结球甘蓝的光合模型筛选与光响应特征比较
2021-12-20张德军郭惊涛李桂莲孟平红
蔡 霞,张德军,郭惊涛,左 晋,张 洁,李桂莲,孟平红
(1.贵州省农科院园艺研究所,贵阳 550006;2.贵州省农业科学院,贵阳 550006;3.贵州省生态气象和卫星遥感中心,贵阳 550002)
结球甘蓝(Brassicaoleracea)是十字花科芸薹属植物,起源于地中海至北海沿岸,16世纪至19世纪下半叶的300多年间引入中国,是东北、华北、西北等较冷凉地区春、夏、秋季的主栽蔬菜,长江流域及其以南地区秋、冬、春季也大面积栽培。结球甘蓝含有丰富的叶酸、维生素C、胡萝卜素等成分,具有较高的营养价值。
植物的生长发育被多种环境因子所影响,其中包括光、温度、水分等。在这些因子中,光具有特殊重要的地位。光不仅是植物光合作用能量的重要来源,同时还以环境信号的形式作用于植物,来调节植物的生长发育进程。光合作用为植物的生长发育提供必要的营养物质。通过研究植物在田间种植条件下的光合特性,探索其高产的光合生理依据,为生产上环境调控提供数据支撑。目前常用的模型有非直角双曲线模型、直角双曲线模型、叶子飘新模型。不同植物对光照强度变化的响应方式差异较大,其适应的光响应模式不一,不同模型对同一植株光响应的拟合也存在差异。近年来,研究结球甘蓝光合特性的报道鲜见,大多是以光合作用性状指标来衡量各处理的优劣,因此,本研究也为完善结球甘蓝光合特性研究提供一个案例。现选用贵州省农科院园艺研究所的结球甘蓝(BrassicaoleraceaL.var.capitataL.)作为研究试材,通过使用光合测定设备人工控制光强,测量同一时期下不同甘蓝品种的叶片光响应,运用不同的数学模型进行拟合,筛选出适合模拟甘蓝叶片光响应的模型,得到其叶片的光饱和点(LSP)、光补偿点(LCP)、最大净光合速率(Pnmax)、暗呼吸速率(Rd)等指标,以此分析叶片光合适应能力,为利用贵州生态环境进行高效育种和发展甘蓝种植提供理论依据[1-3]。
1 材料和方法
1.1 材 料
供试材料为贵州省园艺研究所创制的结球甘蓝材料22718 A 3、52019 C、G 9 B、D 56 A 1、D 68 A 2(文中简写为g 1、g 2、g 3、g 4、g 5,详见表1)。

表1 不同结球甘蓝材料品种描述Table 1 Description of different common head cabbage varieties
1.2 方 法
