独尾草属2个同域种的种群结构与动态特征
2021-12-06刘丹吴玲安静蒋超亮王绍明
刘丹,吴玲,安静,蒋超亮,王绍明
(石河子大学生命科学学院,新疆 石河子 832003)
独尾草属植物在我国分布有4种,有3种主要分布于新疆,分别为阿尔泰独尾草(E.altaicus)、异翅独尾草(E.anisopterus)和粗柄独尾草(E.inderiensis),其中,后两者于古尔班通古特沙漠南缘同域分布。作为类短命植物,异翅独尾草与粗柄独尾草在沙漠固定、保持水土以及植被恢复方面等具有重要意义。
异翅独尾草和粗柄独尾草都有总状花序,种子三菱形,具翅的特点,区别在于前者茎上无毛,花被片有一条暗褐色脉,而后者茎密被短柔毛,花被片上有三条深褐色脉。两者生长在古尔班通古特沙漠中的固定、半固定沙丘上,是古尔班通古特沙漠春季的主要植被景观[1-5]。
生境破碎化是当下的研究热点。生境破碎化会导致物种多样性以及生态功能的变化,从而影响植物种群的大小、灭绝速度等,影响植物的种群动态。自上世纪末起,在古尔班通古特沙漠周边地区,人类活动不断的加剧,致使沙漠南缘的各种群呈现出相互分离的状态[1-3,9-10]。
在不同生境破碎化程度下,独尾草属植物作为一种类短命植物,不同部位的种子大小及萌发特性不同,具备主动适应逆境的独特策略[2,4,9]。
种群动态是种群生态学研究的重点,研究内容主要包括种群年龄结构、静态生命表以及存活曲线。种群动态是指生活在特定环境中的生物种群,随着时间的改变而呈现出个体数目的消长以及分布变迁的现象。研究种群数量在时空上的变化规律不仅有助于分析种群的内部结构,而且有利于了解种群的发展方向[6-9,11]。
