域外的全球海洋中广泛分布,通过卫星遥感图像可观测到颗石藻在很多海域中出现大规模季节性水华[2,4-5],其最具广布性的优势物种Emiliania huxleyi是海洋碳循环研究中的模式物种[6].
颗石藻的各种生理过程,尤其是钙化作用对海洋酸化的响应尤为敏感[7–10],但其响应模式存在明显的种间及株间效应[11].除海洋酸化外,其生长、光合作用及钙化作用还会受到温度、光照强度、营养盐浓度变化等其他环境因子的影响[12–16].氮是对海洋浮游植物生长和初级生产起到重要作用的营养元素,是浮游植物细胞内生物大分子的重要组成元素之一,氮限制或缺乏都会影响浮游植物生长与胞内的生理代 谢[17–18].有研究[19]表明,氮的缺乏会改变藻细胞内物质的组成,如碳水化合物、油脂含量升高,蛋白质含量下降.Berges 等[20]研究表明,在氮限制的条件下威氏海链藻(Thalassbsira weissflogii)细胞内由于诱导产生一种蛋白酶会导致大部分蛋白酶的活性也随之增加,从而使胞内蛋白质含量显著下降.Han等[21]通过对蛋白核小球藻(Chlorella pyrenoidosa)进行半连续培养发现细胞的脂质含量在氮缺乏的环境中升高.还有部分研究人员研究多种微藻在氮限制条件下的生理指标的变化,他们的研究[22–24]结果表明,氮限制的程度与时间会对不同种系微藻的生物量、生长速率及光合速率产生一定程度的影响;研究还发现在低氮供应时,小新月菱形藻胞内的叶绿素(Chl-a)含量及光合效率均呈现下降趋势,减缓了藻细胞的生长.氮限制会影响颗石藻光合和钙化作用中重要功能性蛋白质的生产,硝酸盐的浓度对颗石藻的生长、光合固碳与钙化作用起到重要的调节作用[15],并且可以与海洋酸化对颗石藻的生理过程产生交互效 应[12,25–27].也有研究[16]表明E. huxleyi在硝酸盐供应量减少的条件下细胞颗粒无机碳(PIC)含量增加.然而,很少有研究阐明氮限制条件下颗石藻是否存在株间差异,尤其是钙化和非钙化株系之间的差异可为我们进一步认识颗石藻的钙化作用在其生理调节上的意义提供参考信息.在氮限制条件下,我们假设钙化株系会因为其胞外有钙质化外壳的保护相较于非钙化株系的各生理学参数会受到较小的影响.本研究选取颗石藻优势物种E. huxleyi钙化株系和非钙化株系,采用恒化连续培养的方式进行实验室内受控连续培养实验,研究两株系的颗石藻于指数生长期稳态条件下氮限制对其生理指标的影响.该方法与传统一次性培养实验方法不同:一次性培养实验达到的氮限制条件通常是培养基中营养盐消耗殆尽时,藻细胞进入平台生长期,濒临死亡期,其生长状态与藻类爆发大规模水华后处于衰退期的状态相近;而恒化连续培养更好地模拟了自然稳态条件下低硝酸盐海水中颗石藻的生长状态.
1 材料与方法
1.1 实验设置
选取颗石藻E. huxleyi的两种不同株系,钙化株系E. huxleyiNIWA1108于2009年分离自新西兰以东Chatham Rise海域,非钙化株系E. huxleyiPMLB分离自欧洲北海.两株藻种均采用f/20自然海水培养基[28](f/2培养基稀释10倍)并放置于环境温度为15℃、光照强度为100~120μmol/(m2·s)的恒温光照培养箱中进行保种培养,光暗周期为12h∶12h.配制培养基所使用的自然海水采自南黄海寡营养盐海域的表层海水,并使用玻璃纤维滤膜经隔膜真空泵抽滤后用立式自动压力蒸汽灭菌锅高温灭菌(121℃,15min),以达到无菌的效果,冷却后备用.
