生态因子对欧洲冬青光合特性的影响1)
2021-12-03李燕于冰辛建攀田如男
李燕 于冰 辛建攀 田如男
(南京林业大学,南京,210037)
欧洲冬青(Ilexaquifolium)又名英国冬青,冬青科(Aquifoliaceae)冬青属(Ilex)植物,原产于欧洲南部、中部、西北部,为北极第三纪孑遗物种[1-2]。该树种四季常青,树姿优美,果红时鲜艳美丽、经冬不落,可作绿篱、盆栽,也可孤植或丛植,在城市绿化、园林美化中具有广阔的应用前景。目前,国内外关于欧洲冬青的研究集中于地理分布、种群更新机制、药用价值等方面,关于其对环境因子的生理生态适应性研究较少[3-4]。研究表明,植物对环境的适应性最先表现于生理生态功能,如光合作用类型、水分利用程度等[5]。光合作用作为植物体内重要的代谢反应,是植物生长发育的基础。植物光合能力的大小不仅受内部生理因素(如基因)的影响,还受到大气温度(Ta)、空气相对湿度(RH)、光合有效辐射(PAR)、大气CO2摩尔分数(Ca)等外部环境因子的影响[6-11]。
南京地区属于亚热带季风气候,四季分明,与欧洲冬青原产地自然条件的差异会导致国内引进该树种时出现生长发育缓慢、观赏价值降低等现象[1]。本研究从光合角度出发,结合南京地区主要生态因子,研究不同季节欧洲冬青‘黄果’、‘阿拉斯加’的光合日变化特征,探究生态因子对其的影响,以期为欧洲冬青在亚热带地区的引种驯化、栽培、园林应用提供参考。
1 材料与方法
1.1 试验材料
试验材料为荷兰引进的2年生欧洲冬青‘黄果’、‘阿拉斯加’扦插苗。将植株栽植于直径30 cm的塑料盆内,每盆1株,置于南京林业大学园林国家级实验教学示范中心温室,于通风良好、阳光充足的环境中进行常规养护。
1.2 光合气体交换参数日变化测定
以生长健壮、大小一致的植株为试验材料,分别于晴天(3月22日、7月28日、11月3日、1月17日)08:00—18:00期间,利用CIRAS-2便携式光合仪对供试品种进行叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2摩尔分数(Ci)等参数的测定,并计算出气孔限制值(Ls):Ls=1-Ci/Ca。测定时,统一选取当年生新枝上部向阳方向且生长良好的成熟叶片(从上至下第3或第4片叶),并对测定叶片及植株进行挂牌标记。每个品种重复3次。
1.3 生态因子对净光合速率影响的测定
以生长健壮、大小一致的植株为试验材料,选择4—6月中晴朗无风的1 d,于08:30—11:30期间利用CIRAS-2便携式光合仪进行测定。
将光合测定系统的叶室温度设定为25 ℃,湿度设定为90%,CO2摩尔分数设定为380 μmol·mol-1,光强分别设定为2 000、1 600、1 400、1 200、1 000、800、600、400、200、175、150、100、75、50、0 μmol·m-2·s-1,共15个梯度,记录Pn。采用叶子飘等[12]建立的直角双曲线修正模型进行拟合,得到最大净光合速率(Pnmax)、光补偿点(Lc,p)、光饱和点(Ls,p)、表观量子效率(AQY)等光合特征参数[13-15]。
将光合测定系统的叶室温度设定为25 ℃,湿度设定为90%,光照强度设定为1 000 μmol·m-2·s-1,CO2摩尔分数分别设定为1 500、1 200、1 000、800、600、400、200、150、120、100、80、50、0 μmol·mol-1,共13个梯度,记录Pn。采用直线双曲线修正模型进行拟合,得到Pnmax、CO2补偿点(Cc,p)、CO2饱和点(Cs,p)、羧化效率(CE)等参数。
将光合测定系统的光照强度设定为1 000 μmol·m-2·s-1,叶室湿度设定为90%,CO2摩尔分数设定为380 μmol·mol-1,光合测定系统的叶室温度分别设定为5、10、15、18、20、21、22、23、24、25、28、30、35、38 ℃,共14个梯度,记录Pn。
将光合测定系统的光照强度设定为1 000 μmol·m-2·s-1,叶室温度设定为25 ℃,CO2流量设定为380 μmol·mol-1,光合测定系统的叶室相对湿度分别设定为90%、80%、70%、60%、50%、40%、30%、20%、10%,共9个梯度,记录Pn。
1.4 数据处理
使用SPSS 26软件进行非线性回归及单因素方差分析,采用Duncan法进行多重比较,使用Microsoft Excel软件进行数据整理、图表绘制。
2 结果与分析
2.1 不同季节欧洲冬青叶片光合参数日变化
2.1.1 净光合速率的日变化
Pn是衡量植物光合能力的重要指标,可以表征植物光合产物积累能力的高低。由图1可知,欧洲冬青‘黄果’、‘阿拉斯加’在南京地区出现不同程度的光合“午休”现象。在3月22日、11月3日、1月17日,‘黄果’、‘阿拉斯加’叶片Pn日变化曲线均呈双峰型,其峰值分别出现在10:00及14:00,波谷出现在12:00。1月17日12:00时,‘阿拉斯加’叶片的Pn比‘黄果’增加了270.34%(P<0.05),表明‘阿拉斯加’光抑制程度较弱。7月28日,‘黄果’、‘阿拉斯加’叶片光合“午休”时间为14:00,比其他3个时间段推迟了2个小时。由表1可知,虽然不同时间段欧洲冬青‘黄果’、‘阿拉斯加’叶片Pn日变化趋势相似,但日平均Pn存在一定差异。7月28日,‘黄果’、‘阿拉斯加’叶片日平均Pn最大,分别比其他3个时间段显著增加了25.67%~124.40%和54.58%~81.92%(P<0.05)。