本文对古尔班通古特沙漠南缘中的不同生境下异翅独尾草与粗柄独尾草种群进行样地调查,分析了尾草属2个同域种的种群结构、静态生命表、活曲线、死亡曲线,发现生境破碎化已经对独尾草属2个同域种的种群结构产生了负面影响,这一结果对深刻了解其种群的现状并对其种群的未来进行预测具有重要意义[17-19]。
1 材料与方法
1.1 研究区概况
研究区位于古尔班通古特沙漠南缘,海拔320米左右。通过野外调查发现,研究区内春季的主要植物种有:异翅独尾草、粗柄独尾草、沙大戟(Chrozophorasabulosa)、早熟猪毛菜(Salsolapraecox)、东方旱麦草(Eremopyrumorientale)、梭梭(Haloxylonammodendron)、白梭梭(HaloxylonPersicum)等[10]。
1.2 样地的选择
经过踏查,并考虑研究物种的生长状况,在沙漠南缘选取10个样地,分别编号为D1、D2、D3…D10,并且记录各样地的基本信息(表1)。
1.3 样地类型的划分
样地类型的划分详细情况见表1,图1。
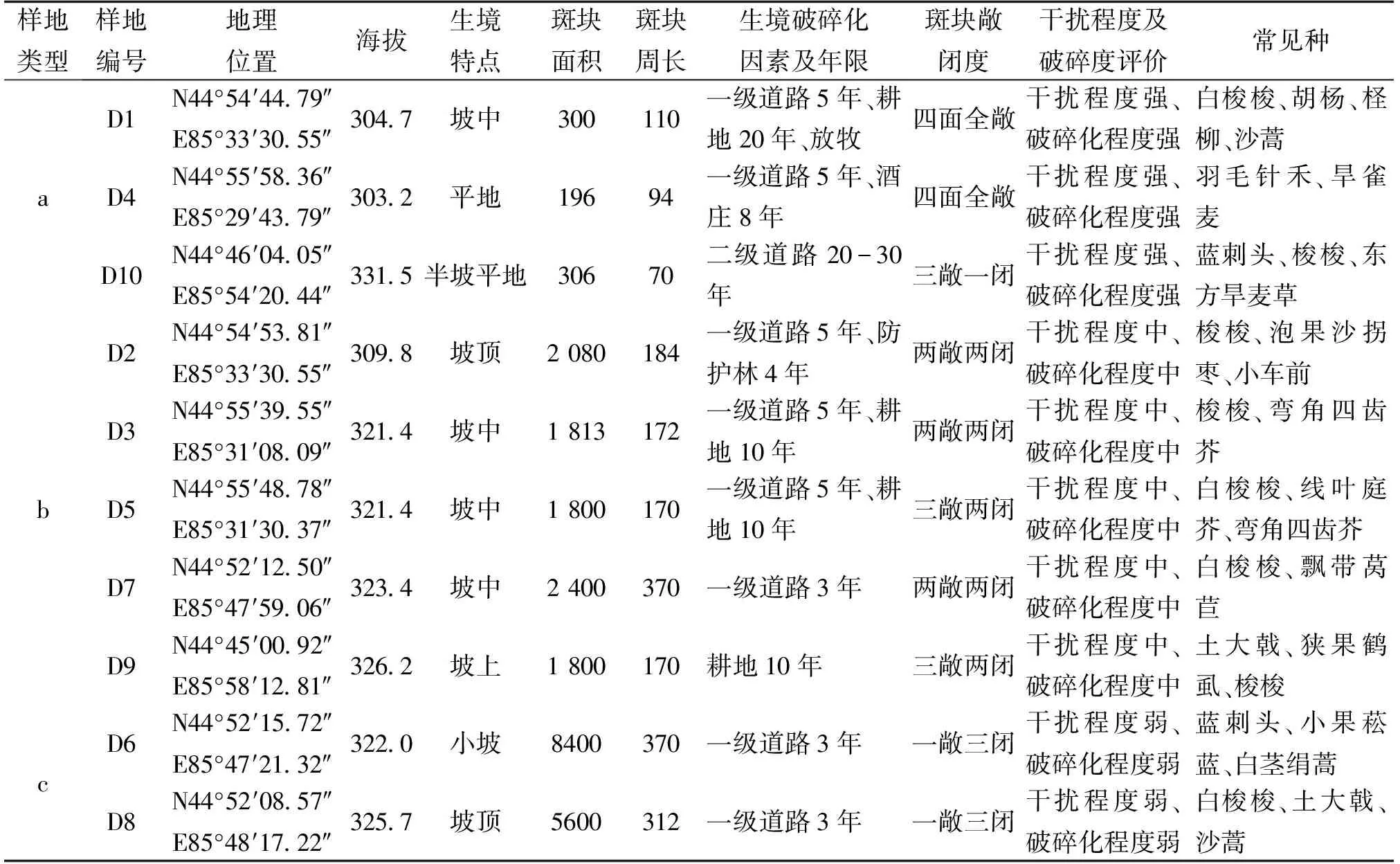
表1 各样地基本信息
根据调查结果,样地信息表中的9个环境因子使用SPSS分析软件的欧式距离离差平方和法做树状聚类分析,可将10个样地分为3个类型,分别记为a类型、b类型、c类型(图1,表1)。