1.2 连续培养实验
培养实验采用自制的恒化培养器(包括蠕动泵、装有搅拌器的培养瓶、排水管等)进行连续培养.对每个株系设置2个营养盐浓度:(1)氮限制,N/P为1.6(物质的量比);(2)氮充足,N/P为16(物质的量比).培养基采用过滤后高温灭菌的南黄海表层海水,磷酸盐、微量元素及维生素按照f/20配方添加.硝酸盐浓度分别添加至N/P=1.6和N/P=16水平.将预培养的处于指数生长期的藻液接种到3.5L装有培养基的培养瓶(聚碳酸酯材质)中,初始细胞密度设定为10000mL-1.在培养箱内采用恒化培养器进行连续培养,将蠕动泵在藻细胞接种后的第3天开启,开始连续培养实验.使用蠕动泵将培养基连续泵入培养瓶中,藻液流出口与每个培养瓶的瓶颈处相连接,以保证各培养体系中藻细胞的丰度和体积维持恒定.对于氮充足处理组的蠕动泵稀释速率设置为0.5d-1,氮限制处理组设置为0.2d-1.实验过程中为了达到培养瓶中藻液分布均匀的状态,采用搅拌器(具有特氟龙涂层)在培养瓶中连续低速搅拌.此连续培养实验在恒温光照培养箱中进行,光照条件和温度与保种条件保持一致.每个处理组下均设置3个平行样.
连续培养开始后,每隔24h取3mL藻液,分成2份:一份用Turner荧光仪测定其活体荧光值;一份用碱性鲁格氏试剂(Lugol’s)固定后,放置到XS–213显微镜下进行观察和细胞计数.培养至其指数生长阶段并进入稳态超过5d(每个处理组中藻细胞丰度保持相对稳定,变化小于10%)后,进行最终采样,采样参数为活体荧光、细胞计数、叶绿素a(Chl-a)含量、沉降速率以及细胞颗粒有机磷(POP)、颗粒有机碳(POC)、颗粒有机氮(PON)含量.本实验最终培养时间为21d.
1.3 样品分析
1.3.1 藻细胞计数及Chl-a质量浓度的测定
取1mL藻液于1.5mL的离心管中,并加入6μL碱性鲁格试剂(Lugol’s)[29],放置于黑暗处4℃保存,最后在光学生物显微镜下用0.070mL的微藻计数框观测计数,藻细胞计数所用样品的保存时间不可以超过1周.
量取50mL藻液经六联过滤器(隔膜真空泵抽滤压力小于0.2MPa)过滤到GF/F玻璃纤维膜上.为避免Chl-a见光分解,将滤膜对折后放置于铝箔纸材质的小袋中,保存于-20℃的冰柜.对样品进行分析时需在暗处进行,将滤膜置于装有5mL体积分数90%的丙酮的棕色试剂瓶中,在-20℃环境下暗处理24h后,使用Turner荧光仪的非酸化模式测定其荧光值,根据式(1)[30]计算Chl-a质量浓度(μg/L).

1.3.2 细胞元素组成的分析
POP含量的测定采用分光光度法[31].准确量取50mL藻液,经六联过滤器过滤到马弗炉灼烧(450℃,4h)过的GF/F玻璃纤维膜上,另取3份等体积的相应背景海水培养基,与上述操作相同,作为空白样.所取样品需用2mL 0.17mol/L Na2SO4溶液润洗经六联过滤器真空泵抽滤.再将抽滤后的滤膜全部浸入到盛有2mL 0.017mol/L的MgSO4溶液且经马弗炉灼烧(450℃,4h)过的样品瓶中,用灼烧过的铝箔纸轻轻盖住瓶口,最后将其置于60℃烘箱中,直至烘干.分析样品前,将装有滤膜的玻璃瓶置于马弗炉中,450℃灼烧2h,冷却到室温后将5mL 0.2mol/L的盐酸加入到样品瓶内,用灼烧过的铝箔纸封住瓶口置于90℃烘箱中烘烤30min后取出.冷却到室温后,将0.5mL显色剂加入到样品瓶中,摇匀,显色10~20min,采用紫外可见分光光度计测定其吸光度,最后根据标准曲线以及过滤体积计算POP含量.
