高盐稀态酱醪中功能性酵母菌的筛选、鉴定及发酵特性
2021-12-03王靖雯赵谋明张佳汇陈子杰冯云子
王靖雯,赵谋明,陈 涛,张佳汇,陈子杰,冯云子,*
(1.华南理工大学食品科学与工程学院,广东 广州 510641;2.上海太太乐食品有限公司,上海 201206)
酱油是一种起源于中国的传统发酵调味品,因其独特的风味受到了全世界人民的喜爱[1]。酱油发酵包含两个步骤,即制曲和酱醪发酵。制曲阶段以米曲霉的生长和作用为主,随着酱醪发酵阶段的推进,米曲霉的作用减弱,耐盐酵母菌和乳酸菌不断增殖,代谢形成丰富的物质,包括氨基酸类、糖类、糖醇类、有机酸类、醇类和酯类等挥发性化合物,对提高酱油的风味品质具有重要贡献[2-4]。其中,耐盐酵母菌对酱油挥发性物质的积累具有重要作用[5]。
随着风味感官组学相关技术的发展以及酱油香气活性物质的鉴定,许多研究逐渐转向酵母对酱油挥发性风味物质的影响。其中报道次数较多的有耐盐生香酵母包括鲁氏结合酵母(Zygosaccharomyces rouxii)、季也蒙毕赤酵母(Pichia guilliermondii)和易变假丝酵母(Candida versatilis)等。Yao Shangjie等[6]研究发现单独接种鲁氏结合酵母可以提升苯乙醇、3-甲基-1-丁醇、2-甲基-1-丙醇等醇类物质的含量。Wah等[7]从酱油发酵1 个月的酱醪中分离得到嗜盐的季也蒙毕赤酵母,发现其单独接种发酵后的酱油中愈创木酚和4-乙基愈创木酚的含量明显提升。张玲[8]研究新发现添加易变假丝酵母可以促进乙醇、乙酸、4-乙基愈创木酚、甲基吡嗪、苯乙醛和4-羟基-2(5)-乙基-5(2)-甲基-3(2H)-呋喃酮等风味活性物质含量的提升。除单菌种发酵对风味影响的研究外,功能菌株混合培养的应用逐渐成为热点。Singracha等[9]发现鲁氏结合酵母与季也蒙毕赤酵母共同接种发酵的酱油中乙醇、2-甲基-1-丙醇、4-羟基-2,5-二甲基-3(2H)-呋喃酮和3-羟基-2-甲基-4H-吡喃-4-酮(麦芽酚)等物质含量明显高于单独接种鲁氏结合酵母发酵的酱油。通过总结国内外文献[10-12]中鉴定酱油酵母菌的种属信息,共统计18 个菌属,关于酱油酵母菌的相关研究有待深入,特别是一些含量较低的功能菌种有待挖掘。
因此,本研究拟从高盐稀态酱油酱渣中筛选分离功能酵母菌,结合基因测序技术确定其种属,选择代表性菌株进行生理生化鉴定并探究其生长特性,将分离纯化的酵母菌种单独接种到酱油体系培养,结合感官评价和气相色谱-质谱(gas chromatography-mass spectrometry,GC-MS)分析判断其风味代谢特点,以期为酱油酵母资源的开发及发酵风味调控提供理论指导。
