人格特质及脑功能连接对社交网络的影响*
2021-12-10李依蔓庄恺祥霍腾宾徐鹏飞罗跃嘉3
李依蔓 刘 程 庄恺祥 霍腾宾 徐鹏飞 罗跃嘉3,4,,6 邱 江,2
人格特质及脑功能连接对社交网络的影响
李依蔓刘 程庄恺祥霍腾宾徐鹏飞罗跃嘉邱 江
(西南大学心理学部,认知与人格教育部重点实验室, 重庆 400715)(深圳大学心理学院,深圳大学脑疾病与认知科学研究中心, 深圳 518060) (北京师范大学心理学部, 北京 100875)(齐鲁师范学院教师教育学院, 心理学与认知神经科学研究所, 济南 250200)
采用网络分析的方法, 本研究从个体受欢迎程度和个体间亲密程度两方面探究了人格特质对社交网络的影响, 并在此基础上进一步探究了个体间大脑静息态功能连接相似性和社交网络的关系。结果发现:(1)高尽责性的个体在需要“值得信任”特质的社交网络中更受欢迎, 高宜人性的个体在需要“共享时光”的社交网络中更受欢迎; (2)在需要“相同兴趣”特质的社交网络中, 个体间人格相似性和社会距离呈显著负相关关系; (3)同样在需要“相同兴趣”特质的社交网络中, 个体间部分功能连接相似性与社会距离呈显著负相关关系, 这些功能连接主要集中在额顶控制网络以及背侧注意网络; 同时, 部分节点功能连接相似性与社会距离呈显著正相关关系, 这些功能连接主要集中在默认网络。研究结果揭示了人格特质对不同社交网络结构的影响, 以及个体间人格特质相似性和静息态脑网络相似性与社会距离的关系。本研究对理解社交网络的结构, 形成规律以及其中的信息传播规律有着重要启示意义。
社交网络, 人格特质, 静息态功能连接
1 引言
人类在本质上是一个社会性物种, 没有大规模的集体行为, 人类的文明不可能存在(Borgatti et al., 2009)。在今天这个信息传递极其便捷的时代, 人际关系对人们生活的影响不容小觑。了解群体人际关系的结构以及形成规律可以帮助我们更好地了解个体的行为模式并预测个体的行为。目前, 人际关系的研究主要基于单个个体, 讨论了个体的人际交往能力或社交意愿等, 但是从群体角度切入讨论社交关系结构的研究较少, 而且对于同一集体内不同社交需求的关注也甚为少见。
社会世界的结构是极其复杂的, 涉及到多个相互作用的单元。理解这些单元是何时、如何以及为什么相互作用的关键在于找到理论和方法来解决这种错综复杂的人际连接模式(Borgatti et al., 2009)。网络科学提供了一种很好的理论和方法。将网络科学应用到人际关系的研究中来, 那么每一个个体就是网络中的一个节点, 人与人之间的联系就是节点之间的边。网络科学可以捕捉到人际关系中丰富的连接模式, 甚至精确定位那些在人际交往中有着影响全局能力的个体, 为研究人际关系提供了新的思路。
人格特质是个体的核心特征, 反映了个体对环境反应跨情境一致性的行为模式(Simon et al., 2020), 是讨论个体的社会行为规律时必不可少的一环。目前讨论人格特质和人际交往之间关系的研究大多从个体层面讨论了人格特质和个体人际交往能力或受欢迎程度的关系(Goldberg, 1992; Graziano et al., 2007; Graziano & Tobin, 2010; Klein et al., 2004)。研究者发现高宜人性个体更容易维持长久的人际关系(Graziano et al., 2007)。高宜人性个体被证明拥有更好的自我控制能力和情绪调节能力, 同时, 也更加擅长于在群体中调和矛盾(Graziano et al., 2007; Jensen-Campbell & Graziano, 2001)。外向性的主要特征是热情、乐群和果断, 比起内向的人, 外向性高的人倾向于参与到更多的社交活动中去, 寻求和渴望吸引更多的社会关注(Feiler & Kleinbaum, 2015)。与外向性相似, 高开放性个体也更倾向于参与到更多新的活动中去, 其中也包括社交活动, 他们更容易接受新的环境, 愿意交新的朋友, 同时也更容易接受他人的新观点(Kaplan et al., 2015)。这些研究讨论了人格特质和个体人际交往能力或社交意愿之间的关系, 证明了不同人格特质在人际交往的不同层面起着重要作用。但由于方法的局限, 大多数研究都只是针对单一个体或关系, 很少研究从群体层面入手。在人际交往能力的测量上, 这些研究大都基于主观测量问卷, 局限于个体层面, 而人际交往是一项特殊的技能, 应该更多的从他人的评价中反映出来, 因此这种测量方式可能会缺乏客观性。
社交网络的方法可以很好地解决目前人际交往研究中出现的问题。社交网络的相关研究正在逐渐增多, 越来越多的研究者认识到利用社交网络可以更好地理解人的心理和行为。社交网络中心度和社会距离两个指标分别从个体和个体之间的关系, 也就是社交网络中的节点和连边两个角度描述了社交网络的结构(Blakemore, 2008; Parkinson et al., 2018), 本研究也将用到这两个指标对社交网络的结构和规律进行深入地探索。社交网络中心度一般指以某个个体为中心, 与这个个体有直接联系的人的数量, 能够比较直观的反映个体人际交往的状况, 以及个体在社交网络中的地位(Gattis et al., 2004; Hyon et al., 2019)。有研究证明了神经质与社交网络中心度有着密切的关系, 神经质得分越低, 个体就越可能处于社交网络的中心位置(Klein et al., 2004)。Landis (2016)等人的研究证明了外向性高的人在社交网络中的地位更高, 他们往往更容易吸引同龄人的注意, 并且更容易参与到社交活动中去。但也有研究表明, 社交网络中心地位和外向性之间并没有显著相关, 反而和尽责性和宜人性有显著相关(Wolfer et al., 2012)。从这些研究结果中不难看出, 即使这些研究所用来衡量人格特质的指标都是大五人格问卷, 他们的研究结果也出现了一些分歧。目前, 社交网络中心地位的相关研究一般集中讨论的是单一人格特质(如外向性)与单一网络(如友谊网络)中的中心度之间的关系。但是, 在实际生活中, 同一个人往往同时存在于不同类型的关系定义的多个网络中。个体可能会向一些人寻求建议, 向另一些人寻求陪伴, 再向另一些人寻求情感支持(Morelli et al., 2017)。这意味着一个人可以在一种类型的网络中占据核心地位, 但在另一些类型的网络中处于边缘位置。社交网络多样性是目前社交网络研究中被普遍忽视的问题。另一方面, 以往社交网络相关研究大都是从网络中心度入手, 较少研究探讨社会距离的影响因素。但在网络研究中, 除了节点, 网络中连接节点的边, 也就是人与人之间的关系也能提供大量有用的信息, 有助于更好的了解人际关系建立的规律以及社交网络的整体结构。在社交网络中, 社会距离被用来表征个体间的亲密程度, 通过最短路径长度的算法获得。这个指标可以衡量每两个个体之间的人际距离, 是依靠已经建立好的无向社交网络得到的。目前讨论人格特质和个体间关系的研究还比较少, 主要集中在对友谊选择的人格特质同质性和异质性的争论上(Feiler & Kleinbaum, 2015; Maarten et al., 2007; Massen & Koski, 2014; Selfhout et al., 2010)。友谊选择的同质性是指个体更倾向于选择和自己相似的人成为朋友, 异质性则相反(McPherson et al., 2001)。荷兰的一项研究发现同性青少年朋友间倾向于在外向性和宜人性中表现出相似性(Maarten et al., 2007)。除此之外, 相似的外向性和人际关系之间的联系也在配偶选择、婚姻压力的相关研究中被证明了(Furler et al., 2013; Gattis et al., 2004; Humbad et al., 2010; Maarten et al., 2007)。综上所述, 虽然目前已有部分研究用社交网络的方法讨论了人格特质和人际交往的关系, 但较少研究从个体间的关系入手, 并且社交网络的多样性也被普遍忽视了。
在讨论社交网络时, 大脑也是不可忽视的一部分。大脑功能和结构为人类行为提供了生物学机制, 而社会网络则为这些行为提供了外部诱因(Falk & Bassett, 2017)。人格体现了人们在感知、解释和对周围环境的反应中的共性, 并且前面已经提到有研究证明了人格的相似性和社会距离的关系。既然人格可以被认为体现了个体认知过程中的共性, 那么认知过程中个体间大脑响应的相似性可能也与社会距离有关。有研究者用功能性磁共振成像的方法回答了个体间认知过程反应的相似性与社会距离的关系这一问题(Parkinson et al., 2018), 发现当受试者在观看电影时朋友间的神经反应异常相似, 并且相似度随着社会距离的增加而降低。这个结果表明, 朋友间感知和应对周围世界的大脑反应是相似的。目前计算个体间脑间相似性的方法在社交网络中的应用还非常少, 而且主要集中在任务态的脑影像研究中。在最近的研究中, 有研究者已经计算了个体间静息态功能连接(Functional Connectivity, FC)的相似性作为个体间脑连接模式相似性的指标来开展研究(Jaaskelainen et al., 2016; Nummenmaa et al., 2012)。这些研究结果证明了静息态脑网络的相似性也同样可以有力的表征个体间大脑的相似性。另一方面, 现有研究中, 比起任务态脑影像研究, 更多与人格有关的脑影像研究都采用了静息态和结构态的脑影像作为主要研究对象(Simon et al., 2020; Wu et al., 2019), 证明了许多特定脑区和人格特质之间的关系, 便于我们从人格特质的角度来解释社交距离和大脑相似性之间的关系。