采用CHN元素分析仪对POC和PON含量进行测定[32].从每个培养瓶中量取2份100mL藻液过滤至预先经马弗炉灼烧(450℃,4h)过的GF/F膜上,置于60℃的烘箱中烘干.其中一份直接用以测定总颗粒有机碳(TPC)以及PON的含量,另一份采用浓盐酸熏蒸3h后再次烘干,用于测定颗粒有机碳(POC)的含量.
1.4 沉降速率的计算
把沉降柱垂直固定于支架上,堵住3个出水口,将藻液混匀,倒入沉降柱中,让藻液充满整个沉降柱,然后轻轻盖上盖子,使沉降柱保持密封(尽量避免产生空隙),置于同一温度下避光静置2~4h.收样时,从上到下分层依次取样,记录所取各层藻液体积,并分别过滤到GF/F玻璃纤维膜上.将滤膜对折,迅速放入叠好的铝箔小袋内,于-20℃冰箱内保存,用于最后测定其叶绿素含量.浮游植物的沉降速率根据最后测定的叶绿素含量并采用Bienfang的沉降速率公式[33]进行计算.
1.5 统计分析
组间差异检验采用t检验(SPSS9.5软件),P<0.05时为显著性差异.文中误差棒均为标准偏差,n=3.
2 结果
2.1 胞内Chl-a含量
颗石藻钙化株系和非钙化株系的胞内Chl-a含量如图1所示.

图1 钙化株系和非钙化株系的胞内Chl-a含量 Fig. 1 Cellular Chl-a contents of calcified strain and noncalcified strain
氮限制(稀释速率0.2d-1)条件下,两株系的胞内Chl-a含量均较低,非钙化株系胞内Chl-a含量与钙化株系胞内Chl-a含量相比降低44.6%.钙化株系在氮限制的条件下相较于氮充足条件的胞内Chl-a含量下降85.2%;非钙化株系的胞内Chl-a含量在氮限制的条件下相较于氮充足条件降低89.4%.
2.2 细胞元素组成及比值
颗石藻钙化株系和非钙化株系的胞内POC含量如图2所示.两个株系的单位细胞内POC含量均受到氮限制的影响.钙化株系的胞内POC含量在氮限制的条件下相较于氮充足的条件降低21.9%;非钙化株系的胞内POC含量在氮限制的条件下也相较于氮充足的条件降低33.6%.在氮限制条件下,非钙化株系的胞内POC含量相比于钙化株系低37.2%.