1 材料与方法
1.1 材料与试剂
高盐稀态酱油发酵终点的酱渣样品,取自广东省某酱油厂。
氢氧化钠、NaCl、盐酸、葡萄糖、硫酸镁、甲基红指示剂、溴酚蓝指示剂 国药集团化学试剂有限公司;乙醇、2-辛醇、2-甲基-3-庚酮(均为分析纯) Sigma(上海)有限公司;PDA固体培养基、麦芽汁液体培养基、麦芽汁琼脂培养基 广东环凯有限公司。
氮源同化培养基:葡萄糖10.0 g,(NH4)2SO4(其他氮源)5.0 g,KH2PO40.85 g,K2HPO40.15 g,MgSO40.50 g,NaCl 0.10 g,CaCl20.10 g,2 g/100 mL琼脂粉,蒸馏水加至1 L。
碳源同化培养基:(NH4)2SO45.0 g,KH2PO40.85 g,K2HPO40.15 g,MgSO40.50 g,酵母浸膏0.88 g,葡萄糖(其他碳源)7.0 g,蒸馏水加至1 L。
1.2 仪器与设备
EL204/EL3002电子天平 梅特勒-托利多仪器有限公司;THZ-82A恒温振荡器 常州澳华仪器有限公司;HH-8数显电子恒温水浴锅 江苏省金坛市宏华仪器厂;UV-2100紫外-可见分光光度计 上海尤尼柯科学仪器有限公司;Finnigan Trace DSQ II气相色谱仪、Trace GOLD TG-5SilMS毛细管色谱柱 美国Thermo Fisher科技公司。
1.3 方法
1.3.1 酵母菌的分离、纯化
酵母菌的分离采用稀释平板涂布法。将高盐稀态酱油酱渣样品进行梯度稀释,取0.1 mL样品稀释液(10-1和10-2)涂布于含10 g/100 mL NaCl的PDA固体培养基中,每个梯度进行2 个重复实验,置于28 ℃培养2~3 d,挑取平板形态近似酵母且差异较大的单菌落进行镜检,将初步镜检为酵母的菌落进行多次平板划线,获得纯培养菌株。纯化后的菌株分别接种于斜面试管和甘油管中,置于-20 ℃进行保藏。
1.3.2 基因测序
1.3.2.1 基因组DNA提取
将分离得到的纯培养酵母菌株送至北京六合华大基因科技有限公司武汉分公司进行鉴定。酵母菌体依次通过十二烷基硫酸钠、蛋白酶K和十六烷基三甲基溴化铵混匀水浴,离心后将沉淀溶于70%乙醇溶液,反复离心后得到的沉淀溶解于含RNase的纯化水中,于4 ℃溶解过夜。
1.3.2.2 ITS rDNA序列分析及系统发育树构建