选择3月中旬晴朗天气,在09:30—11:30时、16:00—18:30时测定。采用美国LI-COR公司Li-6400 XT,红蓝光源设定光合有效辐射梯度2 000、1 800、1 500、1 200、1 000、700、500、200、100、50、20、0 μmol·(m2·s)-1。
在光响应曲线测定前,使用光合仪内置光源,将甘蓝叶片置于1 500 μmol·(m2·s)-1光强下诱导20 min。为保持测定环境的稳定性,设定CO2浓度为400 μmol·(m2·s)-1,测定时化学药品干燥管调节至Scrub位置。改变光照强度后,最少稳定时间设定为3 min,当测量结果变异率<0.05时,由仪器自动记录。选择甘蓝第二层展开叶片,取3株光合数据的平均值。
1.3 数据处理
利用Excel 2011软件进行数据及绘图处理,SPSS 20.0软件进行数据方差分析及光响应过程的非线性回归分析。采用以下3 种模型进行光响应曲线拟合,模型参数初始值设定如下。
非直角双曲线模型k=0.5,Pnmax=30,α=0.05,Rd=2;直角双曲线模型Pnmax=30,α=0.05,Rd=2;叶子飘新模型α=0.01,β=0.000 1,γ=0.001,Rd=0.5,Ic=10。
Pn为净光合速率[μmol·(m2·s)-1];I为光量子通量密度[μmol·(m2·s)-1];Pnmax为最大净光合速率[μmol·(m2·s)-1];Rd为暗呼吸速率[μmol·(m2·s)-1];k为非直角双曲线的凸度,0≤k≤1,凸度越大,曲线的弯曲程度越大;α为初始量子效率;β为修正系数;γ=α/Pnmax;AQE为表观量子效率,通过对I≤200μmol·(m2·s)-1的光响应数据进行直线回归,直线回归方程的斜率即为AQE。
1.3.1非直角双曲线模型[4]
(1)
(2)
(3)
1.3.2直角双曲线模型[5]
(4)
(5)
光饱和点LSP估算方法同非直角双曲线模型。
1.3.3叶子飘新模型[6-7]
(6)
上式中当光照达到光补偿点I=Ic时,Pn(LCP)=0,所以,式(6)可变换为
(7)
(8)
(9)
(10)
2 结果与分析
2.1 不同光强对结球甘蓝叶片光合速率的影响
从表2可以看出,各个品种在不同光强影响下,其净光合速率显示出了相似的正相关趋势;在同一光强下,各品种净光合速率表现出了一定的差异,光合有效辐射在700~2 000 μmol·(m2·s)-1范围内各品种间净光合速率未表现明显差异,净光合速率在光合有效辐射为700 μmol·(m2·s)-1以下则开始表现差异,品种g 5在弱光环境下的净光合作用与其他4个品种表现出显著差异。