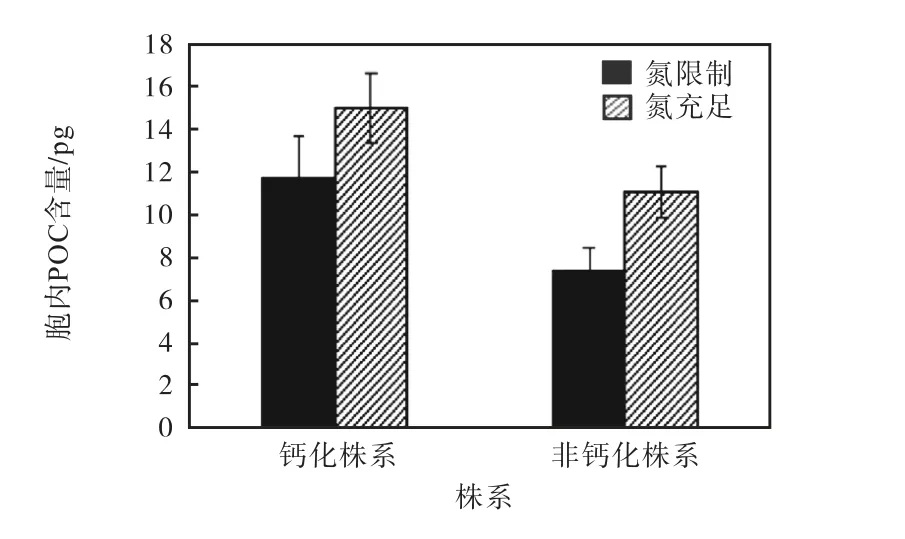
图2 钙化株系和非钙化株系的胞内POC含量 Fig. 2 Cellular POC contents of calcified strain and noncalcified strain
颗石藻钙化株系和非钙化株系的胞内PON含量如图3所示.氮限制显著降低了两个株系的胞内PON含量.其中钙化株系的胞内PON含量在氮限制的条件下相较于氮充足条件降低63.1%;非钙化株系的胞内PON含量在氮限制的条件下相较于氮充足的条件降低62.5%.在氮限制和氮充足条件下,钙化株系的胞内PON含量均高于非钙化株系.
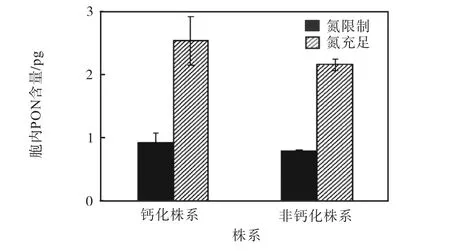
图3 钙化株系和非钙化株系的胞内 PON 含量 Fig. 3 Cellular PON contents of calcified strain and noncalcified strain
颗石藻钙化株系和非钙化株系的胞内POP含量 如图4所示.
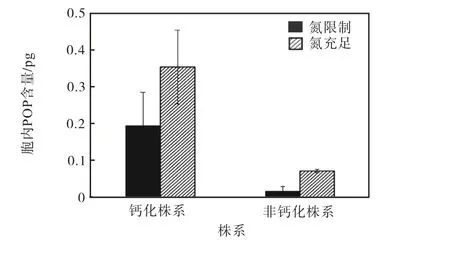
图4 钙化株系和非钙化株系的胞内POP含量 Fig. 4 Cellular POP content of calcified strain and noncalcified strain
两个株系的胞内POP含量同样受到氮限制的显著影响.钙化株系的胞内POP含量在氮限制的条件下相较于氮充足条件降低45.0%;非钙化株系的胞内POP含量在氮限制的条件下相较于氮充足的条件降低75.7%.在氮限制条件下,非钙化株系胞内POP含量相比于钙化株系低91.2%.
钙化株系和非钙化株系的细胞n(POC)/n(PON)、n(POC)/n(POP)、n(PON)/n(POP)比值见表1.