将提取得到的酵母菌模板DNA进行聚合酶链式反应(polymerase chain reaction,PCR)扩增。使用ITS rDNA的通用引物(ITS1:5’-TCCGTAGGTGAACCTGCGG-3’;ITS4:5’-TCCTCCGCTTATTGATATGC-3’);采用30 μL PCR体系:17.8 μL H2O,3 μL Buffer,2 μL dNTP,3 μL Primer1,3 μL Primer2,1 μL模板DNA和0.2 μL酶。PCR条件:95 ℃预热1 min,95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,35 个循环后72 ℃末端延伸10 min,12 ℃保存。取3 μL PCR扩增产物,使用1%琼脂糖凝胶进行电泳检测,然后对PCR产物进行上机测序。使用BLAST将测序结果在美国国立生物技术信息中心(National Center for Biotechnology Information,NCBI)数据库进行同源性比对,并通过MEGA X软件构建系统发育树。
1.3.3 酵母菌生长曲线的绘制
将已鉴定且活化后具有代表性的酵母菌株以3%接种量分别接种于麦芽汁液体培养基,于28 ℃、120 r/min培养72 h,在0~72 h内每隔3 h取样测OD600nm值,绘制生长曲线。
1.3.4 酵母菌碳、氮源利用情况分析
参照《酵母菌的特征与鉴定手册》[13]对分离得到的酵母菌株进行碳、氮源同化实验。
碳源同化实验:将菌种用无菌水制成菌悬液,调整细胞浓度至半透光,取80 μL菌悬液接入碳源同化培养基中,28 ℃培养3~7 d后观察菌株生长情况。所用碳源包括葡萄糖、蔗糖、乳糖、果糖、麦芽糖、可溶性淀粉、木糖和柠檬酸。
氮源同化实验:将1 mL经饥饿培养的菌悬液与氮源同化培养基混匀凝固,蘸取不同氮源化合物点在平板上,观察菌株生长情况。所用氮源包括蛋白胨、尿素、硫酸铵和硝酸钠。
1.3.5 酵母菌耐盐性比较分析
将筛选的酵母菌株分别接入含有0、5、10、15、20、25 g/100 mL NaCl的麦芽汁培养基中,28 ℃、120 r/min培养72 h,观察菌株生长情况,测定菌悬液OD600nm值,比较不同菌株之间的耐盐性。
1.3.6 酵母菌酱油发酵体系
将未灭菌高盐稀态酿造酱油的原油在90 ℃水浴加热灭菌15 min,获得灭菌酱油,将分离得到的各个纯培养酵母菌株,分别在麦芽汁液体培养基中培养48 h后获得酵母种子液,以105CFU/mL的接种量分别接种于灭菌酱油中,静置于常温发酵30 d后,对发酵终点的酱油样品进行喜好度感官评分和风味测定。