综上所述, 虽然已经有不少研究讨论了人格特质和人际交往的关系, 但只有很小一部分从社交网络的角度入手, 更少有研究同时在一个社交群体内建立多个社交网络, 讨论人格特质对不同社交网络的影响。然而, 在人们的实际生活中, 即使是在同一集体中, 因为社交需求的不同也会同时存在多个不同的社交网络, 忽视这一点可能会导致我们过早的对人格特质和人际交往之间的关系下定论, 这也是很多研究结果出现矛盾的原因之一。其次, 大多数研究者都从个体特质入手讨论人格特质对人际交往的影响, 而忽视了个体间关系和人格特质之间的关联, 而在人际交往中, 个体间关系往往能反映出大量信息。因此, 对社会距离的讨论是全面了解社交网络的结构不可或缺的一环。最后, 将大脑功能和结构与社会网络中的活动和结构联系起来也是十分必要的, 这种方法不仅可以提高预测行为的能力、推导出核心的心理学和神经认知原理, 也能帮助区分大脑健康和疾病, 来提高对个体的认识。因此, 在此基础上, 本研究提出三项假设:(1)在同一群体中, 根据社交需求存在不同的社交网络, 并且个体在不同社交网络中的地位也不尽相同, 会受到不同人格特质的影响; (2)个体间人格特质越相似, 个体间社会距离越短; (3)个体间静息态脑功能连接相似性越高, 个体间社会距离越短。为了验证以上假设, 本研究将采用网络科学的方法, 探讨人格特质对个体在社交网络中的中心地位和个体间的社会距离的影响以及个体间大脑静息态功能连接相似性对社会距离的影响。
2 方法
2.1 被试
以往研究表明, 社交网络和个体特质的回归分析达到显著性水平(0.05)时的效果量一般为0.1~0.3 (Morelli et al., 2017), 以此为基础, 采用G*Power 3.1.9.2软件进行分析, 结果显示若要达到0.8的统计检验力(α = 0.05)需要25~92名被试。为了在同一个群体内建立多个社交网络, 本研究一共招募了118名被试完成了社交网络问卷。不同于一般的数据采集, 为了得到更加客观有效的数据, 构建完整真实的社交网络, 本研究的所有被试均来自某学院同一个年级, 占该学院该年级总人数的三分之二。并且, 社交网络的采集时间是在被试入学后14个月左右, 此时被试之间已经有了基本的了解, 形成了稳定的社交网络, 更加便于我们讨论稳定的人际关系。其中有94人完成了大五人格的测试, 纳入了最终的行为数据的分析(19男, 平均年龄19.30 ± 0.80岁)。其中有两名被试并未参加磁共振扫描, 因此静息态脑影像数据的分析只纳入了92名被试(19男, 平均年龄19.29 ± 0.76岁)。另外, 为了讨论不同社交网络的意义, 招募了167名被试完成了针对不同社交网络友谊选择原因的问卷。被试全部为在校大学生(26男, 平均年龄19.32 ± 0.65岁)。所有被试均自愿参加试验并签署了知情同意书, 无精神或神经疾病史, 无生理疾病。
2.2 数据采集
2.2.1 社交网络的构建及人格特质的测量
社交网络的建立采用了问卷的形式, 共测量了118名被试在不同社交需求下的社交网络。问卷一共有9个问题, 要求被试根据问题提示在全年级范围内相互提名(如:“谁是你很亲近朋友?”, 完整问卷见网络版附录)。在每一个问题下, 全年级所有学生的姓名按班级分类并按姓名首字母排序进行了罗列, 被试需根据以上9个问题在这些姓名中进行勾选。问卷是由在PNAS上发表的一篇文章翻译而来(Morelli et al., 2018; Morelli et al., 2017)。在他们的研究中, 此问卷也被用于测量被试的社交网络, 研究结果也证明了问卷有较高的效度(Morelli et al., 2018; Morelli et al., 2017)。
为了测量个体的人格特质, 94名被试完成了大五人格特质问卷(Costa & McCrae, 1992), 该问卷基于大五人格理论, 对人格的五个维度进行测量, 分别是:神经质、外向性、开放性、宜人性及尽责性。问卷内含60个项目, 每个维度有12个项目。所有题目的选项从“非常不符合”到“非常符合”, 全部为5点计分, 部分题目需要反向计分。
为了探究社交网络的内在含义, 即个体在友谊选择时的原因, 167名被试完成了友谊选择维度重要性问卷, 测量了被试在不同社交网络中进行友谊选择时对不同因素的侧重, 该问卷也是由上述社交网络问卷所参考的文章翻译而来(Morelli et al., 2018; Morelli et al., 2017)(如:以“亲近朋友”社交网络为例, 与这个网络相对应的问题是:“当你和某人成为亲近朋友时, 多大程度上是因为以下几点原因?”)。被试需要在以下几个选项上滑动进度条进行“0~100”的评分:“你信任他们”; “他们与你有相同的兴趣, 态度和价值观”; “你们在情感上理解对方”; “他们能提供给你所需要的信息”; “他们能在你的学业或事业上帮助你”; “他们能帮你认识新的有趣的人”; “你们经常在一起做一些有趣的事, 玩的很开心”。“0”代表当被试和某人成为亲近朋友时, 完全不考虑这个因素, “100”代表这个因素非常重要。
2.2.2 静息态数据采集与预处理
静息态功能性磁共振影像数据均采集于西门子扫描仪(3.0T Siemens Trio MRI)。扫描过程中, 所有被试被要求闭上眼睛, 平躺休息, 不做思考和回忆某些特定的事件, 但需要保持清醒。在正式实验前, 每名被试都接受了基于DSM–IV的结构化访谈, 以确保每名被试都没有物质滥用史、神经系统疾病或认知障碍史。所有被试均被告知了研究包含的内容, 且都签署了书面知情同意书。在每次实验结束后, 被试将获得一定金额的报酬。该项目获得了伦理审核批准。静息态扫描使用梯度回波EPI序列(GRE–EPI), 8分钟内连续采集242个全脑静息态3D影像。扫描参数为:回波时间(Echo Time, TE) = 30 ms; 重复时间(Repetition Time, TR) = 2000 ms; 翻转角(Flip Angle, FA) = 9°; 矩阵大小(Matrix) = 64×64; 视野大小(Field of Vision, FOV) = 192 mm × 192 mm; 层数(Slice) = 32; 厚度(Thickness) = 3 mm; 层间距(Distance between Slice) = 1 mm; 体素大小(Voxel Size) = 3.4 mm × 3.4 mm × 4 mm。
静息态数据预处理:(1)为保证信号稳定, 删除fMRI时间序列上的前10个数据点; (2)对留下的232个EPI数据采用sinc函数插值进行时间层校正, 并根据相邻EPI数据的位移进行头动校正; (3)采用EPI模板, 将数据从原始空间映射到MNI空间(Montreal Neurological Institute (MNI)space), 体素大小为3 mm × 3 mm × 3 mm; (3)去除线性趋势, 并将全脑信号、脑脊液信号、白质信号及12个头动参数作为协变量进行回归; (4)滤波(0.01~0.1 Hz)以去除低频信号漂移及高频生理噪声; (5)采用4 mm平滑核进行平滑。上述处理均采用DPABI工具包(http://rfmri.org/DPARSF)。
2.3 数据分析
2.3.1 社交网络中心度的计算和分析
首先根据原始数据建立了94名被试间9个不同的二进制社交网络矩阵, 其中个体间的数值为1代表两者之间有直接联系, 数值为0则代表没有直接联系。接着计算了每个被试在不同社交网络中的社交网络中心度(入度), 度越大, 代表被试越受欢迎。最后, 为了探究社交网络中心度和人格特质的关系, 以9个社交网络中心度为因变量分别进行了9次负二项回归分析(Negative binomial regression), 在每一个回归模型中, 大五人格特质的五个维度都作为自变量, 以检验并分离出每个网络中对社交网络中心度最强的预测因子, 这部分分析使用了R语言中ass包中的glm.nb功能。另外, 计算了167名被试在每个问题中每个选项上评分的均值, 作为每个社交网络中友谊选择维度的重要性指标。在Morelli等人(2017)的研究中, 也同样使用了其它样本描述统计的结果对社交网络进行了有效分类。
2.3.2 社会距离的计算和分析