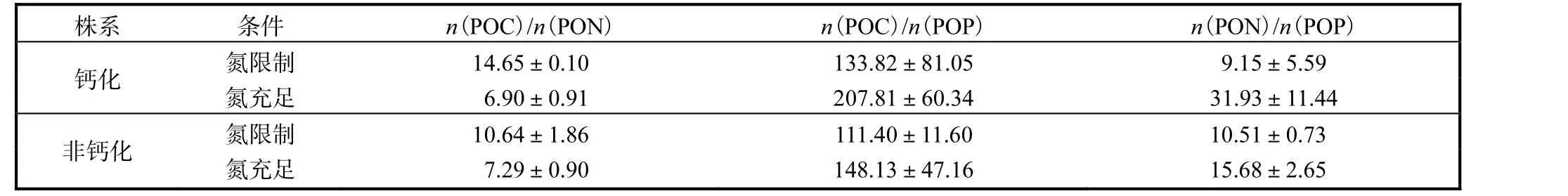
表1 钙化株系和非钙化株系的细胞n(POC)/n(PON)、n(POC)/n(POP)、n(PON)/n(POP)比值 Tab. 1 n(POC)/n(PON),n(POC)/n(POP),n(PON)/n(POP)of calcified strain and non-calcified strain
两株藻细胞的n(POC)/n(PON)在氮限制条件下升高,n(PON)/n(POP)及n(POC)/n(POP)则出现下降.钙化株系的细胞n(POC)/n(PON)比值在氮限制的条件下相较于氮充足条件升高112.2%;非钙化株系的细胞n(POC)/n(PON)值在氮限制的条件下相较于氮充足条件升高46%.氮限制条件下,非钙化株系细胞n(POC)/n(PON)值与钙化株系相比,下降了27.4%.钙化株系的细胞n(POC)/n(POP)值在氮限制的条件下相较于氮充足条件下降低35.6%;非钙化株系的细胞n(POC)/n(POP)值在氮限制的条件下相较于氮充足条件降低24.8%.氮限制条件下,非钙化株系细胞n(POC)/n(POP)比值相较于钙化株系降低16.8%;钙化株系和非钙化株系两种株系细胞的n(PON)/n(POP)值在氮限制条件下均明显低于氮充足处理.与n(POC)/n(PON)、n(POC)/n(POP)不同的是非钙化株系细胞n(PON)/n(POP)值相比于钙化株系上升了14.9%.钙化株系细胞的n(PON)/n(POP)值在氮限制条件下相较于氮充足条件降低71.3%;而非钙化株系细胞的n(PON)/n(POP)值在氮限制的条件下相较于氮充足条件降低32.9%.
钙化株系和非钙化株系的胞内C/Chl-a质量比如图5所示.在氮限制条件下,两种株系胞内的m(C)/m(Chl-a)值均显著高于氮充足条件.其中,钙化株系的胞内m(C)/m(Chl-a)值在氮限制条件下相较于氮充足条件升高476.9%;非钙化株系的胞内m(C)/m(Chl-a)值在氮限制下相较于氮充足处理组升高589.1%.与钙化株系相比,非钙化株系胞内的m(C)/m(Chl-a)值升高14.3%.
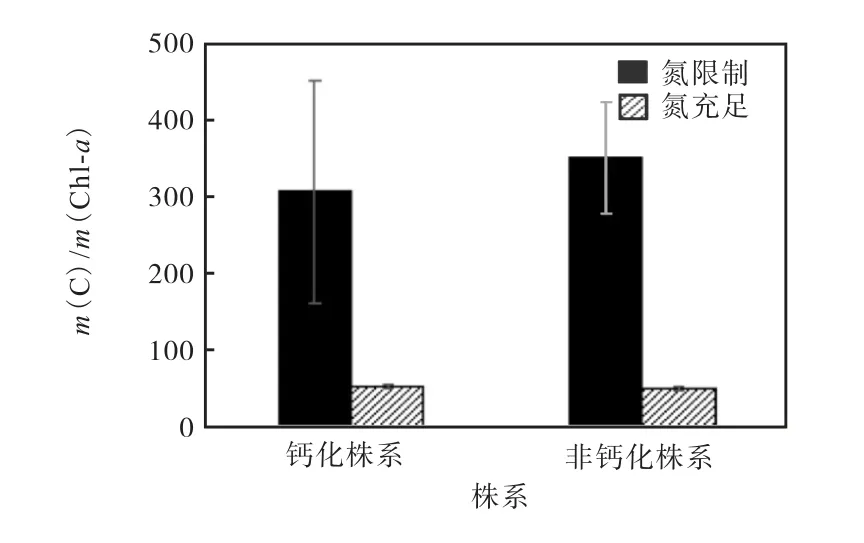
图5 钙化株系和非钙化株系的胞内C/Chl-a质量比 Fig. 5 m(C)/m(Chl-a)of calcified strain and noncalcified strain
2.3 沉降速率
钙化株系和非钙化株系的细胞沉降速率如图6所示.