1.3.6.1 感官评定
由7 位经过挑选、培训的感官评价人员(3 男4 女,20~31 岁)组成感官品评小组,对不同酵母发酵样品进行感官评价,20 mL样品装入50 mL嗅闻瓶中,感评人员以对照组(未添加酵母菌的酱油)为0 分,分别对样品进行喜好度评分,分值区间为-5(非常不喜欢)~5(非常喜欢),最终结果取感官得分平均值。感官实验过程中,样品嗅闻前需带盖稍微摇匀后静置片刻,准备好后开盖嗅闻并评分。
1.3.6.2 挥发性物质的固相微萃取
挥发性化合物萃取的方法参照冯云子[2]已优化的条件,并稍作调整。将酱油原油样5 mL添加于顶空进样瓶中,用NaCl调整样品盐质量浓度至270 g/L,加入20 μL内标(1.724 mg/L 2-甲基-3-庚酮甲醇溶液),使用75 μm CAR/PDMS萃取头进行挥发性成分萃取。
采用TR-5ms弹性石英毛细色谱分析柱(30 m×0.25 mm,0.25 μm),载气为高纯氦气(1.0 mL/min),采用电子电离离子源,电子能量70 eV,电子倍增器电压350 V,离子扫描范围m/z33~350,扫描速率3.00 scans/s,离子源温度250 ℃,传输线温度250 ℃,分流比10∶1。程序升温条件:起始温度40 ℃,保持2 min,以5 ℃/min升到120 ℃,保持2 min,再以7 ℃/min升至220 ℃,保持5 min。
1.4 数据分析
GC-MS化合物的定性分析通过NIST08谱库及对比标准物质科瓦茨保留指数(retention index,RI)完成,RI值通过正构烷烃(C6~C33)计算得到。利用SPSS软件(version 24.0,Chicago,Ill.,美国)进行方差分析,并通过Duncan进行差异显著性分析(P<0.05,差异显著),Origin(version 2018, OriginLab Corporation,Northampton,227 MA,美国)进行数据可视化处理;基于挑选出的关键香气活性化合物,采用XLSTAT(version 2019,Addinsoft,New York,美国)进行主成分分析。
2 结果与分析
2.1 酵母菌株基因鉴定结果
从酱渣分离菌株中挑选代表性的酵母菌进行基因测序鉴定,所有酵母的ITS rDNA基因序列系统进化树如图1所示。共分离得到14 株酵母,总计3 个菌属和4 个菌种,包括9 株假丝酵母属(Candida),3 株粉状米勒氏酵母属(Millerozyma)和2 株红酵母属(Rhodotorula)。