为了得到最客观真实社交网络关系, 分别把8个原始二进制社交网络进行过滤, 只有双方互选的被试之间的边为1, 否则为0(这部分分析并没有将“矛盾发生”社交网络纳入其中, 因为这里讨论的社会距离反映的是个体间的亲密程度, 而这个网络中的边代表的是个体间发生过矛盾, 不符合研究意图)。接着分别计算出8个94×94的社会距离矩阵, 两个被试间有直接互选关系则社会距离为1, 若两人并无直接联系, 但能通过一个共同好友联系起来, 则两人之间的社会距离为2, 以此类推, 两人之间可以通过最少几条边建立联系, 则两人的社会距离就为几。社会距离越小, 表明两人的关系越亲近。接下来计算了个体两两间大五人格特质的欧式距离, 代表个体间总体人格的相似性, 值越大, 个体间人格差异越大, 相似性越小。
为了探究社会距离和人格差异的关系, 将个体间的总体人格差异矩阵和8个社会距离矩阵分别进行了二次分配程序相关分析(Quadratic Assignment Procedure, QAP), 试图探究总体人格差异和社会距离的关系。QAP相关分析是一种对矩阵间进行相关分析的方法, 通过对矩阵中值的比较, 给出两个矩阵的相关系数, 同时对系数进行非参数检验, 通过对邻接矩阵中的行和列进行重新排列, 将实际数据和随机排列的矩阵的相关进行比较。在这里使用了R中的sna包中的qaptest功能进行了此计算, 相关分析采用了斯皮尔曼等级相关系数, 一共进行了5000次数据置换。另外, 为了尽可能得到最真实的人格差异对人际距离的影响, 还计算了个体两两间年龄、性别、种族、利手和所在班级的差异值, 并作为协变量放入分析中。
2.3.3 静息态脑间相似性的分析
静息态数据分析:选用了经典脑功能图谱Power264 (Power et al., 2011)作为脑网络模板, 提取出264个节点的平均BOLD (Blood Oxygenation Level Dependent)时间序列, 该模板一共有264个节点, 将节点分为了14个子网络, 包括视觉网络、听觉网络、感觉运动网络、默认网络、背侧注意网络、腹侧注意网络、扣带鳃盖网络、额顶控制网络、凸显网络、记忆网络、皮质下结构和小脑。接着计算出任意两个节点时间序列的皮尔逊相关系数并将其转化为z分数, 以此来代表节点间的平均FC强度, 得到每个被试一个全脑的264×264的FC矩阵。为了得到个体间FC相似性矩阵, 在全脑水平上, 计算了个体间两两全脑FC矩阵的二次匹配程序相关系数, 得到1个92×92 (被试数)的个体间全脑FC相似性矩阵。在网络水平上, 根据264个节点的网络属性, 从全脑264×264的FC矩阵中提取出每个被试14个脑网络内的FC以及14个脑网络和除自身网络外的其它所有节点的FC分别作为网络内FC和网络外FC, 接下来, 为了得到网络水平上个体间大脑FC的相似性, 分别计算了个体间每个网络内FC的皮尔逊相关(得到14个92×92的相似性矩阵)和个体间每个网络外的FC的皮尔逊相关(得到14个92×92的相似性矩阵)。在节点水平上, 计算了个体间每个节点与其它所有节点的FC的皮尔逊相关系数, 得到了264个92×92的相似性矩阵(见图1)。
为了探究进一步探究人格相似性和社交距离的关系在个体间静息态脑网络相似性上的体现, 分别在个体间全脑水平, 网络水平, 以及节点水平的FC相似性矩阵和“亲近朋友”和“消息分享”这两个社交网络的社会距离之间进行了QAP相关分析, 采用了留一交叉验证(leave-one-out)的方法, 在每一次循环中都剔除一名被试的数据, 剩下的所有数据进行斯皮尔曼相关分析, 并且在每一次分析中都进行了5000次数据置换, 同时计算了被试间年龄、性别、民族、班级以及利手的相似性作为协变量纳入相关分析。在所有循环中都显著相关的网络或节点才纳入最终结果, 也就是说在每一次循环中都和社交距离显著相关的节点或网络才纳入最终结果。

图1 个体间静息态FC相似性和社会距离的关系分析流程
3 研究结果
3.1 社交网络中心度和人格特质的关系分析结果
在根据原始数据建立了9个二进制的社交网络后, 本研究对这9个二进制的社交网络进行了两两的二次分配程序相关分析, 试图探究不同社交网络间的相似性。结果表明, 同一群体中不同社交需求的社交网络间虽然显著相关, 但仍存在着差异, 并且不同网络间的相似性存在差异。为了检验不同人格特质对不同社交网络中心度的影响, 以9个社交网络中心度为因变量分别进行了9次负二项回归分析。结果表明, 宜人性可以显著正向预测“亲近朋友” (= 0.24, SE = 0.09,= 2.65,= 0.01)和负向预测“矛盾发生” (= –0.29, SE = 0.11,= –2.61,= 0.01)的社交网络中心度; 尽责性显著正向预测了“支持给予”社交网络的中心度(= 0.25, SE = 0.10,= 2.57,= 0.01); 神经质显著负向预测了“积极感受”社交网络的中心度(= –0.24, SE = 0.11,= –2.14,= 0.03) (见图2A–D)。
计算了167名被试在每个问题中的每个选项上评分的均值, 作为每个社交网络中友谊选择各维度的重要性。结果表明, 在不同社交网络中进行友谊选择时, 个体考虑因素的重要性有着很大的区别。在“亲近朋友”、“日常活动”和“积极感受”社交网络中, 被试友谊选择原因中“共享时光”的均值最大; “社交建议”、“寻求帮助”、“消息分享”和“支持给予”的网络中, “值得信任”的均值最大; 在“善解人意”的网络中, “情感接近”均值最大(见图3)。
3.2 社会距离和人格相似性关系的分析结果
为了探究社会距离和人格相似性的关系, 将个体间的人格差异矩阵和8社会距离矩阵分别进行了二次分配程序相关分析。结果表明, 在亲近朋友(= 0.13,= 0.03)和消息分享(= 0.10,= 0.04)这两个社交网络中, 个体间大五人格的欧式距离和其社会距离呈显著正相关关系, 个体间大五人格的差异越大, 社会距离就越远, 基本符合本研究的假设。但在其余的6个社交网络中并没有出现这种效应(见表1)。

图2 在不同社交网络中大五人格特质各维度的分布情况(A–D:颜色越深, 该人格特质得分越高, 节点越大, 节点度越大)及社交网络间相关系数矩阵(E)

图3 不同社交网络中友谊选择的考虑因素

表1 个体间整体人格差异性和社会距离的关系
3.3 社会距离和静息态脑网络相似性的关系分析结果
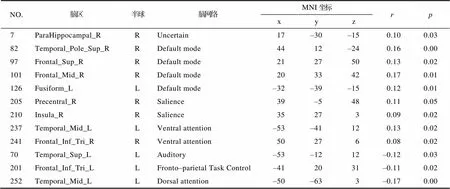
在全脑水平, 将研究二中得到的和整体人格特质的相似性显著相关的两个社交网络:“亲近朋友” “消息分享”的社会距离分别和个体间的全脑FC相似性进行了矩阵相关分析, 发现个体间全脑水平和脑网络水平的FC相似性和两个网络的社会距离并无显著关系。在节点水平, “亲近朋友”社交网络的社会距离和部分节点FC相似性呈显著正相关, 包括右侧楔前叶, 右侧额上回, 右侧额中回, 右侧扣带前回, 左侧舌回, 左侧梭状回, 右侧枕中回, 其中大部分节点都属于默认网络。同时, “亲近朋友”社交网络的社会距离也和另外一些节点的FC相似性呈显著负相关, 包括右侧额中回和右侧角回, 分别属于额顶控制网络和凸显网络。在“消息分享”这个社交网络中, 虽然和“亲近朋友”社交网络得到的节点不尽相同, 但也发现了相似的结果, “消息分享”社交网络的社会距离和部分节点的FC相似性呈显著正相关, 包括右侧海马旁回、右侧颞上回、右侧楔前叶、右侧额中回、右侧额上回、左侧梭状回、右侧脑岛、右侧三角部额下回、左侧颞中回, 其中大部分节点都属于默认网络。并且, “消息分享”社交网络的社会距离也和部分节点的FC相似性呈显著负相关, 包括左侧颞中回、左侧颞上回、左侧三角部额下回, 分别属于背侧注意网络, 听觉网络和额顶控制网络(见图4)。
4 讨论
本研究主要采用了网络科学的方法, 在同一群体内测量了不同的社交网络, 选取了社交网络中心度和社会距离两个指标探究人格特质对不同社交网络的影响, 以及个体间大脑静息态功能连接与社会距离的关系。结果发现:高尽责性的个体在需要“值得信任”特质的社交网络中更受欢迎, 高宜人性的个体在需要 “共享时光”的社交网络中更受欢迎; 在需要“相同兴趣”特质的社交网络中, 个体间人格相似性和社会距离呈显著负相关关系; 在“亲近朋友”和“消息分享”社交网络中, 个体间部分FC相似性与社会距离呈显著负相关关系, 这些FC分布在额顶控制网络, 背侧注意网络, 同时, 个体间部分FC相似性与社会距离呈显著负相关关系, 这些FC主要集中在默认网络。

图4 节点水平个体间FC相似性与社会距离显著相关的节点(A, B); 随着社会距离增大, 个体间节点以及网络水平FC相似性的变化(C, D) (红色代表和社会距离呈显著正相关的节点, 蓝色代表和社会距离呈显著负相关的节点, 节点越大, 相似性越强。为了方便比较, 此处单个节点的不同社交距离的平均脑间相似性进行了z值化处理。