图1 欧式距离离差平法和法分层聚类谱系树状图
根据样地的分类结果,a类型为高生境破碎程度的样地组合,由3个斑块构成:D1、D4、D10;斑块敞闭度为近全敞开型;b类型由中等破碎程度的斑块组合而成,组成的斑块有 D2、D3、D5、D7、D9;斑块敞闭度大多为两敞两闭型;c类型为低破碎程度的斑块构成,包括 D6、D8两个斑块;斑块敞闭度近全闭合型。
1.4 研究方法
1.4.1 调查方法
采用样方法对3种类型样地中的2个独尾草属植物进行研究。在每个样地内分别选取3个10 m×10 m样方,调查各样方内的2种独尾草株数以及轮叶数(即叶片数,每3片轮叶划分为一个龄级)。
1.4.2 龄级划分
不同的物种其龄级的确定可以采取不同的方法,对于独尾草属植物来讲,不宜使用径级大小作为划分依据。根据野外调查的实际情况以及相关文献[15],本研究采取以轮叶数代替年龄的方法,绘制独尾草属2个同域种的种群年龄结构图。
1.4.3 静态生命表
由于独尾草属植物属于百合科多年生的类短命植物,因此采取“空间代替时间”的方法编制种群的静态生命表,进而分析其动态变化[16]。根据静态生命表的编制要求,以调查得出的3种类型下两种独尾草在每个样地中每一龄级的平均存活数为基础,绘制2016年异翅独尾草的静态生命表(表2)与粗柄独尾草的静态生命表(表3)。
1.4.4 存活曲线和死亡曲线
以龄级作为横坐标,以计算得到的标准化存活数和死亡率为纵坐标,绘制得到该种群的存活曲线和死亡曲线。
2 结果与分析
