大花铁线莲花芽分化及花器官发育特性1)
2021-12-03李海微张永胜王非
李海微 张永胜 王非
(东北林业大学,哈尔滨,150040)
目前,我国园林绿化植物资源丰富,但高寒地区可用于垂直立体绿化的藤本植物资源有限,种类单一[1]。铁线莲属植物具有花期长,适应性强和较高的观赏性,被誉为“攀缘植物皇后”[2]。大花铁线莲因花朵硕大、花色丰富、耐寒耐旱,而常常用于栅栏、屋顶花园、廊架以及亭子的绿化点缀[3]。作为东北地区固有的野生藤本资源,大花铁线莲兼具较高的观赏特性和抗性,是垂直绿化的优良材料。故本试验选用大花铁线莲进行研究,一方面开发对野生资源的园林利用,充分发挥其生态效益和观赏价值;另一方面也可以扩大高寒地区垂直绿化应用的藤本植物种类的选择。
花芽分化是植物开花生物学的重要特征之一,而雌雄配子体的发育是花芽分化的一个延续过程。其过程的研究对于花期调控、提高植物开花数量和质量以及植物的遗传育种工作开展具有重要的意义[4-7]。目前,对大花铁线莲的研究主要集中在花部特征、花粉活力、繁育系统等方面[8-9]。花芽分化方面的研究鲜有,仅限于大花铁线莲3个花被类型的重被花起源方式的探索试验上[10],且并未对大花铁线莲生殖生物学方面进行探讨。因此,本试验针对大花铁线莲的野生栽培种的花芽分化过程进行观测和分析,同时在雌雄配子体发育方面进行了探索和研究,从而为大花铁线莲的花期调控和杂交育种等方面研究奠定一定的基础。
1 材料与方法
试验材料是从吉林省引至哈尔滨栽培了3年的大花铁线莲(Clematispatens)。试验地点为黑龙江省哈尔滨市香坊区东北林业大学园林学院实验室内进行,试验时间为2017年4月份至2018年6月份。
花芽采集的时间为4—6月上旬。材料每5 d进行1次采集(在花芽日变化不显著时),或者每2 d取材1次(花芽变化较快的时期)。材料采集之后立即用FAA固定液进行固定,于冰箱中4°固定24 h以上,之后放到酒精中进行保存。