原始社交网络的两两相关分析发现, 8个不同的社交网络间都呈显著正相关关系, 但不同网络间的相关程度不同。这一发现也与以往研究结果一致, 一定程度上证明了个体在建立社交网络时会有自己的偏好, 并且随着社交需求的不同而建立不同的网络(Morelli et al., 2017)。具体来说, 人格特质和社交网络中心度的分析结果发现, 在同一社交群体内确实存在着多个不同的社交网络, 且人格特质对不同社交网络中心度的影响存在着差异。宜人性显著正向预测了“亲近朋友”, 负向预测了“矛盾发生”的社交网络中的中心度; 尽责性显著正向预测了“支持给予”社交网络的中心度。以往研究表明, 宜人性的主要特征是同理心, 宽容和乐于助人(Goldberg, 1992; Graziano & Tobin, 2010), 高宜人性的个体被证明拥有更好的自我控制能力和情绪调节能力, 同时, 也更加擅长于在群体中调和矛盾(Graziano et al., 2007; Jensen-Campbell & Graziano, 2001)。另外对友谊选择原因的分析结果表明, “亲近朋友”社交网络中的友谊选择最看重值得信任和共享时光两个特征, 这也证明了高宜人性个体可能更能带给他人值得信赖和积极的感受, 从而在此社交网络中更受欢迎。除了宜人性以外, 当前研究结果表明尽责性高的个体在“支持给予”社交网络中更受欢迎, 并且这一社交网络中的友谊选择最看重值得信赖这一特质。Roberts等人(2005)认为, 尽责性是一个包含诚实、有社会责任感、以规则为导向、有力的自我控制等因素的整体特质, 而这些特质都会诱发一定程度上的人际吸引。并且, 大量研究都表明了尽责性高的个体具有更高的自我控制能力(Jensen- Campbell et al., 2002; Jensen-Campbell & Graziano, 2005), 而自我控制能力对于青少年做出符合社会规范的行为, 在更大的同辈群体中被接受, 以及与朋友建立牢固的社会关系都是至关重要的(Rothbart et al., 2000; Vohs & Baumeister, 2016; Yang et al., 2015)。因此, 高尽责性的个体可能是由于有着良好的自我控制能力从而让人感到更加值得信赖而在“支持给予”社交网络中更受欢迎。在大量的以往的社交网络相关研究中, 研究者们普遍都只在同一群体内建立单一的社交网络(Wolfer et al., 2012)。这些研究结果出现了一些分歧, 在部分研究中和社交网络有着显著相关关系的人格特质在另一些研究中则和社交网络无关(Landis, 2016)。然而, 在实际生活中, 同一个人往往同时存在于不同类型的关系定义的多个社交网络中, 个体可能会向一些人寻求建议, 向另一些人寻求陪伴, 再向另一些人寻求情感支持(Morelli et al., 2017; Wolfer et al., 2012)。当前研究的结果也支持了多个社交网络同时存在的假说, 证明了在同一群体内同时存在着不同的社交网络, 解决了以往人格特质和社交网络的相关研究中结果不一致的问题, 证明了一个人可以同时在一种类型的网络中占据中心地位, 但在另一些类型的网络中处于边缘位置。
大五人格相似性和社会距离的相关分析结果表明, 人格特质的差异性和“亲近朋友”和“消息分享”这两个社交网络中个体间的社会距离呈显著正相关关系, 即个体间人格差异越大, 社会距离就越远。友谊选择中人格的“同质性”假设已经在多项研究中被证明了。荷兰的一项研究发现同性青少年朋友间倾向于在外向性和宜人性中表现出相似性(Maarten et al., 2007)。除此之外, 相似的外向性和人际关系之间的联系也在配偶选择、婚姻压力和友谊选择的相关研究中被证明了(Furler et al., 2013; Gattis et al., 2004; Humbad et al., 2010; Maarten et al., 2007)。当前研究在整体人格特质的层面上证明了人格的相似性和个体间的人际吸引的相关关系。值得注意的是这种“同质性”吸引只存在于“亲近朋友”和“消息分享”两个社交网络中。友谊选择重要性问卷的分析结果表明, 这两个网络中的友谊选择都非常看重“相同的兴趣、爱好、价值观”和“共享时光”这两个特质。并且我们计算了不同网络在友谊选择因素上的相似性, 发现在“相同兴趣”这一友谊选择因素上这两个社交网络的相似性最强(= 0.54,< 0.001, 见网络版附录图1)。虽然仍不能完全排除其他造成这个两个社交网络同质性的因素, 但我们认为, “相同兴趣”这一友谊选择因素确实在“亲近朋友”和“消息分享”这两个社交网络的同质性中起着重要作用。早期观点认为个体间的相似性吸引主要来自两个方面, 一是个体间对环境的理解和反应的相似性增加了彼此在社会交往中的思想和行为的可预测性(Berger & Calabrese, 1975), 二是与人分享相似的价值观的同时会强化自己价值观和观点, 从而产生内隐的积极的情感反应, 这种积极的情感体验又会反过来促使人们在友谊选择时选择更加相似的人(Clore & Byrne, 1974)。无论是哪一种解释都强调了价值观和思想的相似性。因此, 整体人格的相似性可能通过个体间价值观的相似性对社会距离产生影响, 从而导致了在人际交往中人格特质越相似关系越亲密的现象。
通过选取“亲近朋友”和“消息分享”两个社交网络的社会距离与从大脑整体, 网络以及节点水平, 由大到小的三个尺度上的脑间相似性进行分析, 结果发现在整体大脑和网络水平上, 社会距离和脑间相似性都无显著相关, 只在节点水平上, 部分节点FC相似性和社交距离呈显著相关。大脑由多个功能网络组成, 不同的网络负责不同的功能系统, 但在更小的尺度, 节点, 甚至是体素水平上, 仍然存在着功能分化, 同一个节点与不同节点的功能连接都可能指向不同的认知功能(Power et al., 2011; Wig, 2017)。因此, 本研究的结果也在一定程度上说明, 大脑的FC并不是完全遵循越相似社交距离越近的原则, 而是在节点水平的FC相似性上显示出了两种不同的趋势:首先, 个体间静息态脑功能连接相似性与特定社交网络(“亲近朋友”和“消息分享”网络)的社会距离有着显著负相关关系, 这些FC分布在在额顶控制网络以及背侧注意网络等。额顶控制网络是脑网络研究中的热点之一, 包括部分外侧前额叶皮层和后顶叶皮层, 被认为和行动抑制, 情绪等过程有密切关系(Zanto & Gazzaley, 2013), 并且参与了多个高级认知任务, 在适应性认知控制中扮演了重要角色(Alahyane et al., 2014; Fassbender et al., 2006), 而这些认知过程都和个体的社会行为有着密切联系。最近一项讨论个体在观看自然影片时的大脑相似性和社会距离的关系的研究发现, 在与注意力分配、故事的解释和情感反应等过程相关的大脑区域(包括额顶网络)的神经反应强度的相似性和社交关系的亲密程度有着显著正相关关系(Parkinson et al., 2018)。因此, 个体间额顶控制网络内节点的FC相似性可能与个体间一般性认知过程的相似相对应, 进一步地, 由于个体间对环境的理解和反应的相似性会增加彼此在社会交往中的思想和行为的可预测性(Berger & Calabrese, 1975), 从而促进个体间产生人际吸引。背侧注意网络(dorsal attention network, DAN), 也叫视空间注意网络, 功能是提供自上而下的注意定向(Vogel et al., 2012)。在一项研究中, 研究者让被试观看不同情绪的影片, 分析个体间脑信号的相似性, 发现背侧注意网络对情绪的同步有重要作用, 并且研究者认为情绪的同步可以促进社会互动和人际理解(Nummenmaa et al., 2012)。另一项研究发现, 社交网络中关系密切的人在感知和解释周围的世界时, 注意力状态可能会随着时间的推移而发生异常相似的变化(Hyon et al., 2019)。因此, 本研究推断个体间背侧注意网络内FC的相似性可能和注意力和情绪的同步相对应, 而注意力和情绪的同步会促使社会交往中双方产生好感。
另外, “亲近的朋友”和“分享好消息”社交网络的社交距离还和部分节点FC相似性呈显著正相关, 其中大部分节点都属于默认网络, 包括右侧额上回、右侧扣带回等区域。这一结果表明这些节点的个体间FC模式越相似, 这两个社交网络中社交距离就越长, 个体间关系就越疏离。默认网络分布在大脑的内侧和外侧顶叶、内侧前额叶、内外侧颞叶皮质, 扣带回与楔前叶等区域(Greicius et al., 2009)。它通常被描述为一个统一的系统, 和情景记忆、心理理论、自我评价和反思等功能有关(Sestieri et al., 2011)。先前基于任务相关范式的研究表明, 当人们对自己的技能, 个性和未来做出评价时, 默认网络中的内侧前额叶, 前扣带皮层和内侧眶额皮层都会被激活(Goldin et al., 2014; Wu et al., 2015)。同时, 研究表明默认网络中的额上回, 颞中回以及眶额区域都为自我意识的产生和维持提供了支持(Fingelkurts & Fingelkurts, 2011)。自我意识被定义为一个人对自己精神或身体状态的内隐和外显的意识, 在社会生活中扮演着重要作用(Vogeley & Fink, 2003)。在一些更加强调人际关系的工具性价值的场合, 比如工作中, 群体的多样性是团队良性发展的重要因素之一(Ely & Thomas; McKinney, 2013; Schaffer, 2019)。并且研究者认为, 对于个体本身来说, 选择与自己不同的人作为合作伙伴更能体现自己的价值并便于在集体中找到自己的位置(Harrison et al., 1998)。本研究中的大学生群体即将步入社会, 逐渐更加注重人际关系的工具性价值(Schaffer, 2019)。这些证据表明在大学生的人际交往中, 个体在受到相似性的人际吸引的同时可能也会考虑到差异性带来的价值, 并且这可能和默认网络的自我意识功能有关。