2.1 年龄结构分析
从图2中可以看出:a类型样地中,异翅独尾草植株数量较b类型和c类型样地的少,其中异翅独尾草平均仅有6株,出现某一或多个龄级植株缺失的情况,尤其是高龄级的植株严重缺失,但是在其他龄级均有一定数目的分布。而粗柄独尾草则在每个龄级都有分布,其中低龄级植株(即1~3龄级)的数量相对多,高龄级植株(大于11龄级)的数量少;b类型样地中,除了异翅独尾草在2个高龄级(大于11龄级)有龄级缺失的情况外,其他龄级均有一定数目的植株个体分布,其中第1、2、3龄级的植株占植株总数的 39.82%,幼苗个体数占据绝对的优势,而粗柄独尾草则在每个龄级均有分布,其中,第1、2、3龄级的植株占植株总数的46.03%;c类型样地中的两种独尾草属植物种群的个体数均高于a类型和b类型样地,且两种独尾草属植物在3个较高龄级中均出现了植株数量少甚至缺失的情况。

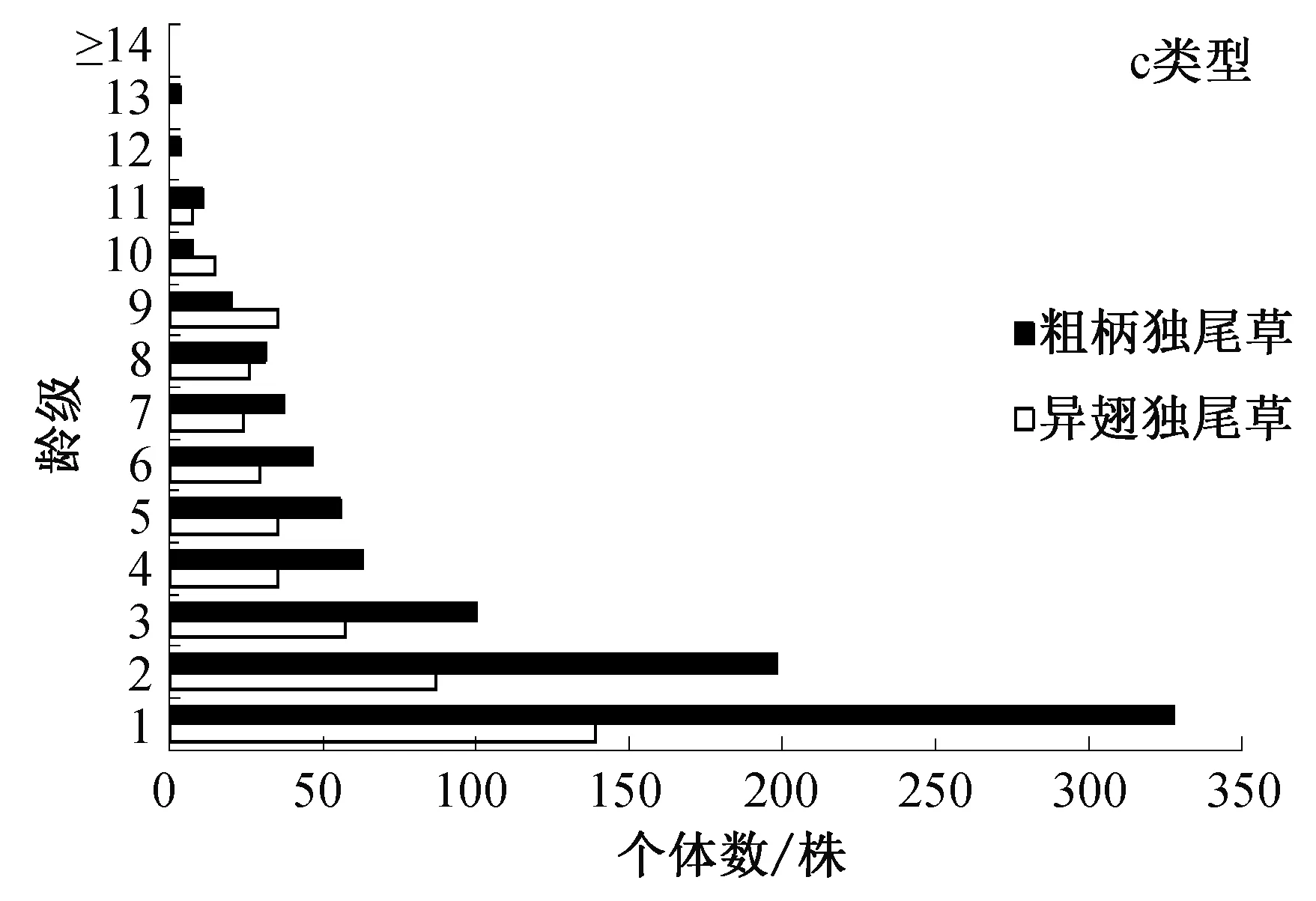
图2 3种样地类型中两种独尾草属植物的种群年龄结构
总体而言,a类型样地中,粗柄独尾草种群在各个龄级均有分布,而异翅独尾草出现高龄级缺失的情况,两种独尾草的种群数量也较另两种类型少,且幼苗数量相对少,不能维持种群稳定,属于衰退型种群;b类型的样地受到中等程度的干扰,虽然异翅独尾草的种群因幼苗数目较大而表现为稳定型,但其幼苗死亡率较高,因此将其归属于稳定至衰退型种群。