于2017年4月底至6月初,每隔3 d,于每日08:00—10:00进行顶生单花的采集,将各个发育时期的大小各异的雌雄花蕾采集过后用FAA固定液固定并进行保存。
采用常规石蜡法进行制片。用LEICA RM 2255切片机将材料切割成若干个薄片,薄片的厚度8~10 μm。然后,用番红对薄片进行染色,中性树胶封片,最后将封制好的材料放在LEICA DM 2255显微镜下观察和拍摄。从材料采集到图片拍摄,需要试验的反复操作,直至得到该生长期的图像,重复图像择优。应用Excel 2007和SPSS 22.0对数据进行处理分析。
2 结果与分析
2.1 大花铁线莲花芽分化过程
如图1所示,大花铁线莲花芽分化分5个时期,即营养生长期、花芽分化初期、萼片原基分化期、雄蕊原基分化期、雌蕊原基分化期。大花铁线莲4月下旬便开始萌动进行营养生长,萌动5~7 d时花芽开始分化,大花铁线莲花芽分化主要是发生在顶芽,但是也存在一些发生在叶腋的特殊情况(图1T),这种特殊情况是极少存在的。在5月中旬左右,大花铁线莲进入开花期,6月上旬开始结实,在这期间花芽分化继续进行,为群体花期的延长提供了途径。
营养生长期:大花铁线莲在植株萌动后开始营养生长,此时生长锥细胞体积较小、排列整齐且分裂旺盛,整个生长锥头部较尖(图1A)。在外观上,大花铁线莲顶芽幼嫩,且外层包被多层叶芽,体积较小。
花芽分化初期:在分化初期,细胞体积变大,排列整齐(图1B),生长锥的横纵茎显著变长,其营养生长锥渐渐发展成阔圆、平坦的形状,细胞核较为明显。
花萼原基分化期:营养生长锥在此时期继续生长,与此同时,花原基的两侧逐渐分化出萼片原基,形似两个对称的突起(图1C)。随后萼片原基内细胞不断分裂,萼片原基分化为萼片,逐渐包住萼片内部的花芽原基(图1D-G)。
雄蕊原基分化期:随着花萼原基的不断生长,大花铁线莲生殖生长锥也在生长,体积变大(图1H)。细胞继续分裂,生殖生长锥基部两侧细胞分裂旺盛且排列紧密(图1I),随后分化出雄蕊原基(图1J),发育的顺序是由下而上呈中心对称进行的(图1J-P)。雄蕊原基不断生长分化,最终生长为花药和花柱,花药里面分化出花粉,花粉在花粉囊内产生(图1U)。