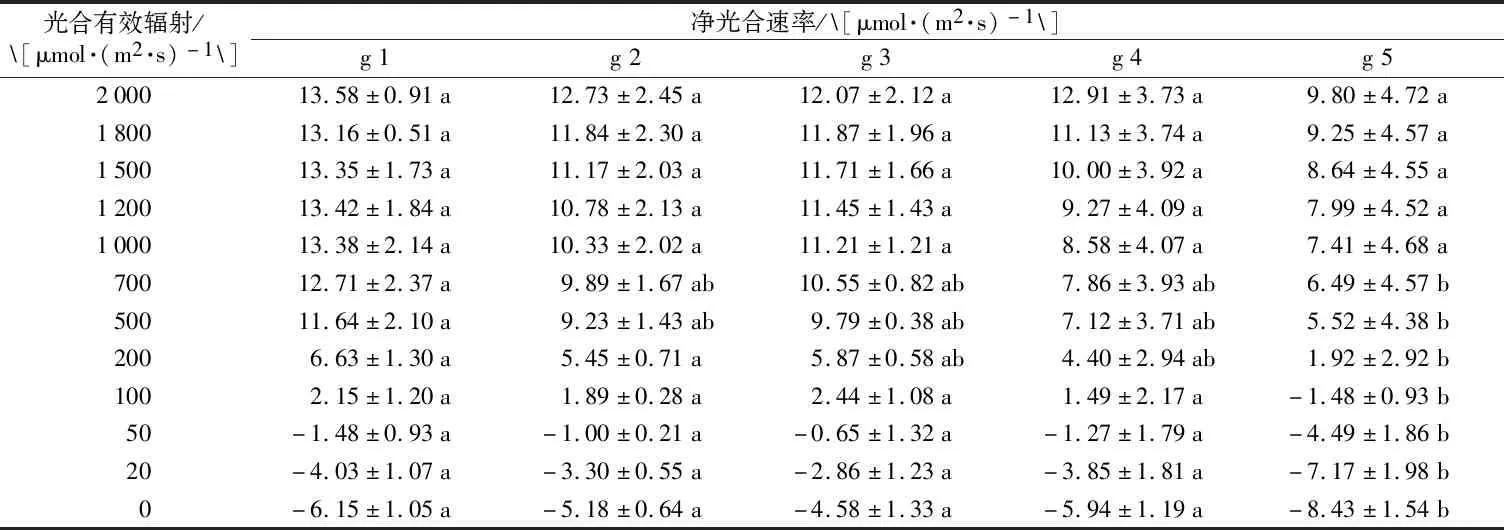
表2 不同光强对叶片净光合速率的影响Table 2 Effects of different light intensity on net photosynthetic rate of leaves
2.2 不同结球甘蓝品种光响应模型拟合效果
从图1~图3可以看出,直线双曲线模型和非直角双曲线模型拟合曲线,整体呈现出两段上升区间,第一段为叶片净光合速率在光合有效辐射为0~500 μmol·(m2·s)-1范围内出现了急速上升的趋势,光合有效辐射为500~2 000 μmol·(m2·s)-1范围内上升速度变慢,期间并未出现峰值,无法计算其光饱和点,因此通过直线双曲线模型和非直角双曲线模型无法准确拟合甘蓝光响应变化。而通过叶子飘新模型的拟合曲线可以看出拟合曲线整体仍然呈现出两段上升区间,不同之处在于5个品种拟合曲线均在光合有效辐射为1 000~2 000 μmol·(m2·s)-1时出现了峰值,结合表3计算数据发现,叶子飘新模型拟合的各项指标与实测值最为接近[8-11]。

图1 直角双曲线模型模拟光响应曲线Fig.1 Simulated light response curve based on rectangular hyperbolic model
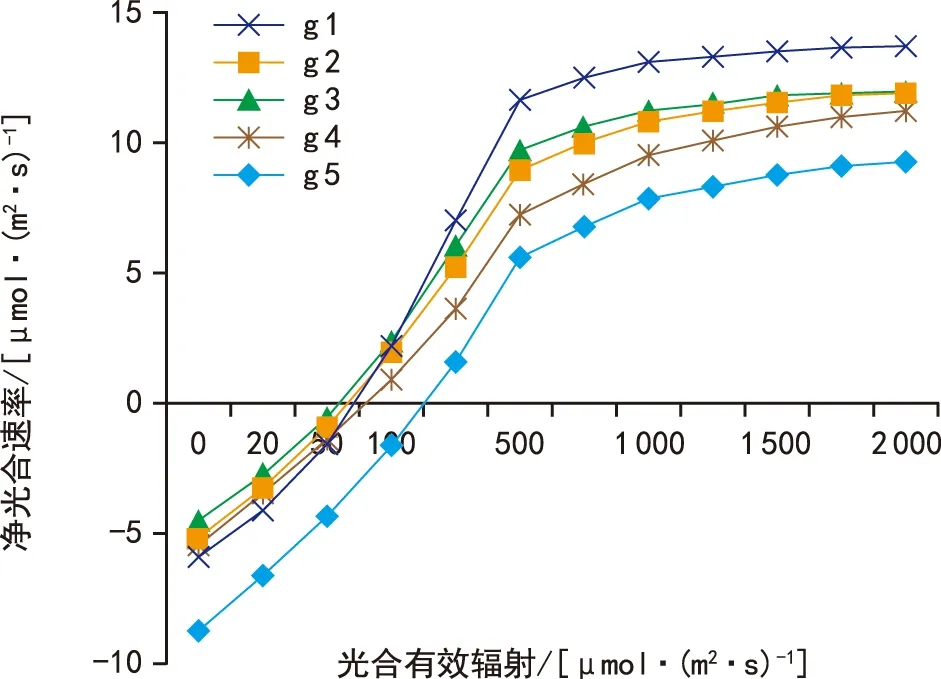
图2 非直角双曲线模型模拟光响应曲线Fig.2 Simulated light response curve based on non-rectangular hyperbolic model
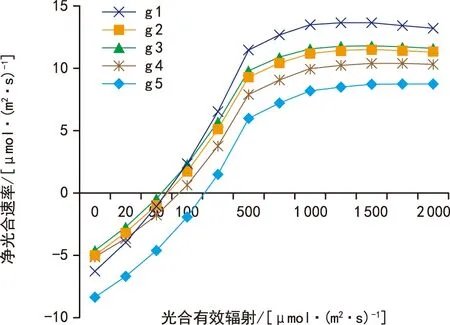
图3 新模型模拟光响应曲线Fig.3 Simulated light response curves based on Yezipiao model
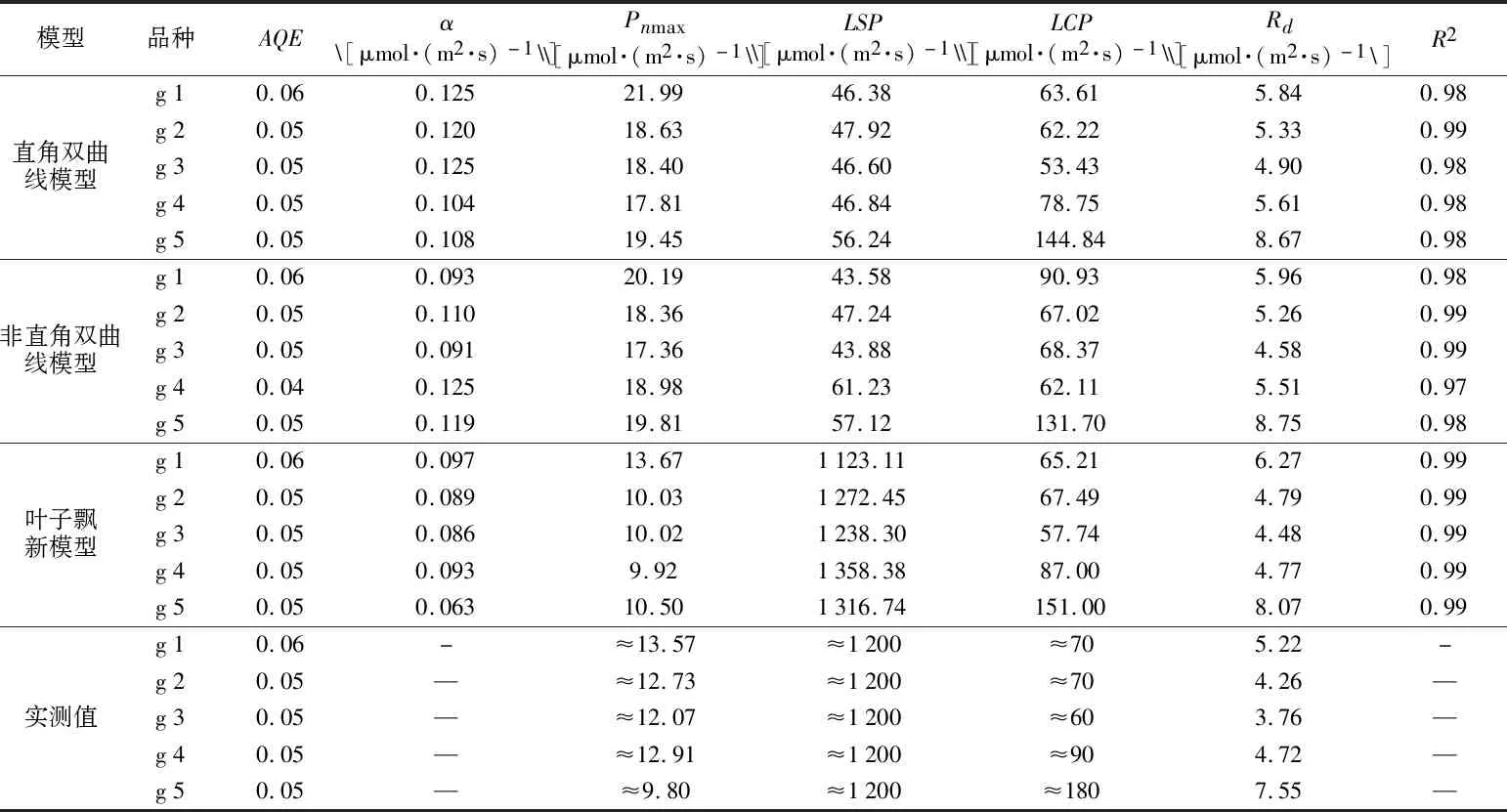
表3 Pn光响应参数实测值与模型拟合值Table 3 Comparasion of measured values and model fitting values of light response parameters of Pn