而粗柄独尾草种群幼苗数量丰富,种群规模大,维持有足够的后续资源,更新潜力较强,处在一种良性的状态,属于稳定型;c类型样地中,异翅独尾草种群受到较轻的人为干扰,种群规模大且幼苗数目丰富,种群更新潜力较强,处在一种增长状态。粗柄独尾草幼苗个体数目占据绝对优势,年龄结构呈现金字塔形,属于典型的增长种群,粗柄独尾草可以较好的适应较轻的生境破碎化程度,能够长期稳定的自我发展。
2.2 种群静态生命表
根据静态生命表的编制要求,编制了2种独尾草属植物种群的静态生命表(表2、表3)。由表2可以看出,对于异翅独尾草而言,a类型样地的第2、4、5、11龄级、b类型样地的第3龄级以及c类型样地的第7、8龄级,都是低龄级植株的存活数小于比其高一龄级植株的存活数,使得生命表中相应龄级的dx、qx、Kx表现为负值,反映了异翅独尾草种群在该龄级段里幼株的缺乏程度。同样的情况也存在于粗柄独尾草种群中,通过表3我们发现,a类型样地中的第1、4、13龄级、b类型样地中的第1、4、7、11龄级的存活数也小于比其高一龄级植株的存活数。相反,在c类型样地中,粗柄独尾草种群幼苗数量较大,在每一个龄级都有分布,只在第10龄级出现了负值的情况。
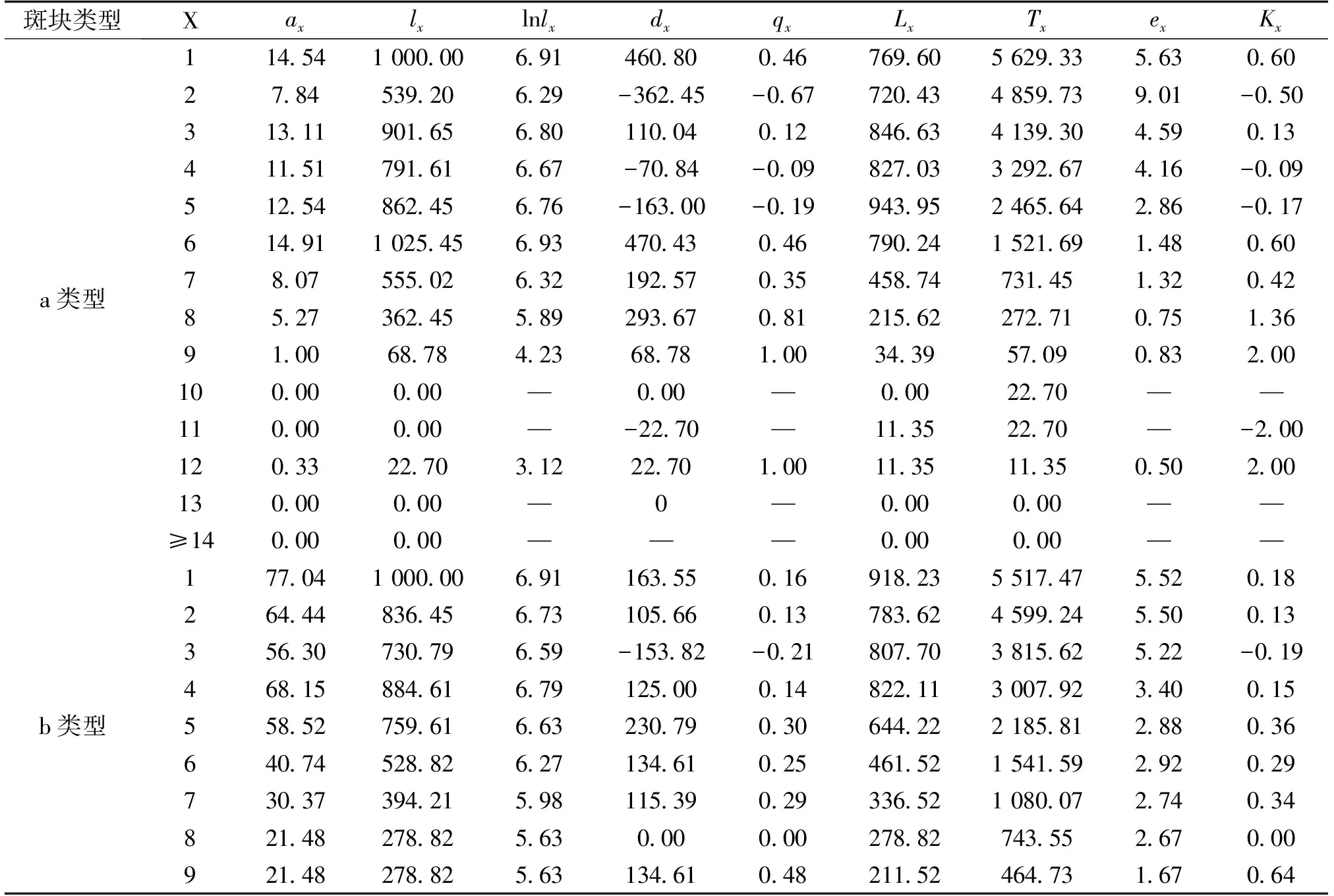
表2 3种样地类型中异翅独尾草种群的静态生命表
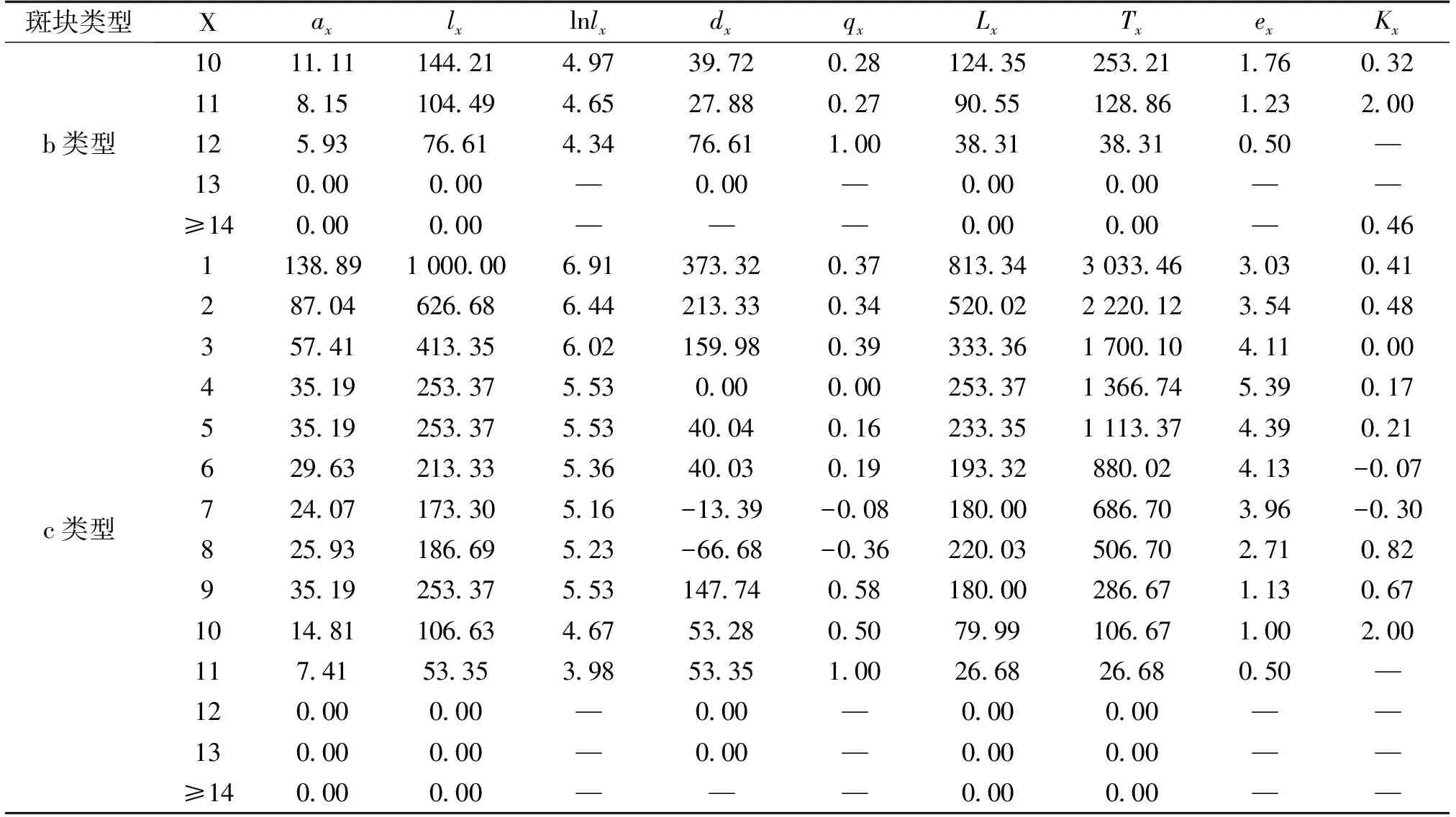
表2续