图1 不同时间段欧洲冬青叶片净光合速率日变化

表1 不同季节欧洲冬青叶片日平均净光合速率
2.1.2 气孔导度的日变化
气孔是CO2进入及水分散失的门户,Gs的变化与光合作用、蒸腾作用密切相关。由图2可知,在3月22日、11月3日、1月17日,欧洲冬青‘黄果’、‘阿拉斯加’叶片Gs日变化规律和Pn一致,均为双峰型曲线。叶片Gs于10:00时出现第1个波峰,12:00时出现波谷,这与PAR、Ta增加,Ca、RH下降有关。7月28日,欧洲冬青‘黄果’、‘阿拉斯加’叶片Gs日变化趋势不同于其他3个时间段,其中‘黄果’叶片Gs日变化呈下降趋势;‘阿拉斯加’叶片Gs日变化呈倒置单峰曲线,于12:00时出现波谷(50.75 mmol·m-2·s-1)。

图2 不同时间段欧洲冬青叶片气孔导度日变化
2.1.3 蒸腾速率的日变化
从图3可知,欧洲冬青‘黄果’、‘阿拉斯加’叶片Tr日变化曲线为双峰型曲线。3月22日,‘黄果’、‘阿拉斯加’叶片Tr均在10:00、14:00达到峰值,其中‘黄果’峰值分别为4.46、3.06 μmol·m-2·s-1,‘阿拉斯加’峰值分别为4.75、2.71 μmol·m-2·s-1。中午12:00时,2种叶片的Tr出现波谷,这与叶片Gs下降有关,该现象有利于防止欧洲冬青体内水分耗散。11月3日、1月17日,‘黄果’、‘阿拉斯加’叶片Tr也表现出相似的日变化趋势。7月28日,‘阿拉斯加’叶片Tr日变化曲线为双峰型曲线,其最大值(4.48 μmol·m-2·s-1)出现在14:00;‘黄果’叶片Tr日变化曲线为单峰型曲线,于12:00达到最大值(4.79 μmol·m-2·s-1)。