图1 酱醪酵母ITS rDNA基因序列系统进化树Fig.1 Phylogenetic tree of moromi-derived yeast based on their ITS rDNA genes
2.2 酱醅酵母菌落形态观察结果
鉴定的14 株酵母菌归属于4 个种,因此各挑选1 株代表性菌株进行生理生化特性实验,以了解各类菌种的特性,测定结果如表1和图2所示。镜检形态表明,粉状米勒氏酵母(M.farinosa)以瓜子椭圆形的形态为主,而胶红酵母(R.mucilaginosa)、近平滑假丝酵母(C.parapsilosis)和似平滑假丝酵母(C.metapsilosis)均以椭圆形为主。平板形态观察结果表明,胶红酵母表现为橙色圆形且表面隆起的小菌落,近平滑假丝酵母和似平滑假丝酵母均表现为白色圆形小菌落,表面湿润,均符合《酵母菌的特征与鉴定手册》[13]中菌种形态特征。而粉状米勒氏酵母表现为表面干燥褶皱,边缘不齐且不隆起的圆形白色大菌落,与Maresova等[14]研究该菌种的形态一致。

表1 4 株酱醪酵母的菌落特征及其生理生化测定结果Table 1 Colony features and biochemical characteristics of four strains of moromi-derived yeast

图2 4 株酱醪酵母的菌落平板及镜检形态Fig.2 Colony and microscopic morphology of four strains of moromi-derived yeast
2.3 碳、氮源利用情况分析
4 株代表性酵母菌株碳、氮源利用情况如表2所示。以葡萄糖、蔗糖、果糖、麦芽糖、可溶性淀粉作为碳源时,各菌株均能正常生长;但乳糖、木糖和柠檬酸作为碳源时,各酵母菌株生长情况不一致。在乳糖培养液中,各酵母菌株均不能生长,而在木糖培养液中只观察到胶红酵母和粉状米勒氏酵母的增殖;此外,只有粉状米勒氏酵母能够利用柠檬酸繁殖,与Tolieng等[15]报道该菌种利用碳源的情况一致。在氮源中,蛋白胨和尿素培养液中均能观察到各酵母菌株的生长,且近平滑假丝酵母和似平滑假丝酵母均不能同化硝酸钠,而胶红酵母不能同化硫酸铵及硝酸钠;与《酵母菌的特征与鉴定手册》[13]中各菌种的同化结果相符。
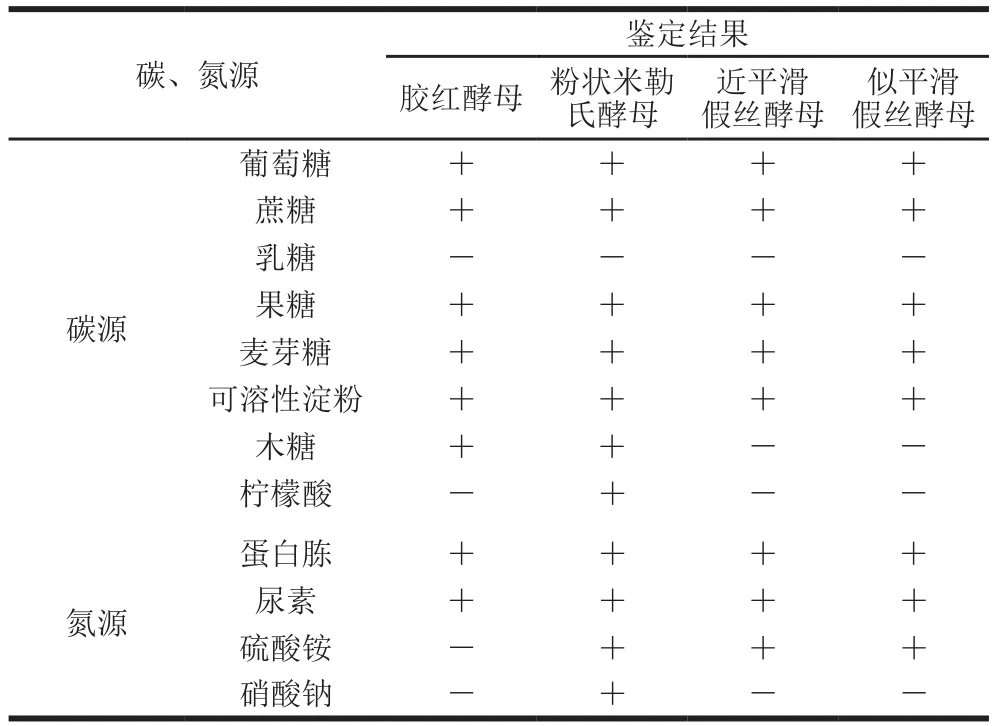
表2 碳、氮源对4 株酱醪酵母菌生长的影响Table 2 Effects of different carbon and nitrogen sources on the growth of four strains of moromi-derived yeast