综上所述, 本研究在同一群体内测量了多个社交网络, 为社交网络相关研究结果出现的分歧提供了一种解决思路, 有助于我们更加精确和客观地理解社交网络的结构及其影响因素。并且, 本研究综合考虑了人格特质对社交网络中节点和连边的影响, 有助于我们全方位的理解人格特质对社交网络的重要影响, 对我们理解社交网络的结构, 形成规律以及其中的信息传播规律有着重要意义。此外, 本研究将静息态大脑功能连接相似性纳入了社会距离的分析, 为人际交往中个体间的大脑同步性研究提供了新的证据, 同时, 也在一定程度上为人格特质相似性和社会距离关系的研究提供了脑影像层面的证据。但是, 本研究仍存在一些不足和可以继续挖掘的地方。本研究采集社交网络数据的时间是在被试大一下学期进行的, 这个时间点他们的人际网络已经逐渐稳定, 这些数据只能反映他们目前稳定的人际关系, 但不能进一步反映和讨论他们在刚刚接触阶段的友谊选择状况, 可能会错过部分友谊选择的关键信息。因此, 未来的研究也许可以针对社交网络开展纵向研究, 阶段性考察社交网络的形成过程, 并将更多的影响因素考虑进来。其次, 本研究在对不同网络进行划分时只参考了样本二的描述性结果, 但样本二中囊括的影响友谊选择的因素是有限的并且描述性统计的结果能反应数据形态的程度也是有限的, 因此可能不足以客观全面的作为划分社交网络的依据。在以后的研究中也许能通过图论等方法或开创性的实验设计对社交网络进行更加客观全面的分类。
5 结论
本研究采用网络科学方法, 探讨了人格特质对个体在社交网络中的中心地位和个体间的社会距离的影响以及个体间大脑静息态功能连接相似性对社会距离的影响, 具体发现:
第一, 在同一群体中, 根据社交需求存在不同的社交网络, 并且个体在不同社交网络中的地位也不尽相同, 会受到相应人格特质的影响。具体而言, 高尽责性的个体在需要“值得信任”特质的社交网络中更受欢迎, 高宜人性的个体在需要 “共享时光”的社交网络中更受欢迎。
第二, 在需要“相同兴趣”特质的社交网络中, 人格的相似性对个体间的人际交往有促进作用, 这种促进作用可能是通过价值观和兴趣的相似性诱发的人际吸引来实现的。
第三, 个体间静息态脑功能连接相似性可能对特定社交网络(“亲近朋友”和“消息分享”)中的人际交往有促进作用, 这些FC分布在额顶控制网络以及背侧注意网络, 可能反应了个体在认知控制以及对环境的加工偏向上趋于相似, 从而增加了彼此的人际吸引, 缩短了相互之间的社会距离。同时, 与社交距离呈显著正相关的节点FC相似性主要分布在默认网络, 表明在大学生的人际交往中, 个体在受到相似性的人际吸引的同时可能也会考虑到差异性带来的价值, 并且这可能和默认网络的自我意识功能有关。
社交网络问卷(Morelli et al., 2018; Morelli et al., 2017):
(1)“谁是你很亲近朋友?”;
(2)“你经常和谁待在一起(课余时间)进行一些社交活动, 比如吃饭, 看电影, 逛街等等?”;
(3)“你经常和谁寻求社交生活上(工作学习除外)的建议?”;
(4)“当有不好的事儿发生的时候, 你会向谁求助?”;
(5)“你会和谁消息分享?”;
(6)“你经常感受到谁的支持和关心?”;
(7)“你觉得谁是非常善解人意?”;
(8)“你觉得谁通常会让你感觉很积极(例如, 快乐、热情)?”;
(9)“你和谁矛盾发生?”。
附表1 和“亲近朋友”社交网络中的社会距离呈显著相关的节点信息

NO.脑区半球脑网络MNI坐标rp xyz 22Precuneus_RRSensory/somatomotor Hand8–48690.12 0.04 97Frontal_Sup_RRDefault mode2127500.15 0.03 101Frontal_Mid_RRDefault mode2033420.13 0.06 110Cingulum_Ant_RRDefault mode73700.19 0.00 124Lingual_LLDefault mode–25–41–80.17 0.00 126Fusiform_LLDefault mode–32–39–150.16 0.00 194Occipital_Mid_RRFronto–parietal Task Control35–66380.12 0.02 193Frontal_Mid_RRFronto–parietal Task Control29957–0.16 0.03 204Angular_RRSalience52–4736–0.13 0.01
附表2 和“日常活动”社交网络中的社会距离呈显著相关的节点信息
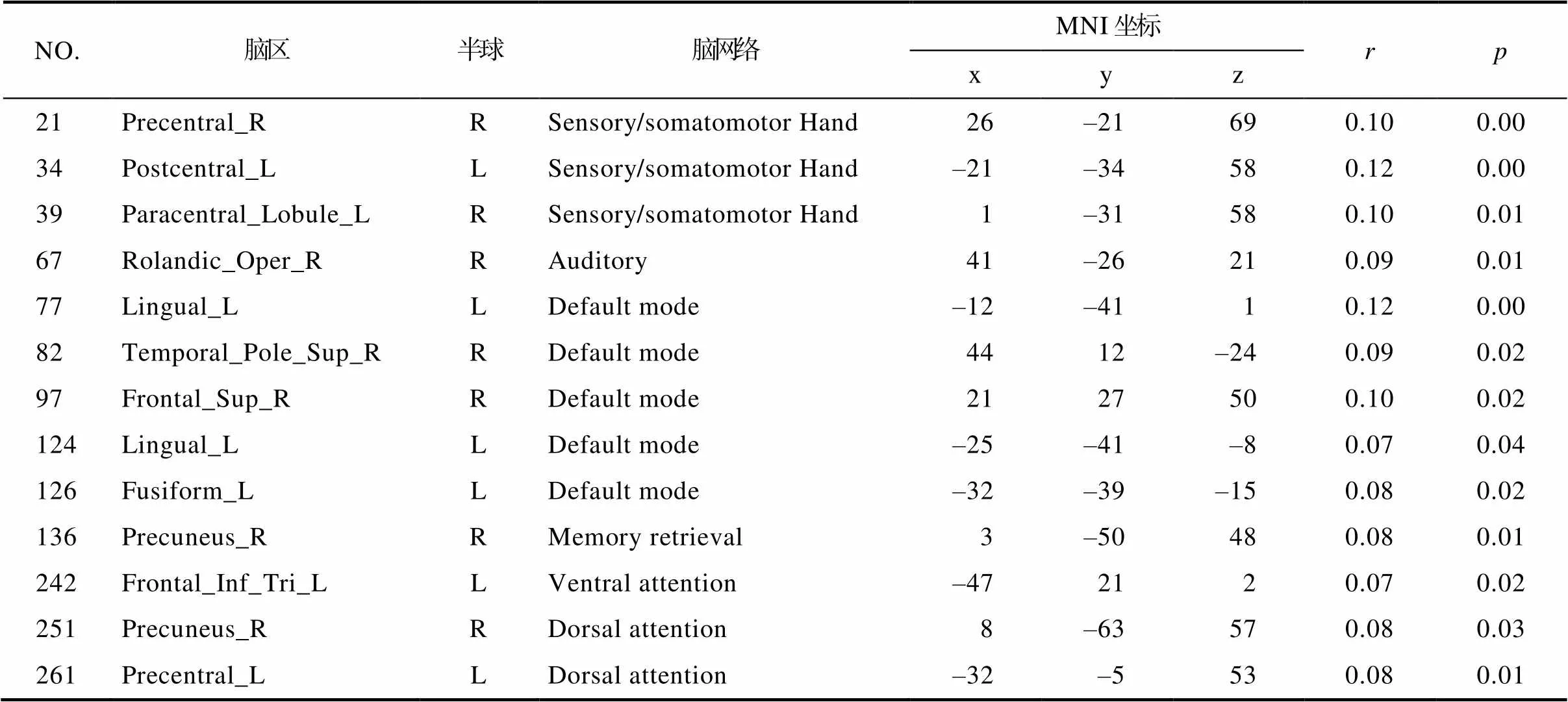
NO.脑区半球脑网络MNI坐标rp xyz 21Precentral_RRSensory/somatomotor Hand26–21690.100.00 34Postcentral_LLSensory/somatomotor Hand–21–34580.120.00 39Paracentral_Lobule_LRSensory/somatomotor Hand1–31580.100.01 67Rolandic_Oper_RRAuditory41–26210.090.01 77Lingual_LLDefault mode–12–4110.120.00 82Temporal_Pole_Sup_RRDefault mode4412–240.090.02 97Frontal_Sup_RRDefault mode2127500.100.02 124Lingual_LLDefault mode–25–41–80.070.04 126Fusiform_LLDefault mode–32–39–150.080.02 136Precuneus_RRMemory retrieval3–50480.080.01 242Frontal_Inf_Tri_LLVentral attention–472120.070.02 251Precuneus_RRDorsal attention8–63570.080.03 261Precentral_LLDorsal attention–32–5530.080.01
附表3 和“社交建议”社交网络中的社会距离呈显著相关的节点信息

NO.脑区半球脑网络MNI坐标rp xyz 82Temporal_Pole_Sup_RRDefault mode4412–240.06 0.01 117Temporal_Mid_LLDefault mode–53–15–90.06 0.00 248Fusiform_LLUncertain–29–12–330.04 0.01 8Fusiform_LLUncertain–35–30–24–0.04 0.03 79Temporal_Mid_LLDefault mode–44–6118–0.05 0.02
附表4 和“寻求帮助”社交网络中的社会距离呈显著相关的节点信息

NO.脑区半球脑网络MNI坐标rp xyz 242Frontal_Inf_Tri_LLVentral attention–472120.07 0.01 33Parietal_Inf_LLSensory/somatomotor –44–3444–0.09 0.00 37Precentral_LLSensory/somatomotor –38–1866–0.06 0.03 159Cuneus_RRVisual14–7728–0.07 0.02 220Frontal_Mid_LLSalience–384521–0.07 0.01 228PutamenLSubcortical–15010–0.05 0.03
附表5 和“消息分享”社交网络中的社会距离呈显著相关的节点信息
NO.脑区半球脑网络MNI坐标rp xyz 7ParaHippocampal_RRUncertain17–30–150.10 0.03 82Temporal_Pole_Sup_RRDefault mode4412–240.16 0.00 97Frontal_Sup_RRDefault mode2127500.13 0.02 101Frontal_Mid_RRDefault mode2033420.17 0.01 126Fusiform_LLDefault mode–32–39–150.12 0.01 205Precentral_RRSalience39–5480.11 0.05 210Insula_RRSalience352730.09 0.02 237Temporal_Mid_LLVentral attention–53–41120.13 0.02 241Frontal_Inf_Tri_RRVentral attention502760.08 0.02 70Temporal_Sup_LLAuditory–53–1212–0.12 0.03 201Frontal_Inf_Tri_LLFronto–parietal Task Control–412031–0.11 0.02 252Temporal_Mid_LLDorsal attention–50–633–0.17 0.00