2.3 光响应特征参数
表观量子效率AQE通过3种模型拟合,与实测值几乎一致,未表现出差异。初始量子效率α值均在0.06~0.15之间。最大净光合速率Pnmax实际测量值在9.8~13.57 μmol·(m2·s)-1之间,在模拟过程中,通过非直角双曲线模型、直角双曲线模型两种模型拟合曲线所得最大净光合速率Pnmax均远大于实际测量值,叶子飘新模型拟合值与实测值最为接近。光饱和点LSP估计值在1 200 μmol·(m2·s)-1左右,新模型拟合5个品种甘蓝光饱和点分别为1 132.11 μmol·(m2·s)-1,1 272.45 μmol·(m2·s)-1,1 238.30 μmol·(m2·s)-1,1 358.38 μmol·(m2·s)-1,1 316.74 μmol·(m2·s)-1,与估计值接近,非直角双曲线模型与直角双曲线模型拟合值误差超过1 000 μmol·(m2·s)-1。光补偿点LCP与暗呼吸速率Rd在3种模型拟合值与估计值相差不大[12-15]。
3 结 论
在不同光强照射下,不同品种甘蓝叶片净光合速率出现了不同程度的差别,尤其在光合有效辐射为700 μmol·(m2·s)-1以下,品种g 5与其他4个品种的净光合速率呈现显著差异,表明在弱光环境下,不同品种对光的利用程度各有差异。
对比非直角双曲线模型、直角双曲线模型和叶子飘新模型的拟合结果可以看出,初始量子效率均在0.06~0.15之间,接近理论值,拟合方法是合理的。5个品种甘蓝光饱和点非直角双曲线模型与直角双曲线模型拟合值误差超过1 000 μmol·(m2·s)-1,不宜采用其作为拟合甘蓝叶片光响应模型。叶子飘新模型拟合光饱和点与实测值接近,拟合效果最好,是拟合甘蓝叶片光响应最佳模型,利用该模型可以较好地了解甘蓝生长过程中叶片对光的需求特性。
结球甘蓝的5个品种中,暗呼吸速率的拟合值从小到大的排序为:g 3、g 4、g 2、g 1、g 5;光饱和点的拟合值从小到大的排序为:g 1、g 3、g 2、g 5、g 4;光补偿点的拟合值从小到大的排序为:g 3、g 1、g 2、g 4、g 5;而最大净光合速率的拟合值从小到大的排序为:g 4、g 3、g 2、g 5、g 1;初始量子效率拟合值从小到大的排序为:g 5、g 3、g 2、g 4、g 1。
4 讨 论