2.4 酵母菌株生长曲线及耐盐性分析
由图3a可知,似平滑假丝酵母菌生长活力最强,3~18 h处于对数生长期,20 h以后为稳定期。此外,粉状米勒氏酵母和近平滑假丝酵母的生长规律相类似,3~25 h为对数生长期,25 h以后为稳定期。胶红酵母的生长曲线则明显区别于其他3 株酵母,进入对数生长期较晚,表现为0~13 h处于延迟期,13~45 h为对数生长期。
高盐稀态酱油发酵过程中NaCl质量浓度在17~18 g/100 mL左右,菌株的耐盐性是酱油功能菌株发挥作用的一个重要基础。图3b结果表明,在0~5 g/100 mL的NaCl质量浓度下,各菌株均能正常繁殖,当NaCl质量浓度高于10 g/100 mL时,近平滑假丝酵母和似平滑假丝酵母的生长均受到抑制,NaCl质量浓度越高,生长受抑制的程度越大,在NaCl质量浓度高于15 g/100 mL时,粉状米勒氏酵母的生长受到明显抑制,结果表明粉状米勒氏酵母的耐盐性强于近平滑假丝酵母,与Suzuki[16]研究中粉状米勒氏酵母耐盐性较强的结果一致。而胶红酵母在高盐环境下生长趋势的变化不明显,能够保持稳定增长。
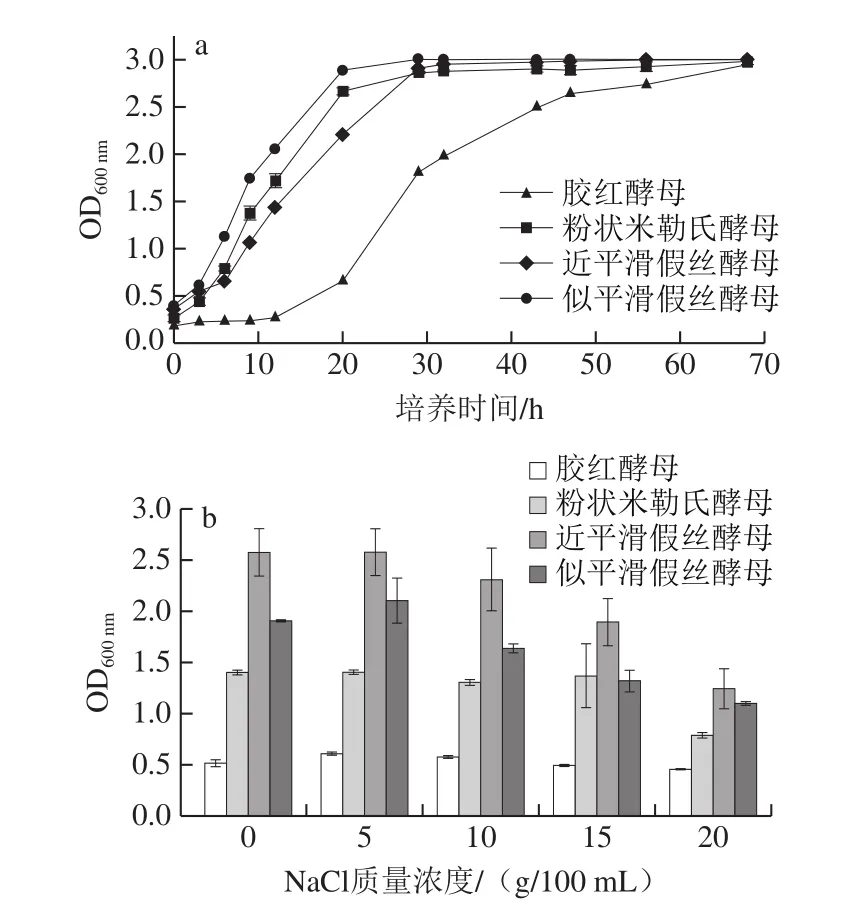
图3 酱醪酵母的生长曲线(a)和耐盐性(b)Fig.3 Growth curves (a) and salt tolerance (b) of four strains of moromi-derived yeast
2.5 感官评价和GC-MS分析结果
由图4A可知,共有8 株酵母发酵后的酱油气味喜好度得分为正值,包括4 株似平滑假丝酵母、2 株胶红酵母和2 株近平滑假丝酵母,其中似平滑假丝酵母SWJS W320和胶红酵母SWJS W201评分最高。而6 株酵母发酵后的样品得分为负值,包括3 株粉状米勒氏酵母和3 株近平滑假丝酵母,其中粉状米勒氏酵母(SWJS W214、SWJS W323、SWJS W213)的感官评分最低,风味喜好度显著低于其他菌株(P<0.05),其繁殖对酱油风味有明显的负面效果。然而,彭东等[17]研究认为粉状米勒氏酵母发酵后具有水果香和麦芽香,是具有潜力的酱油酿造生香酵母,与本研究结果不一致,这可能是由于嗅闻样品制备方法的差异引起,彭东等[17]研究中,感官评价样品采用160 g/L NaCl豆芽汁液体培养基接种培养7 d制备,而本研究直接将微生物接种到酱油中进行嗅闻,更能还原其在酱油发酵中的作用效果。