图3 不同时间段欧洲冬青叶片蒸腾速率日变化
2.1.4 胞间CO2摩尔分数的日变化
由图4可知,3月22日、11月3日,欧洲冬青‘黄果’、‘阿拉斯加’叶片Ci日变化曲线为倒置双峰型曲线,分别于10:00、14:00出现波谷,这与叶片非气孔因素有关(图2)。7月28日,‘黄果’、‘阿拉斯加’叶片Ci日变化趋势与3月22日、11月3日的相似,但2个波谷均推迟2个小时。1月17日,‘黄果’、‘阿拉斯加’叶片Ci日变化曲线均为倒置单峰型曲线,于12:00达到最低值,分别为307.50、306.00 ppm。
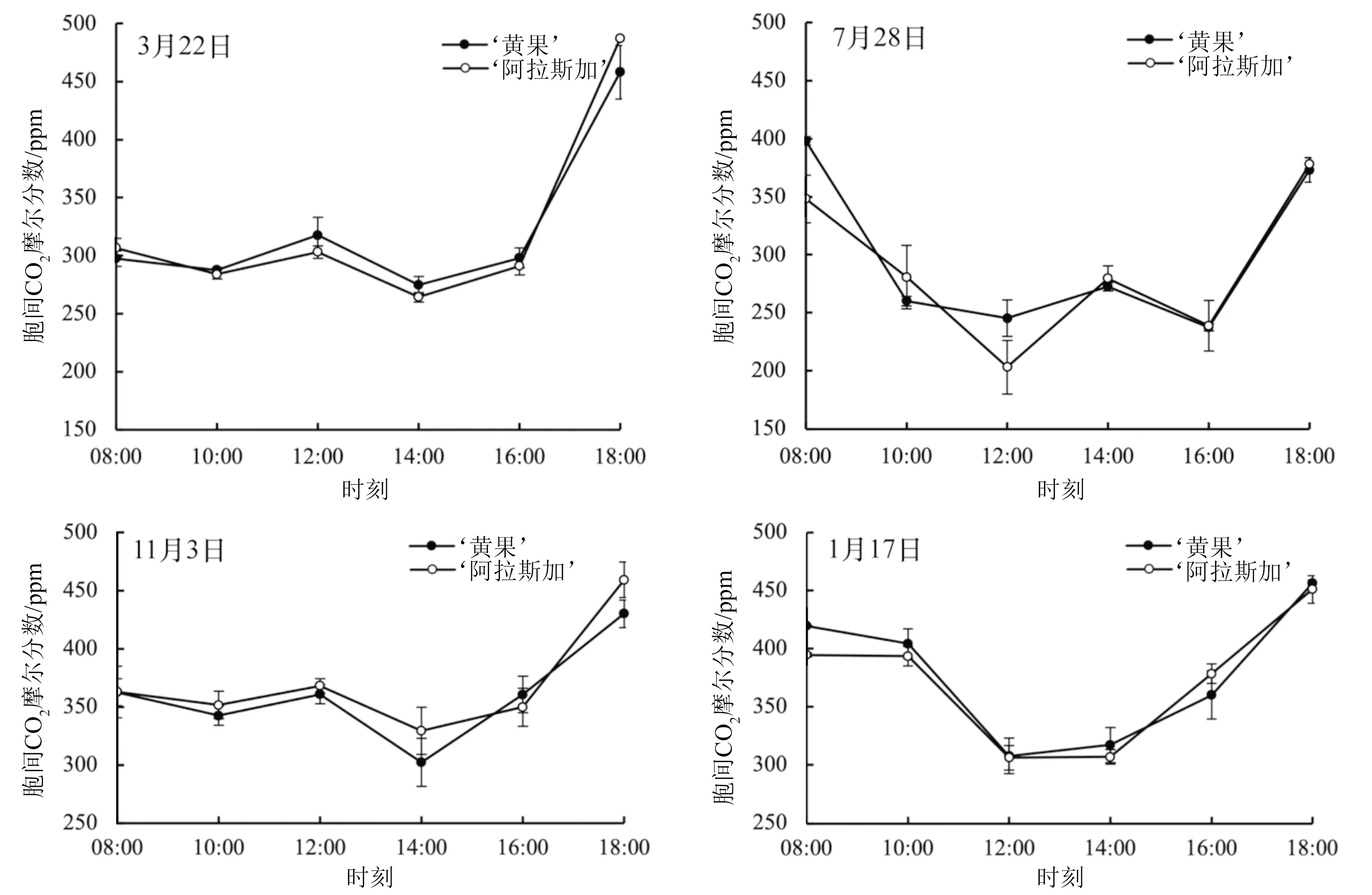
图4 不同时间段欧洲冬青叶片胞间CO2摩尔分数日变化
2.1.5 气孔限制值的日变化
Ls可以反映植物对大气CO2的利用效率,与Ci密切相关。从图5可知,在3月22日、7月28日、11月3日,欧洲冬青‘黄果’、‘阿拉斯加’叶片Ls日变化曲线为双峰型曲线,第1次波峰后,Ls均下降,与Ci变化趋势相反,此时非气孔因素是导致2个品种产生光抑制的原因。1月17日,‘黄果’、‘阿拉斯加’叶片Ls日变化曲线为单峰型曲线。