利用3种光合模型对5个结球甘蓝品种的光强-光合响应曲线进行拟合,为保证拟合光响应特征参数的准确性,需要对不同光响应模型的适用性进行分析。可以通过计算决定系数R2的大小来验证方程拟合的精度,R2越大,模型的精确度越高,拟合数值的准确性越高[16]。在3个光响应曲线模型中,直线双曲线模型和非直角双曲线模型拟合曲线,整体呈现出两段上升区间,期间并未出现峰值,无法计算出其光饱和点,而叶子飘新模型的拟合曲线整体仍然呈现出两段上升区间,但5个品种拟合曲线均在光合有效辐射为1 000~2 000 μmol·(m2·s)-1时出现了峰值,且叶子飘新模型拟合的各项指标与实测值最为接近,所以叶子飘新模型是结球甘蓝的最适光合模型。
直角双曲线、非直角双曲线模型只能拟合出一条没有极值的渐近线, 拟合得到的最大净光合速率远大于实测值,而且这2种光合模型也不能准确地描述出净光合速率随光强增加而下降的光响应曲线。叶子飘新模型可以很好地解决这一问题。该模型是一条有峰值的曲线,可以准确地拟合出最大净光合速率和光饱和点等光响应特征参数,同时可以用它来描述植物在光强超过饱和光强时的光响应曲线[17-18]。该模型自改进以来就被广泛应用。张赟齐等[19]研究无患子的光合模型时,得出叶子飘新模型是拟合无患子光响应曲线的最佳模型。刘子凡等[20]也认为,叶子飘新模型能更好地拟合木薯光合响应曲线,得到的光合响应参数也比较准确。
光响应曲线是衡量植物光合作用的重要指标。不同品种结球甘蓝的光响应曲线趋势基本一致,在低光照范围内[PAR≤500 μmol·(m2·s)-1],各品种结球甘蓝的净光合速率随光强的增加而迅速上升,光合作用对低光照敏感响应。随光强持续增强,各品种结球甘蓝的净光合速率缓慢增加至最大值,达到最大净光合速率,该点的光强即为光饱和点,并且,因为结球甘蓝的品种不同,光补偿点、光饱和点、初始光能利用率值和最大净光合速率也不同。当光强达到光饱和点时,净光合速率不再随着光强的增加而增大,反而出现下降现象,造成光抑制。光抑制是植物进化出的光系统保护机制,以消散过度吸收的太阳能,从而保护光合结构。
最大净光合速率是反映植物叶片光合的潜力。光饱和点和光补偿点直接反映了植物对光的需求和利用能力[21]。本研究中,g 3的光补偿点最低,表明了其对弱光的利用能力较强;g 4的光饱和点最高,其对太阳能的利用率也较高,生产潜力较大。
本研究5个结球甘蓝品种光饱和点的高低与净光合速率大小并未呈现绝对一致性,说明植物最大净光合速率的大小不仅仅取决于光饱和点的高低,还要受到其他因子的共同影响。另外,本研究仅选择结球甘蓝抽薹期进行光响应特征对比,其他生长时期的光合参数还有待进一步研究。