附表6 和“支持给予”社交网络中的社会距离呈显著相关的节点信息

NO.脑区半球脑网络MNI坐标rp xyz 13Precuneus_LLSensory/somatomotor Hand–8–54570.08 0.01 82Temporal_Pole_Sup_RRDefault mode4412–240.09 0.01 97Frontal_Sup_RRDefault mode2127500.08 0.02 76Frontal_Med_Orb_RRDefault mode842–9–0.08 0.02
附表7 和“善解人意”社交网络中的社会距离呈显著相关的节点信息

NO.脑区半球脑网络MNI坐标rp xyz 23Postcentral_LLSensory/somatomotor Hand–23–33690.14 0.01 56Insula_RRCingulo–opercular Task Control47430.15 0.02 61Heschl_RRAuditory30–29140.14 0.01 63Temporal_Sup_RRAuditory55–19100.15 0.01 64Rolandic_Oper_LLAuditory–37–35160.12 0.02 65Temporal_Sup_LLAuditory–58–27130.16 0.01 66Temporal_Sup_LLAuditory–47–2850.12 0.02 72SupraMarginal_RRAuditory56–21300.16 0.01 99Frontal_Sup_LLDefault mode–1723540.13 0.02 104Frontal_Sup_LLDefault mode–2039420.13 0.02 105Cingulum_Ant_RRDefault mode548210.14 0.02 111Cingulum_Ant_LLDefault mode–1139120.13 0.03 117Temporal_Mid_LLDefault mode–53–15–90.13 0.02 119Temporal_Mid_RRDefault mode62–33–60.14 0.02 139Frontal_Inf_Orb_RRDefault mode4730–60.14 0.01 140Lingual_RRUncertain8–90–90.09 0.04 143Lingual_RRVisual17–48–90.21 0.00 145Calcarine_RRVisual8–7290.13 0.02 150Fusiform_RRVisual26–60–90.17 0.00 151Lingual_LLVisual–14–72–90.14 0.02 159Cuneus_RRVisual14–77280.14 0.01 167Cuneus_LLVisual–3–81180.14 0.02 177Parietal_Inf_LLFronto–parietal Task Control–51–50390.11 0.01 184Cerebelum_Crus2_RRUncertain17–79–340.13 0.02 227Pallidum_LLSubcortical–214–20.14 0.01 229Putamen_RRSubcortical29–1740.14 0.02 231Putamen_RRSubcortical27–370.16 0.01 242Frontal_Inf_Tri_LLVentral attention–472120.12 0.01 1Lingual_LLUncertain–23–96–15–0.14 0.00 201Frontal_Inf_Tri_LLFronto–parietal Task Control–412031–0.11 0.02
附表8 和“积极感受”社交网络中的社会距离呈显著相关的节点信息

NO.脑区半球脑网络MNI坐标rp xyz 20Parietal_Inf_LLSensory/somatomotor Hand–52–25410.11 0.02 35Paracentral_Lobule_LLSensory/somatomotor Hand–14–21720.12 0.00 176Frontal_Inf_Oper_LLFronto–parietal Task Control–457240.10 0.01 187Precentral_LLFronto–parietal Task Control–402330.11 0.01 194Occipital_Mid_RRFronto–parietal Task Control35–66380.09 0.01 222Thalamus_RRSubcortical6–2610.08 0.01 13Precuneus_LLSensory/somatomotor –8–5457–0.10 0.02 76Frontal_Med_Orb_RRDefault mode842–9–0.11 0.02 217Cingulum_Ant_RRSalience91730–0.12 0.01
为了探究友谊选择考虑因素对网络间相似性的影响, 我们计算了不同网络在友谊选择因素上的相似性, 发现在所有友谊选择因素上, 所有网络也都呈显著相关, 也就是说虽然个体在不同社交网络中进行友谊选择时考虑因素的排序会有所不同, 但在任意社交网络中, 个体对同一个友谊选择因素的看重程度是相似且相对稳定的。

附图1 不同社交网络在友谊选择因素上的相关矩阵图:第一个矩阵是原始社交网络间的相关矩阵图, 剩下的是根据友谊选择因素计算出来的网络间相似性矩阵, 所有矩阵从上到下从左到右的网络分别是:亲近朋友, 日常活动, 社交建议, 寻求帮助, 消息分享, 支持给予, 善解人意和积极感受网络。
Alahyane, N., Brien, D. C., Coe, B. C., Stroman, P. W., & Munoz, D. P. (2014). Developmental improvements in voluntary control of behavior: Effect of preparation in the fronto-parietal network?, 103–117.
Berger, C. R., & Calabrese, R. J. (1975). Some explorations in initial interaction and beyond: Toward a developmental theory of interpersonal communication., 99–112.
Blakemore, S. (2008). The social brain in adolescence.(4), 267–277.
Borgatti, S. P., Mehra, A., Brass, D. J., & Labianca, G. (2009). Network analysis in the social sciences., 892–896.
Clore, G. L., & Byrne, D. (1974). A reinforcement-affect model of attraction., 143– 170.
Costa, P. T., & McCrae, R. R. (1992). Normal personality assessment in clinical practice: The NEO personality inventory.(1), 5.
Ely, R. J., & Thomas, D. A. Cultural diversity at work: The effects of diversity perspectives on work group processes and outcomes., 229– 273.
Falk, E. B., & Bassett, D. S. (2017). Brain and social networks: Fundamental building blocks of human experience.(9), 674–690. doi:10.1016/j.tics. 2017.06.009
Fassbender, C., Simoes-Franklin, C., Murphy, K., Hester, R., Meaney, J., Robertson, I. H., & Garavan, H. (2006). The role of a right fronto-parietal network in cognitive control.(4), 286–296. doi:10.1027/ 0269-8803.20.4.286