图5 不同时间段欧洲冬青叶片气孔限制值日变化
2.2 生态因子对欧洲冬青叶片净光合速率的影响
2.2.1 光照对叶片净光合速率的影响
AQY是光合作用中光能转化效率的一种量度,其值越高,表明植物对弱光的利用能力越强。由表2可知,欧洲冬青‘黄果’叶片AQY比‘阿拉斯加’的增加了6.45%(P<0.05),其对弱光的利用能力较强。Ls,p则表现出相反的变化趋势,说明‘阿拉斯加’叶片对强光环境的适应能力较强。与‘黄果’相比,‘阿拉斯加’叶片暗呼吸速率(Rd)、Lc,p、Pnmax均无明显变化(P>0.05)。

表2 欧洲冬青2个品种光合-光强响应曲线参数
2.2.2 CO2对叶片净光合速率的影响
CE受1,5-二磷酸核酮糖羧化酶/氧合酶活性及活化1,5-二磷酸核酮糖羧化酶/氧合酶质量分数的限制。Rd反映了植物维持自身生命活动所消耗有机物的含量。Pnmax是衡量叶片光合潜力的重要指标,反映光合电子传递、光合磷酸化活性。由表3可知,欧洲冬青‘黄果’叶片CE、Pnmax、Rd分别比‘阿拉斯加’高25.93%、23.34%、24.41%(P<0.05),说明‘黄果’叶片1,5-二磷酸核酮糖羧化酶/氧合酶活性较强、光合潜力较高,其叶片呼吸对光合产物的需求更多。与‘黄果’相比,‘阿拉斯加’叶片Cc,p、Cs,p均无明显变化(P>0.05)。‘黄果’、‘阿拉斯加’的Cs,p均远高于Ca(图1),这意味着可以通过增加CO2摩尔分数提高欧洲冬青叶片光合能力。

表3 2个欧洲冬青品种光合-CO2响应曲线参数
2.2.3 温度对叶片净光合速率的影响
由图6可知,欧洲冬青‘黄果’、‘阿拉斯加’叶片Pn对Ta的响应曲线均为双峰型曲线。随着Ta升高,‘黄果’叶片Pn逐渐升高,分别于18、24 ℃时达到峰值,峰值分别为8.33、9.03 μmol·m-2·s-1。‘阿拉斯加’叶片Ta对Pn的影响也表现出相似的变化趋势,分别于20、25 ℃时达到峰值,峰值分别为7.67、8.60 μmol·m-2·s-1。由上述结果可知,欧洲冬青‘黄果’、‘阿拉斯加’喜温暖气候。

图6 温度对2个欧洲冬青品种叶片净光合速率的影响
2.2.4 空气相对湿度对叶片净光合速率的影响
由表4可知,随RH的下降,欧洲冬青‘黄果’叶片净光合速率呈下降趋势。当RH为10%~70%时,其叶片Pn与RH为80%时相比,下降了10.16%~63.59%(P<0.05),‘阿拉斯加’叶片Pn也表现出相似的变化趋势,说明湿润环境有利于‘黄果’和‘阿拉斯加’光合作用。