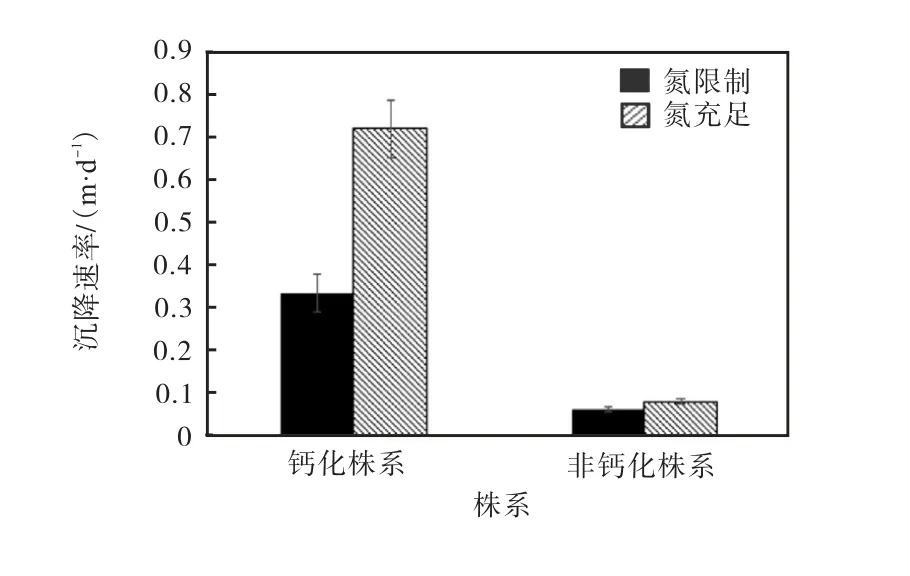
图6 钙化株系和非钙化株系的细胞沉降速率 Fig. 6 Sinking rates of calcified strain and non-calcified strain
氮限制显著降低了两个株系的沉降速率(P<0.05).其中钙化株系的沉降速率在氮限制的条件下相较于氮充足条件降低54.0%;非钙化株系的沉降速率在氮限制的条件下相较于氮充足的条件降低24.7%.在氮限制和氮充足条件下,钙化株系的沉降速率均高于非钙化株系.
根据t检验分析结果得出氮限制对钙化株系与非钙化株系两株系细胞的Chl-a含量、PON含量、n(POC)/n(PON)、沉降速率均有显著性影响,氮限制对非钙化株系的POP含量、m(C)/m(Chl-a)有显著性影响.
3 讨论
本实验结果表明,硝酸盐浓度的变化对颗石藻两株系的胞内元素组成起到重要的调节作用.与氮充足条件相比,氮限制条件使钙化株系和非钙化株系两株系细胞的Chl-a含量、PON含量及沉降速率均显著降低[34].其中,两株系藻的胞内POP含量受氮限制影响均大于各自的胞内POC含量,其主要原因是藻细胞在生长条件较差的环境下会优先进行固碳作用,从而在氮限制的条件下两株系藻的胞内POP含量相较于氮充足条件下的变化量大于各自的胞内POC含量.核酸与蛋白质等生物大分子是细胞中重要的生命物质,而氮是合成这些生物大分子的生源要素.与其他营养盐(如磷限制)相比,氮限制对颗石藻的生长、光合和钙化速率的影响尤为明显[15].这是因为氮限制使得藻细胞中一些作为光合作用重要转运体的蛋白质会严重匮乏[35–36],细胞在合成核酸与蛋白质等生物大分子过程中受阻,从而导致合成光合色素的能力也减弱,影响藻类细胞的光合固碳能力[37–38].颗石藻E. huxleyi的钙化和非钙化株系对氮限制的响应显示出了明显的株间特性.在氮充足条件下,钙化株系的胞内POC及PON的含量均略高于非钙化株系.氮限制对非钙化株系的胞内POC及Chl-a含量的降低幅度均高于对钙化株系的影响;而对非钙化株系的胞内PON降低幅度略低于钙化株系.这意味着氮限制对停止钙化作用的非钙化株系的叶绿素合成和固碳速率影响要高于对钙化株系的影响.相比于对胞内POC合成的削弱,氮限制对两株系的胞内Chl-a及胞内PON合成抑制作用更大.这导致了两株系的n(POC)/n(PON)和m(C)/m(Chl-a)比值均在氮限制条件下有较大幅度升高.