雌蕊原基分化期:雄蕊原基呈现“向心型”发育,当雄蕊发育到最后一轮的时候,雌蕊原基同时发生分化过程,雌蕊原基主要发生在生长锥的顶部平滑的位置。随着雌蕊原基不断分化,数量越来越多(图1Q-S)。与此同时,大花铁线莲生长锥的体积也在不断缩小。雌蕊原基最终分化为雌蕊,结构为倒生胚珠及其他附属结构(图1V)。
2.2 花芽纵横长度数量变化
在花芽分化的过程中,花芽的形态在发生着变化,同时花芽的体积和数量关系也在发生着变化,花芽在不同的分化阶段横纵长度有所变化(表1)。花芽横长先变短后变长,到雌蕊原基分化期达到了最大值,平均为553 μm,从萼片原基到雄蕊原基增长速度最快,增长了84.8%。花芽的纵长变化规律同横长类似,最快增长速率达100.4%。花芽的横纵长从分化初期到分化结束分别增长了235.2%和248.1%,花芽纵长的增长率要高于花芽横长的增长率。

表1 大花铁线莲花芽分化不同时期横纵长度
2.3 雄配子发育过程
2.3.1 花药壁发育
大花铁线莲的花药具有4个小孢子囊,位于4个角隅处的细胞迅速分裂形成花药雏体(图2A-D),细胞继续分裂逐渐形成形似蝴蝶型的成熟花药(图2E)。最初形成的花药雏体中存在分裂能力较强的细胞,其形态各不相同(图2F)。随着细胞分裂的进行,表皮里侧的细胞分化出孢原细胞,经平周分裂形成初生壁细胞和造孢细胞(图2G、H)。造孢细胞不断分裂分化(图2J、K),为花粉母细胞的形成奠定基础。而初生壁细胞经过分裂则形成2层次生壁细胞(图2L、M)。这2层次生壁细胞从外至内分化成药室内壁,中层和绒毡层多层细胞结构,即花药壁。随着花药发育的进行,细胞体积逐渐变大但药室内壁的储藏物质逐渐变少(图2O-X),后期称为纤维层。中层细胞起初含有丰富的淀粉等物质,具有两层结构(图2O),到花粉母细胞减数分裂的时期,中层细胞被周围挤压,靠里的一层结构被吸收掉(图2P-S),另一层细胞也被挤压而变得扁平但不消失,最终在成熟的花药层细胞中类似于纤维层(图2X)。随着小孢子逐渐分化成为花粉,绒毡层在此过程中渐渐解体(图2T-W)。最终,在花粉成熟阶段,绒毡层仅留一点或完全解体(图2X)。