Feiler, D. C., & Kleinbaum, A. M. (2015). Popularity, similarity, and the network extraversion bias.(5), 593–603. doi:10.1177/0956797615569580
Fingelkurts, A. A., & Fingelkurts, A. A. (2011). Persistent operational synchrony within brain default-mode network and self-processing operations in healthy subjects.(2), 79–90. doi:10.1016/j.bandc.2010.11. 015
Furler, K., Gomez, V., & Grob, A. (2013). Personality similarity and life satisfaction in couples.(4), 369–375. doi:10.1016/j. jrp.2013.03.002
Gattis, K. S., Berns, S., Simpson, L. E., & Christensen, A. (2004). Birds of a feather or strange birds? Ties among personality dimensions, similarity, and marital quality.(4), 564–574. doi:10. 1037/0893-3200.18.4.564
Goldberg, L. R. (1992). The development of markers for the Big-Five factor structure..
Goldin, P. R., Ziv, M., Jazaieri, H., Weeks, J., Heimberg, R. G., & Gross, J. J. (2014). Impact of cognitive-behavioral therapy for social anxiety disorder on the neural bases of emotional reactivity to and regulation of social evaluation., 97–106. doi:10. 1016/j.brat.2014.08.005
Graziano, W. G., Habashi, M. M., Sheese, B. E., & Tobin, R. M. (2007). Agreeableness, empathy, and helping: A person x situation perspective.(4), 583–599. doi:10.1037/0022-3514.93. 4.583
Graziano, W. G., & Tobin, R. M. (2010). Agreeableness: Dimension of personality or social desirability artifact?.(5), 695–728.
Greicius, M. D., Supekar, K., Menon, V., & Dougherty, R. F. (2009). Resting-state functional connectivity reflects structural connectivity in the default mode network.(1), 72–78. doi:10.1093/cercor/bhn059
Harrison, D. A., Price, K. H., & Bell, M. P. (1998). Beyond relational demography: Time and the effects of surface-and deep-level diversity on work group cohesion.(1), 96–107.
Humbad, M. N., Donnellan, M. B., Iacono, W. G., McGue, M., & Burt, S. A. (2010). Is spousal similarity for personality a matter of convergence or selection?(7), 827–830. doi:10.1016/j.paid. 2010.07.010
Hyon, R., Kleinbaum, A. M., & Parkinson, C. (2019). Social network proximity predicts similar trajectories of psychological states: Evidence from multi-voxel spatiotemporal dynamics., 116492. doi:10. 1016/j.neuroimage.2019.116492
Jaaskelainen, I. P., Pajula, J., Tohka, J., Lee, H. J., Kuo, W. J., & Lin, F. H. (2016). Brain hemodynamic activity during viewing and re-viewing of comedy movies explained by experienced humor., 27741. doi:10. 1038/srep27741
Jensen-Campbell, L. A., Adams, R., Perry, D. G., Workman, K. A., Furdella, J. Q., & Egan, S. K. (2002). Agreeableness, extraversion, and peer relations in early adolescence: Winning friends and deflecting aggression.(3), 224–251. doi:10.1006/jrpe.2002.2348
Jensen-Campbell, L. A., & Graziano, W. G. (2001). Agreeableness as a moderator of interpersonal conflict..
Jensen-Campbell, L. A., & Graziano, W. G. (2005). The two faces of temptation: Differing motives for self-control., 287–314.
Kaplan, S. C., Levinson, C. A., Rodebaugh, T. L., Menatti, A., & Weeks, J. W. (2015). Social anxiety and the big five personality traits: The interactive relationship of trust and openness.(3), 212–222. doi:10.1080/16506073.2015.1008032
Klein, K. J., Lim, B.-C., Saltz, J. L., & Mayer, D. M. (2004). How do they get there? An examination of the antecedents of centrality in team networks.(6), 952–963. doi:10.2307/20159634
Landis, B. (2016). Personality and social networks in organizations: A review and future directions., S107–S121. doi:10.1002/job. 2004
Maarten, S., Susan, B., & Wim, M. (2007). Similarity in adolescent best friendships: The role of gender.(2), 42–48. doi:10.1007/bf03061061
Massen, J. J. M., & Koski, S. E. (2014). Chimps of a feather sit together: Chimpanzee friendships are based on homophily in personality.(1), 1–8. doi:10.1016/j.evolhumbehav.2013.08.008
McKinney, A. P. (2013). What lies ahead: Emerging trends in the diversity pipeline among new entrants in the workforce.(1/2), 83–95.