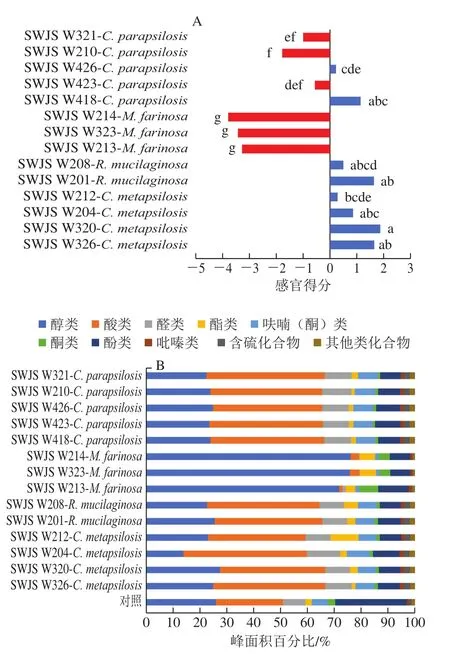
图4 14 株酵母菌发酵样品感官得分(A)和挥发性组分类别组成(B)Fig.4 Sensory scores (A) and volatile components (B) of soy sauce samples fermented with 14 yeast strains
如图4B所示,GC-MS共鉴定的105 种挥发性化合物可分为10 大类,包括酯类(21 种)、酮类(16 种)、醛类(15 种)、呋喃(酮)类(13 种)、吡嗪类(10 种)、含硫类化合物(7 种)、酸类(6 种)、醇类(7 种)、酚类(3 种)和其他类化合物(7 种)。对照本团队前期酱油挥发性物质检出总结[2],大部分化合物已有相关报道[18-19]。整体而言,3 个粉状米勒氏酵母发酵样品(SWJS W214、SWJS W323、SWJS W213)挥发性物质组成明显不同于其他菌种,醇类平均占比为74.4%,酸类仅2.7%,此外,酮类物质占比明显提升,这表明粉状米勒氏酵母的代谢特性较为特殊[20];该菌在酱油中的繁殖改变了酱油原有风味的平衡和特性,可能是该系列样品感官评分最低的原因。另外的11 个样品醇和酸在所有挥发性成分中占比最高,平均占比分别约为19.6%和43.4%,且样品组中酸、醛、呋喃类物质百分比含量均高于对照组,酚类和含硫化合物低于对照组。
基于各样品中48 个关键香气活性化合物的峰面积结果,对其进行主成分分析。如图5A所示,因14 个酵母发酵样品在图中分布象限不同,可将其分为2 个样品组。样品组1包括3 株粉状米勒氏酵母发酵样品,位于第2、3象限,样品组2包括胶红酵母、近平滑假丝酵母和似平滑假丝酵母发酵样品,均分散在第4象限,且与落在第1象限的对照组距离较远。
如图5B所示,有17 种挥发性物质位于第2、3象限,主要包括1,2,3-三甲氧基苯、2-壬酮、2-辛酮、2-甲氧基苯酚、2-戊酮、3-甲基丁酸,苯乙醇、3-甲基-1-丁醇、乙酸甲酯等。与对照组相比,这些物质在粉状米勒氏酵母中含量明显提升,其中2-辛酮和2-壬酮含量均提升约60 倍,且1,2,3-三甲氧基苯仅在该菌种对应样品中检出,被报道具有异味特征[2]的3-甲基丁酸也提升约5 倍,这一结果可能是造成感官得分较低(图4)的主要原因。有研究表明α-酮酸在氨基转移酶作用下形成相应的酮[21],酮类物质含量大幅提高,侧面说明粉状米勒氏酵母中该酶活性较强。另外,粉状米勒氏酵母发酵样品中苯乙醛含量降低而苯乙醇含量明显增加,前人研究报道中苯乙醛在醇脱氢酶作用下还原形成苯乙醇[22],说明该菌种可能对这一过程具有明显促进作用,与Choi等[23]研究粉状酵母发酵饮料中苯乙醇含量较高的结果一致。综上,粉状米勒氏酵母的代谢特点具有明显差异,且大量繁殖时会产生不良风味。