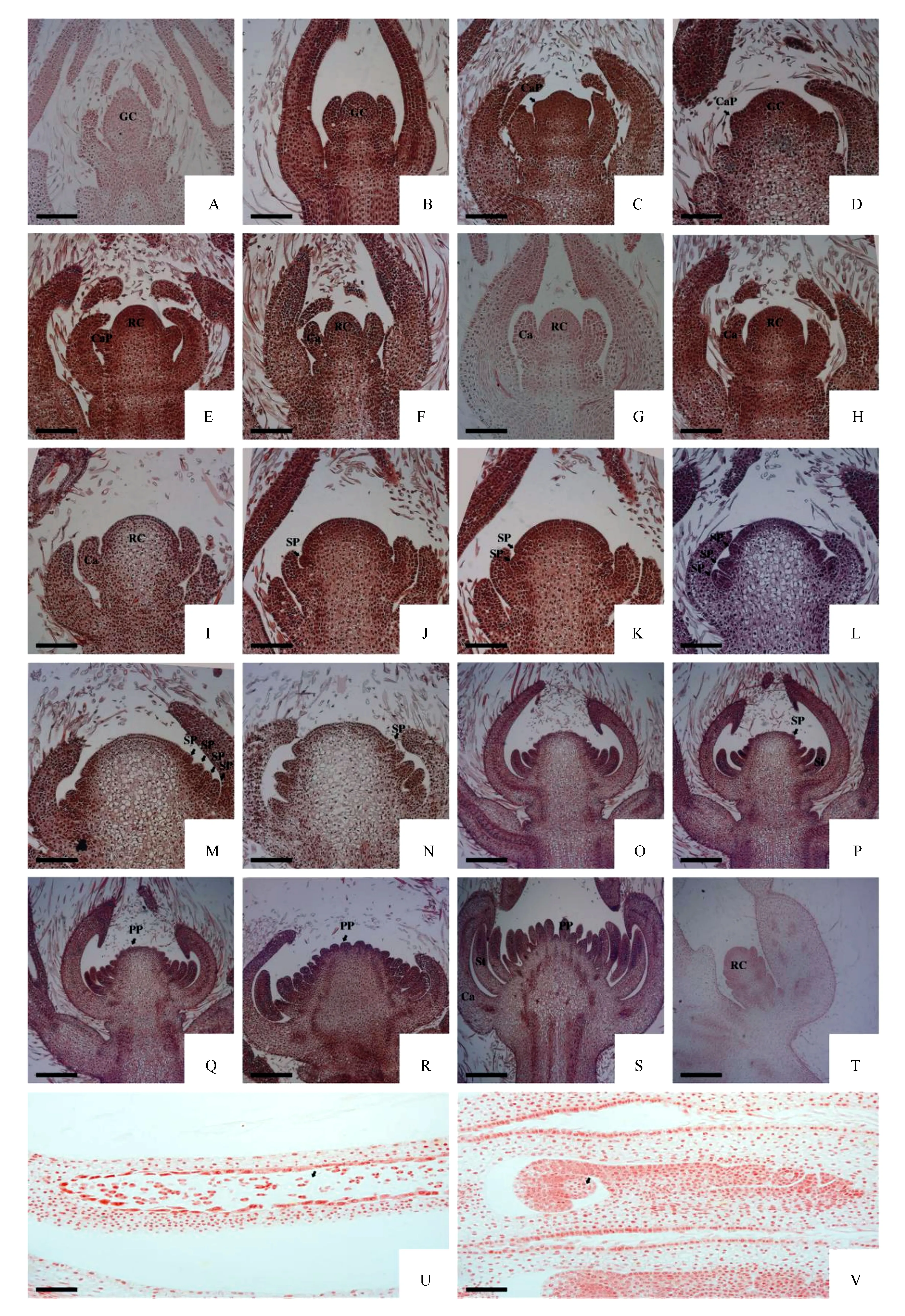
A.营养生长期;B.花芽分化初期;C-E.萼片原基分化期;E-I.生殖生长锥;J.雄蕊原基分化期;K-P.雄蕊原基增多,外层雄蕊逐渐生成;Q-S.雌蕊原基分化期;T.腋生花芽;U.花药结构;V.雌蕊结构;GC.营养生长锥;RC.生殖生长锥;CaP.花萼原基;Ca.花萼;SP.雄蕊原基;PP.雌蕊原基;St.花柱(显微标尺:A-Q,T,U,V=100 μm;R-S=20 μm)。

A-E.花药发育成型过程;F.花药雏体;G-I.孢原细胞平周分裂;J-K.初生造孢细胞分裂;L.初生壁细胞分裂;M.初生壁细胞平周分裂形成两层次生壁细胞;N.次生壁细胞分裂;O.花药壁(小孢子母细胞时);P-V.花药壁(小孢子母细胞减数分裂时);W.花药壁(单核小孢子时);X.花药壁(成熟花粉时);asc.孢原细胞;psc.初生造孢细胞;epi.表皮;end.药室内壁;ml.中层;tap.绒毡层(显微标尺:A-E=50 μm,F-X=20 μm)。
2.3.2 小孢子产生
小孢子母细胞由最初的初生造孢细胞经多次的有丝分裂而得到(图3A),之后不断发育,历经2次减数分裂得到4个细胞核(图3B-I)。而4个细胞核进行胞质分裂形成四面体型或者左右对称型的四分体(图3J-O),即为4个小孢子。最初分化得到的4个小孢子依旧处在胼胝质壁内,且互相隔离。后期在绒毡层解体时会分泌出胼胝质酶,胼胝质壁在酶的作用下而分解(图3P-W)。自此,从中释放出的4小孢子成为了独立状态,并形成4个单核的花粉粒(图3X)。大花铁线莲的减数过程中,在减数第一次分裂完成之后有的能立即产生新的细胞壁,有的不立即产生新的细胞壁,上述两种情况同时出现,因此认为,大花铁线莲胞质分裂时,连续型和同时型2种分裂方式同时存在。