其中:x:龄级;ax:在x龄级内现有个体数;lx:在x龄级开始时标准化存活个体数(一般转换为1000);dx:从x到x+1龄级间隔期内标准化死亡数;qx:从x到x+1龄级间隔期间死亡率;Lx:从x到x+1龄级间隔期间还存活的个体数;Tx:从x龄级到超过x龄级的个体总数;ex:进入x龄级个体的生命期望或平均期望寿命;Kx:消失率。下表同。
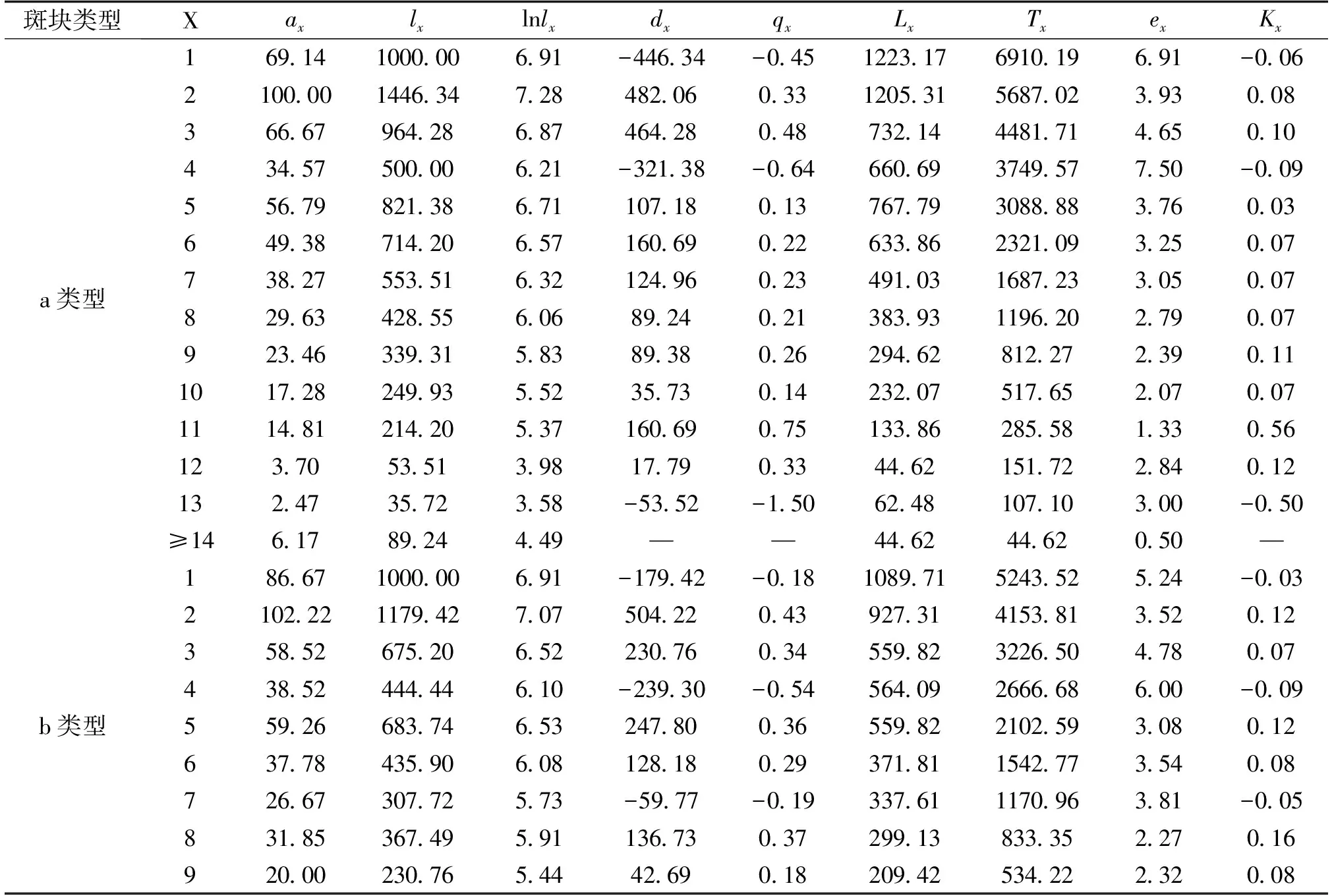
表3 3种样地类型中粗柄独尾草种群的静态生命表
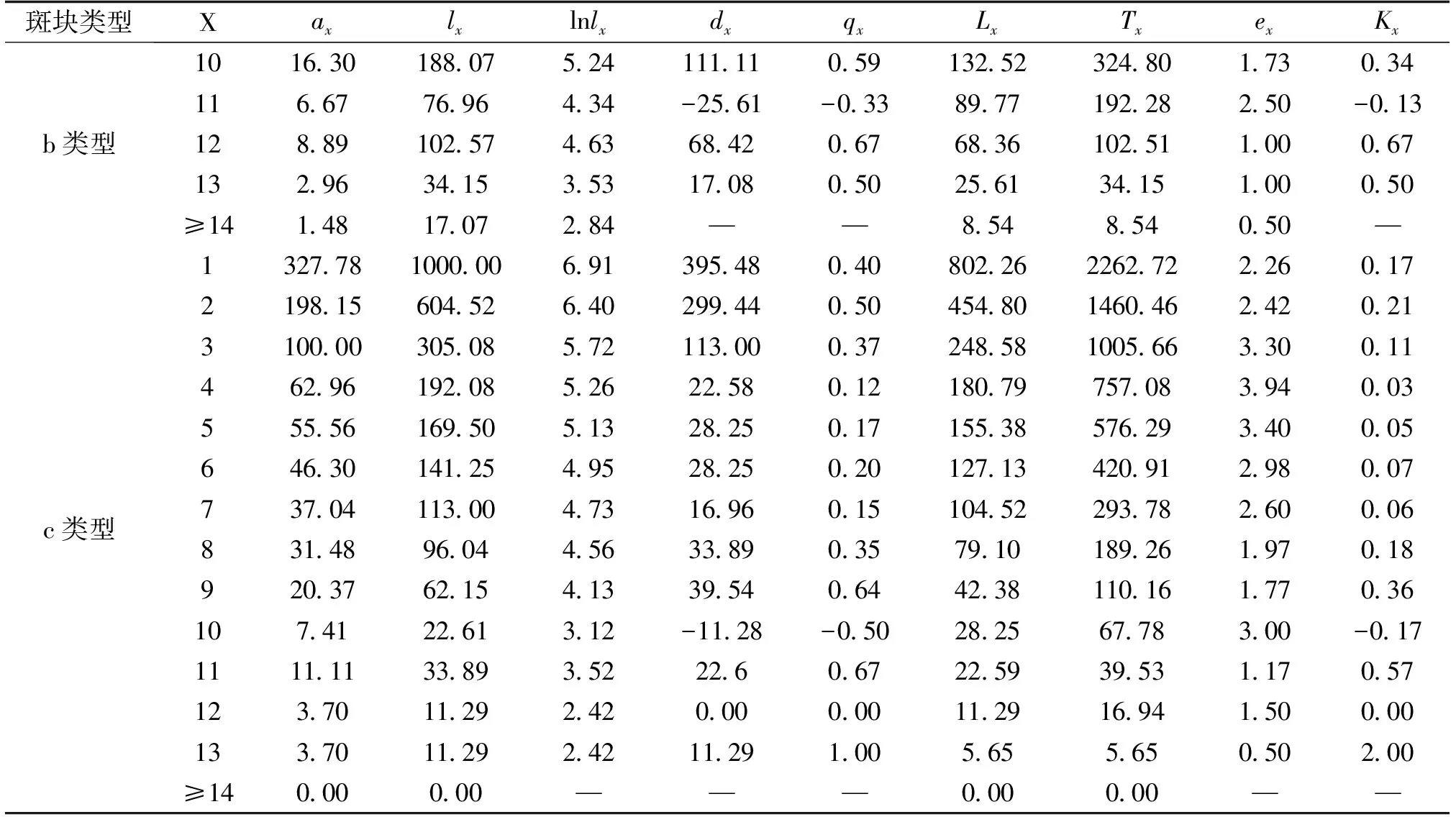
表3续
2.3 存活曲线和死亡曲线
绘制独尾草属2个同域种的存活曲线(图3A)和死亡曲线(图3B)。
从图3A可以看出,处于衰退期的植株在受到人类活动严重干扰的a类型样地中的异翅独尾草种群中缺失,在粗柄独尾草种群中植株数量少,说明人为干扰强度大而且持续时间较长;b类型样地中的粗柄独尾草种群的存活曲线则表现为先升高,在第2龄级达到最大存活数后开始下降,说明自然环境中幼苗的存活率较低;c类型样地中,两种独尾草属植物种群的存活曲线表现为DeeveyⅢ型,即幼苗时期的死亡率高。


图3 3种样地类型中两种独尾草属植物的种群存活曲线(A)与死亡曲线(B)
根据2种独尾草属植物的死亡曲线(图3B)可知,在a类型的样地中,2种独尾草属植物种群的死亡率不稳定,产生这种结果的原因是a类型样地的破碎程度高,受到了较强的人为干扰,种群遭到破坏,且各龄级植株数目不一,龄级缺失;在b类型样地中,粗柄独尾草的第1、4、7、11龄级的死亡率qx出现负值,这说明这些龄级的植株存活数小于其高一龄级的植株存活数,以