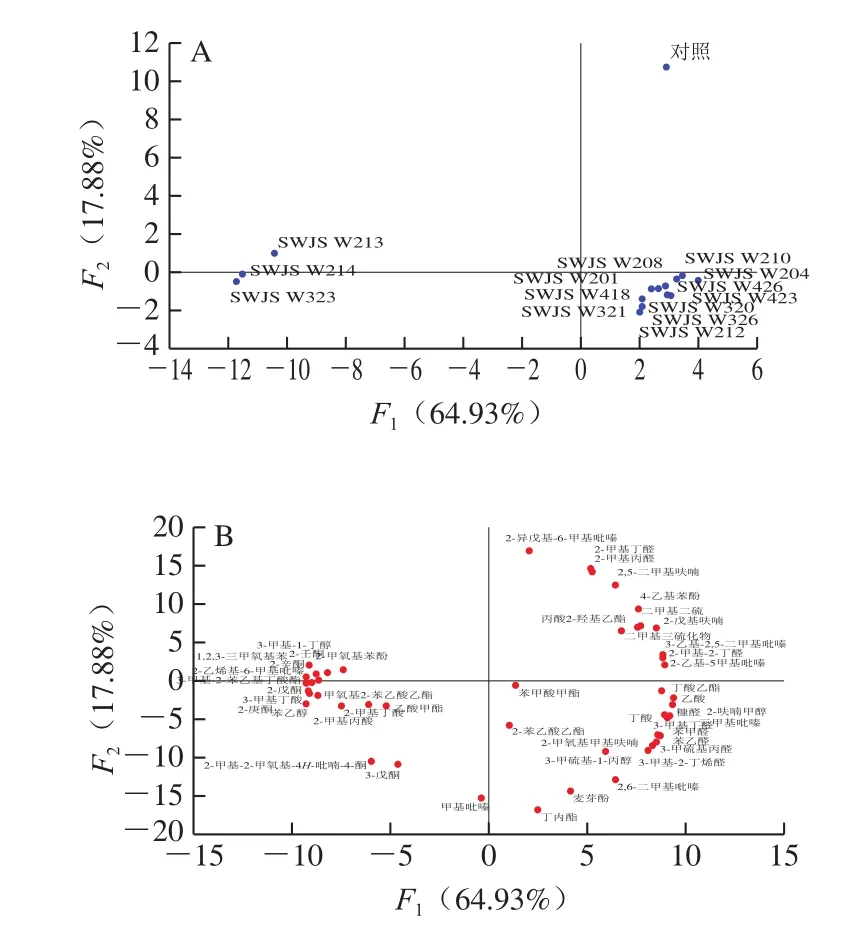
图5 酵母单独发酵样品挥发性物质的主成分分析得分图(A)和载荷图(B)Fig.5 PCA score plot (A) and loading plot (B) of volatile substances in soy sauce samples fermented using pure yeast strains
此外,2,6-二甲基吡嗪、3-甲硫基丙醛、3-甲硫基-1-丙醇、2-苯乙酸乙酯、苯乙醛等物质均落在第4象限。相对于对照组,近平滑假丝酵母可以提升甲硫醇、3-甲基-2-丁烯醛、2,6-二甲基吡嗪等物质的含量。而胶红酵母和似平滑假丝酵母可以提升苯乙醇、3-甲硫基丙醛、3-甲硫基-1-丙醇、2-苯乙酸乙酯、苯甲醛和2,6-二甲基吡嗪等关键风味物质的峰面积含量,这些物质均被报道为重要的活性风味成分[24-25],其中3-甲硫基丙醛和2-苯乙酸乙酯被报道分别赋予独特的烤土豆香[26]和玫瑰花香[27],与感官得分结果(图4)相符。胶红酵母被报道具有较高的油脂合成能力,可用于生产类胡萝卜素和类脂化合物[28-29]。综上所述,胶红酵母和似平滑假丝酵母具有一定酱油生香酵母的开发潜力。
3 结 论
从高盐稀态酱油酱渣中筛选分离得到14 株功能酵母菌,包括胶红酵母、粉状米勒氏酵母、近平滑假丝酵母和似平滑假丝酵母4 个菌种类型,选取代表性菌株研究菌落形态和生理生化特征。通过酱油体系菌株单独发酵产香并结合嗅闻感官评价,鉴定出对酱油风味具有负面影响的粉状米勒氏酵母和近平滑假丝酵母。特别是粉状米勒氏酵母,其菌落表面干燥褶皱,是酱油风味的有害酵母,其大量繁殖将大幅提高酱油中2-辛酮、2-壬酮和1,2,3-三甲氧基苯的含量,带来明显的不良风味。另外,筛选到对酱油风味具有提升潜力的胶红酵母和似平滑假丝酵母,可以提高苯乙醇、2-苯乙酸乙酯、2,6-二甲基吡嗪和3-甲硫基丙醛等风味活性物质含量。本研究仅考虑了单菌种的风味贡献效果,而在实际酱油发酵体系中,还需要考虑与其他菌群的交互作用,如常用的生香鲁氏结合酵母等。因此,具有潜力的风味功能酵母菌将进一步加入到酱油发酵过程中进行风味贡献验证,并且在酿造过程中如何提升有益菌种的作用,同时抑制有害酵母的繁殖代谢值得进一步深入研究。