在氮限制的条件下,两株系的n(POC)/n(PON)、n(POC)/n(POP)值都分别大于Redfield比值106/16和106/1,钙化株系的n(POC)/n(PON)、n(POC)/n(POP)值相比于非钙化株系还要更高一些.其主要原因可能是钙化株系在氮限制的条件下会优先进行光合固碳生成POC,由于硝酸盐的匮乏,导致了其蛋白质、核酸等生物大分子的合成受到影响,降低了对营养盐的吸收与有机氮、有机磷的合成,导致胞内PON、POP含量也随之降低[39].
值得注意的是,氮限制显著降低了两株系颗石藻的沉降速率,而且对钙化株系降低幅度显著高于非钙化株系.这主要由其对钙化株系颗石藻的钙化作用削弱引起的[27].与其他环境因子相比,氮限制也会通过对钙化作用相关的功能蛋白合成的影响而显著降低颗石藻的钙化速率[15,39].由于颗石粒的压重效应是颗石藻向下沉降的主导因素,其钙化作用的削弱会引起沉降速率的降低.另一方面,浮游植物的有机碳生产也通常会受到氮限制的削弱[40].这意味着在氮限制的环境中,颗石藻相关的雨率(PIC与POC的质量比)及由颗石粒的压重效应引起的由表层海洋向深海的碳输出均会受到显著削弱[1].
在预测未来全球气候变化下的海洋环境中,二氧化碳浓度升高导致的温室效应会引起冰川融化和降雨模式的改变,从而增强海水分层、改变海洋混合层深度,混合层深度变浅将会削弱从深海向表层海水的营养盐补充,寡营养盐海域也会逐渐扩张[1,41].海洋颗石藻主要生活在混合层以上的表层水体中,很容易受到环境变化的影响,因此,为了更加准确地预测海洋颗石藻在全球气候变化下海洋颗石藻的生理学响应以及该响应对海洋生物地球化学的反馈,氮限制的研究是其中重要的一环.此外,在全球变化背景下,其他环境因子与氮限制对颗石藻生理过程的交互效应的影响也不容忽视[27–28].
4 结语
通过对稳定生长条件下的颗石藻E. huxleyi钙化株系和非钙化株系进行恒化连续培养,实验结果显示:氮限制降低了两株系颗石藻的胞内Chl-a、细胞颗粒有机氮及颗粒有机碳含量,非钙化株系胞内各元素含量则相较于钙化株系降低的幅度更大.氮限制明显升高了两株系颗石藻的细胞n(POC)/n(PON)及n(POC)/n(POP)比值,而降低了n(PON)/n(POP)及m(C)/m(Chl-a)比值,相较而言,氮限制对钙化株系的元素比值影响高于非钙化株系.两株系的沉降速率在氮限制下均呈显著降低趋势,其中钙化株系降低幅度远高于非钙化株系.氮限制对颗石藻的生理指标有着重要影响,并且存在着较大的株间差异.以上结果有助于我们厘清颗石藻钙化作用的生理学作用,并为进一步认知和预测在变化的海洋环境中颗石藻这一重要的浮游植物功能群的响应机制及其生物地球化学效应提供了一定的科学依据.