及粗柄独尾草种群的低龄级植株较少,缺乏幼苗补充;在c类型样地中,从1-8龄级开始,2种独尾草种群的死亡率逐渐下降,第9龄级表现出一个小高峰,表明在第9龄级之后存活下来的个体,经历了一定强度的环境筛选。
3 讨论
异翅独尾草种群结构在生境破碎化程度高的a类型中呈现衰退型;在中度生境破碎化程度的b类型中为稳定至衰退型;在低生境破碎化程度的c类型中呈现增长型。a类型样地多为平地,受到放牧、农业活动的干扰,种群波动比较大,出现龄级缺失现象,而静态生命表中的负值也体现出了植株的缺乏状况。有研究表明[20],当以种子繁殖为主的植物种群在大量缺少幼苗的情况下,一般就断定该种群属于衰退型种群,因此在a类型样地中的独尾草种群在未来面临死亡或消失的可能性较大。b类型样地多位于坡中,受到开垦和道路的干扰,但是种群幼苗的存活率较低,这就导致了中等龄级的植株数量少,种群的更新比较困难,如果生境破碎化程度继续增加,异翅独尾草种群就可能出现衰退的风险。c类型样地位于坡顶,受干扰程度最低,种群处于发展过程中,这一结果说明生境破碎化程度的适当增加对种群的生存具有积极影响。
粗柄独尾草的年龄结构随着生境破碎化程度的增加也发生了变化。在生境破碎化程度高的a类型下表现为衰退型;中度生境破碎化程度的b类型中为稳定型;低生境破碎化程度的c类型中表现为增长型。在静态生命表中,也仅有几个数值为负值,且多位于中间龄级,这意味着种间竞争增加,后期竞争力度下降,种群趋于稳定。有研究表明,破碎化生境中的植物物种(除藤本植物外)的丰富度会较原生境中的下降40%[21]。与本研究结果一致,生境破碎化程度的增加,会降低种群数量,甚至会改变种群结构。