McPherson, M., Smith-Lovin, L., & Cook, J. M. (2001). Birds of a feather: Homophily in social networks., 415–444.
Morelli, S. A., Leong, Y. C., Carlson, R. W., Kullar, M., & Zaki, J. (2018). Neural detection of socially valued community members.(32), 8149–8154.
Morelli, S. A., Ong, D. C., Makati, R., Jackson, M. O., & Zaki, J. (2017). Empathy and well-being correlate with centrality in different social networks.(37), 9843–9847. doi:10.1073/ pnas.1702155114
Nummenmaa, L., Glerean, E., Viinikainen, M., Jaaskelainen, I. P., Hari, R., & Sams, M. (2012). Emotions promote social interaction by synchronizing brain activity across individuals.(24), 9599–9604. doi:10.1073/pnas.1206095109
Parkinson, C., Kleinbaum, A. M., & Wheatley, T. (2018). Similarneural responses predict friendship.(1), 332. doi:10.1038/s41467-017-02722-7
Power, J. D., Cohen, A. L., Nelson, S. M., Wig, G. S., Barnes, K. A., Church, J. A., … Petersen, S. E. (2011). Functional network organization of the human brain.(4), 665–678. doi:10.1016/j.neuron.2011.09.006
Roberts, B. W., Chernyshenko, O. S., Stark, S., & Goldberg, L. R. (2005). The structure of conscientiousness: An empirical investigation based on seven major personality questionnaires., 103–139.
Rothbart, M. K., Ahadi, S. A., & Evans, D. E. (2000). Temperament and personality: Origins and outcomes.(1), 122.
Schaffer, B. S. (2019). Examining reactions to workplace diversity: The role of dissimilarity-attraction in teams.(1), 57–69. doi:10.1002/cjas.1476
Selfhout, M., Burk, W., Branje, S., Denissen, J., van Aken, M., & Meeus, W. (2010). Emerging late adolescent friendship networks and big five personality traits: A social network approach.(2), 509–538. doi:10.1111/j.1467-6494.2010.00625.x
Sestieri, C., Corbetta, M., Romani, G. L., & Shulman, G. L. (2011). Episodic memory retrieval, parietal cortex, and the default mode network: Functional and topographic analyses.(12), 4407–4420. doi:10.1523/JNEUROSCI.3335-10.2011
Simon, S. S., Varangis, E., & Stern, Y. (2020). Associations between personality and whole-brain functional connectivity at rest: Evidence across the adult lifespan.(2), e01515. doi:10.1002/brb3.1515
Vogel, A. C., Miezin, F. M., Petersen, S. E., & Schlaggar, B. L. (2012). The putative visual word form area is functionally connected to the dorsal attention network.(3), 537–549. doi:10.1093/cercor/bhr100
Vogeley, K., & Fink, G. R. (2003). Neural correlates of the first-person-perspective.(1), 38–42.
Vohs, K. D., & Baumeister, R. F. (2016).. Guilford Publications.
Wig, G. S. (2017). Segregated systems of human brain networks.(12), 981–996. doi:10.1016/j.tics.2017.09.006
Wolfer, R., Cortina, K. S., & Baumert, J. (2012). Embeddedness and empathy: How the social network shapes adolescents' social understanding.(5), 1295–1305. doi:10.1016/j.adolescence. 2012.04.015
Wu, J., Dong, D., Jackson, T., Wang, Y., Huang, J., & Chen, H. (2015). The neural correlates of optimistic and depressive tendencies of self-evaluations and resting-state default mode network., 618. doi:10.3389/fnhum.2015.00618
Wu, X. R., He, H., Shi, L., Xia, Y. M., Zuang, K. X., Feng, Q. Y., … Qiu, J. (2019). Personality traits are related with dynamic functional connectivity in major depression disorder: A resting-state analysis., 1032–1042. doi:10.1016/j.jad.2018.11.002
Yang, W. J., Cun, L. L., Du, X., Yang, J. Y., Wang, Y. Q., Wei, D. T., … Qiu, J. (2015). Gender differences in brain structure and resting-state functional connectivity related to narcissistic personality.(1). doi:10. 1038/srep10924
Zanto, T. P., & Gazzaley, A. (2013). Fronto-parietal network: flexible hub of cognitive control.(12), 602–603. doi:10.1016/j.tics.2013.10.001
The influence of personality traits and brain functional connectivity on social networks
LI Yiman, LIU Cheng, ZHUANG Kaixiang, HUO Tengbin, XU Pengfei, LUO Yuejia, QIU Jiang
(School of Psychology, Southwest University;Key Laboratory of Cognition and Personality, Chongqing 400715, China) (School of Psychology, Shenzhen University;Center for Brain Disorders and Cognitive Sciences, Shenzhen 518060, China)(Faculty of Psychology, Beijing Normal University, Beijing 100875, China) (Teacher Education College, Qilu Normal University;Institute of Psychology and Cognitive Neuroscience, Jinan 250200, China)
Humans are a social species that are constantly involved in complex relationships, reacting to the actions of others, and intentionally or unintentionally changing our own behavior. Personality traits reflect the behavioral pattern of an individual's response to the environment, which also includes social behavior. In addition, the brain is also an important factor when discussing social networks. The brain provides biological mechanisms for human behavior, while social networks provide external triggers for these behaviors. Linking personality traits and brain activity to social networks can help us better understand the structure of group relationships, improve our understanding of individual human beings, and help us better predict individual social behaviors and find the rules of information transmission in interpersonal relationships.From the perspective of a network, we collected nine social networks from 94 undergraduate students in the same grade according to their different social needs. We used the graph theory and resting-state functional magnetic resonance imaging to explore the influence of personality traits on social networks based on individual popularity and closeness between individuals and the relationship between the similarity of brain resting-state functional connectivity and social distance between individuals. Specifically, regression analysis was carried out, with personality traits as the independent variables and the different degrees of social networks as the dependent variables. Then, a correlation analysis was performed for the social distance and similarity of personality traits. Finally, the correlation between the similarity of the brain networks and social distance was calculated. The results showed that (1) individuals with high conscientiousness were more popular in social networks requiring "trust" traits, while individuals with high agreeableness were more popular in social networks requiring "fun" traits. These findings showed that in the same group, there are different social networks according to social needs, and the popularity of individuals in different social networks is not similar as it will be affected by the corresponding personality traits; (2) Inthe social networks requiring "shared interests & values, " personality similarity and social distance between individuals were significantly negatively correlated. Personality similarity promotes interpersonal communication between individuals, which may be realized through interpersonal attraction induced by the similarity of values and interests; (3) In the same social network, there is a significantly negative correlation between similarities in functional connections (FCs) and social distance among individuals, and these FCs are mainly concentrated in the fronto-parietal task control network and the dorsal attention network. The similarity of resting brain FCs among individuals may promote interpersonal communication, possibly due to the similarity of individuals in cognitive control and environmental processing bias, which increases the interpersonal attraction and shortening the social distance between individuals.The results revealed the influence of personality traits on the structure of different social networks, the relationship between personality trait similarity among individuals, and the similarity between resting brain networks and social distance, which has important implications for understanding the structure of social networks, the formation rules, and the information transmission rules among them. In addition, this study discussed the relationship between the similarity of resting-state FC and social distance, providing new evidence for studies on brain synchronization in interpersonal communication and brain imaging evidence for the study of the relationship between the similarity of personality traits and social distance.
social networks, personality traits, resting-state functional connectivity
2021-01-11
国家自然科学基金项目(31771231), 重庆市自然科学基金项目(cstc2019jcyj-msxmX0520), 重庆市社会科学规划项目(2018PY80), 中央高校基本科研业务专项资金(SWU119007), 国家社科重大项目(20&ZD153),广东省科技项目(2019A050510048)。
李依蔓和刘程为共同第一作者。
邱江, E-mail: qiuj318@swu.edu.cn
B849: C91; B845