表4 不同空气相对湿度时2个品种欧洲冬青的光合参数
3 讨论
3.1 不同季节欧洲冬青叶片光合参数日变化
在自然环境条件时,植物叶片光合参数的日变化可以反映光合作用与自身生理、外界环境变化间的关系[16]。Pn的大小反映植物同化作用的强弱及光合产物积累的多少,其不仅受植物生物学特性的影响,还受PAR、Ca、Ta、RH等多种生态因子的影响[17-18]。本研究中,不同季节欧洲冬青‘黄果’、‘阿拉斯加’均出现了明显的光合“午休”现象,表明其在光照过强时产生了光抑制。其中,春季(3月22日)、秋季(11月3日)光合“午休”期间,叶片Ci增加,Ls下降,夏玉米(Zeamays)[19]、欧洲卫矛(Euonymuseuropaea)‘八仙花’[20]等多种植物也发现了相似现象,表明非气孔因素是导致2个欧洲冬青品种出现光抑制的主要原因,这与3月22日、11月3日RH较小有关。冬季(1月17日)叶片Ci减少,Ls上升,意味着2个欧洲冬青品种光合“午休”现象与气孔限制密切相关[21]。与‘黄果’相比,‘阿拉斯加’在1月17日对光抑制敏感性较低,与Valladares et al.[22]对冬季田间条件下欧洲冬青光抑制敏感性的研究结果相一致。本研究中,冬季Ta月均值为5.8 ℃,最低值仅为-3 ℃,这会抑制叶片1,5-二磷酸核酮糖羧化酶/氧合酶活性,降低羧化效率,从而降低光合能力[23-25]。
康雅茸等[26]发现,夏季芦苇叶片Pn日变化曲线呈明显不对称的双峰型曲线,光合“午休”现象明显,与本研究中欧洲冬青‘黄果’、‘阿拉斯加’叶片Pn日变化规律相似,由此可推测‘黄果’、‘阿拉斯加’叶片Pn日变化曲线在7月28日也呈双峰型变化。此外,夏季(7月28日)光合“午休”期间,2个品种叶片Ci增加、Ls下降,说明非气孔限制是导致此时光合“午休”现象的主要原因,牡丹(Paeoniasuffruticosa)、桃(Prunuspersica)等植物也有类似现象[27-28],这是因为夏季(7月28日)中午高温(35 ℃)时,欧洲冬青叶片叶绿体结构受损、1,5-二磷酸核酮糖羧化酶/氧合酶活性降低,叶片暗呼吸、光呼吸加强[29-30]。因此,做好夏季降温措施有利于欧洲冬青的引种、栽培。
3.2 生态因子对欧洲冬青叶片净光合速率的影响
光响应曲线是研究植物光合能力的一种重要手段,可以反映植物对环境的适应能力[31-32]。AQY作为光合作用中光能转化效率的一种指标,其值一般在0.04~0.07[33]。本研究中,欧洲冬青‘黄果’、‘阿拉斯加’叶片AQY分别为0.033、0.031,可以看出2个品种叶片光能转化效率较低,对弱光的利用能力较弱。Lc,p、Ls,p作为植物耐阴性评价的重要指标,反映了植物对光照条件的要求及其适应性的强弱[34-35]。研究表明,阳生植物叶片Lc,p高于10 μmol·m-2·s-1,叶片Ls,p高于1 000 μmol·m-2·s-1[36-37],因此,欧洲冬青‘黄果’、‘阿拉斯加’均属于阳生植物,对环境适应能力强。
植物的碳同化受到羧化效率和1,5-二磷酸核酮糖(RuBP)再生速率的限制[38]。CO2响应曲线中初始斜率(即CE)与1,5-二磷酸核酮糖羧化酶/氧合酶活性、活化1,5-二磷酸核酮糖羧化酶/氧合酶质量分数呈正相关[33]。本研究中,欧洲冬青‘黄果’叶片CE比‘阿拉斯加’增加了25.93%(P<0.05),表明‘黄果’叶片中1,5-二磷酸核酮糖羧化酶/氧合酶活性较高及活化的1,5-二磷酸核酮糖羧化酶/氧合酶质量分数较多,对大气CO2有更高的光合利用效率[39]。当CO2处于饱和阶段时,Pnmax反映了光合机构由受1,5-二磷酸核酮糖羧化酶/氧合酶限制转变为依赖CO2受体-RuBP再生速率[40]。本研究中,欧洲冬青‘黄果’叶片Pnmax比‘阿拉斯加’明显增加了23.34%,说明‘黄果’叶片光合机构中RuBP的再生速率较高,对CO2的同化能力较强,有利于其光合产物的形成、积累[41]。
欧洲冬青原产于荷兰,属温带海洋性气候,Ta为0~22 ℃。本研究中,‘黄果’、‘阿拉斯加’叶片Pn分别在24、25 ℃达到最大值,证明了欧洲冬青喜温暖的生态学特性[3]。有研究发现,银杏(Ginkgobiloba)、樟树(Cinnamomamcamphora)、乐昌含笑(Micheliachapensis)、广玉兰(Magnoliagrandiflora)、玉兰(Magnoliadenudata)等多数植物叶片Pn对Ta的响应曲线为单峰型曲线,在35 ℃时Pn达到最大值[42],由此表明,欧洲冬青叶片光合作用对高温比较敏感。同时,本研究认为,湿润的大气条件可以促进欧洲冬青‘黄果’、‘阿拉斯加’叶片光合作用,这是其对原产地气候条件的适应性表现。
4 结论
欧洲冬青‘黄果’、‘阿拉斯加’属阳生植物,喜温暖、湿润环境。其中,‘阿拉斯加’对较强光照具有良好的适应能力,‘黄果’具有较好的光合能力。在春季(3月22日)、夏季(7月28日)、秋季(11月3日),非气孔因素是导致欧洲冬青‘黄果’、‘阿拉斯加’出现光抑制现象的主要原因;冬季(1月17日)叶片气孔导度下降,原因与‘黄果’、‘阿拉斯加’出现光合“午休”现象密切相关。在亚热带地区引种欧洲冬青品种时,应做好夏季降温、增湿等措施,以增强其适应能力。