相较于异翅独尾草,粗柄独尾草则在种群结构方面表现出了较好的适应性。生长在中度生境破碎化程度的b类型样地中,异翅独尾草的年龄结构表现为稳定至衰退型,而同生境条件下的粗柄独尾草则表现为稳定型,这其中最主要的原因是粗柄独尾草具有更多的幼苗数量,能够应对人为干扰所造成的生境破碎化问题。根据安静[10]的研究可知,粗柄独尾草的种子千粒重明显小于异翅独尾草,且萌发率高于异翅独尾草。这意味着粗柄独尾草能够获得较大的生存空间以及生存几率。与此同时王海娟[9]还发现异翅独尾草会受到环境干扰后会使得种群的花粉活力降低,而粗柄独尾草在破碎化程度较高的生境中,会产生种翅变大、重量较轻的种子,这是植物适应环境的一种策略。但是植株下部的种子数量不会随破碎化程度的加重而下降,相反,植株下部的种子萌发率会随之提高。
4 结论
在古尔班通古特沙漠南缘中,不同的生境导致了异翅独尾草与粗柄独尾草种群之间年龄与动态的差异,并表现出不同的结构特征。通过对独尾草属2个同域种种群结构和动态特征的分析,结果表明:受人类扰动越强、生境破碎化程度越高的样地中的独尾草属植物种群趋于衰退的风险更大,但是在相同生境破碎化背景下,粗柄独尾草比异翅独尾草更能适应环境的变化。因此,结合独尾草属植物重要的生态意义以及本文的研究结果,认为:对于独尾草属植物的保护应结合野外生境中种群的生存现状,因地制宜地制定可实施的保护和恢复策略。