A.小孢子母细胞;B减数I前期细线期;C.减数I前期偶线期;D.减数I前期粗线期;E.减数I前期双线期;F.减数I前期终变期;G.减数I中期;H.减数I后期;I.减数I末期;J.减数Ⅱ前期;K.减数Ⅱ中期;L、M.减数Ⅱ后期;N.四面体型四分体;O.左右对称型四分体;P-U.胼胝质溶解;V、W.释放到药室中的小孢子;X.单核小孢子(显微标尺:A-X=20 μm)。
2.3.3 雄配子体发育
单核花粉粒随着胼胝质的溶解而逐渐呈分散的游离状态(图4A)。刚分散开的单核花粉粒细胞在显微镜下观察可以看到,细胞没有明显的液泡,呈干瘪皱缩状,细胞壁较薄,细胞核此时处于细胞的中心位置,周围有大量的内质网等细胞器。随后,单核花粉粒吸收大量的营养,体积也随之变大,大液泡逐渐在视野中呈现出来,并且将细胞核挤到一旁,此时期称之为单核靠边期(图4B)。单核小孢子继续发育,经有丝分裂变成了二细胞花粉,在花粉粒中存在双核,因有丝分裂过程中胞质分裂不均等,2个子核大小不一,体积较大的为营养核,另外一个是小体积的生殖核(图4C-F)。二细胞花粉粒逐渐成熟,在外形上呈近圆形且可见两层花粉壁,而外壁上能够观察到萌发孔的痕迹(图4G、H)。
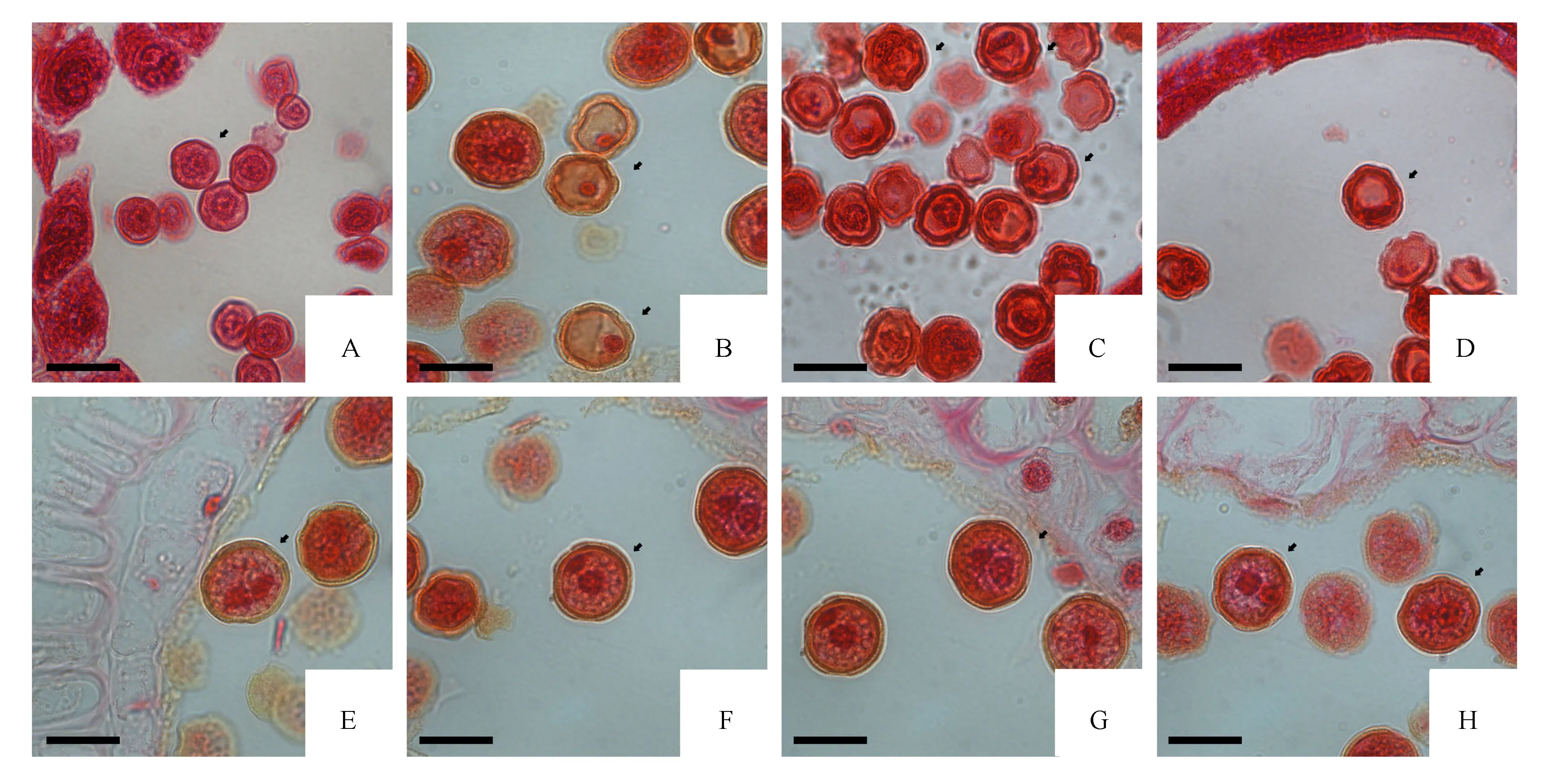
A.单核花粉粒;B.单核靠边期小孢子;C-E.营养细胞与生殖细胞的产生过程;F.初期形成的二细胞花粉粒;G、H.逐渐成熟的二细胞花粉(显微标尺:A-H=20 μm)。
经试验发现,大花铁线莲花药壁的发育过程和小孢子及雄配子的发育过程在发育时间上存在一定的对应关系,当花药壁完成最后的发育过程时,二细胞花粉也发育成熟(表2)。

表2 小孢子及雄配子发育阶段对应的花药壁发育阶段
2.4 雌配子发育过程
2.4.1 大孢子发生
在子房的胎座上着生着胚珠(图5I),胚珠原基(图5A、B)就是由大花铁线莲子房腹缝线(图5H)内表皮下的一些细胞经过不断地平周分裂而产生的。原基尾端细胞经发育而成了珠柄,与胎座连在一起,另一端形成突起,即为珠心。珠心尾部有一群细胞分裂旺盛,经不断分裂不断延伸把珠心包围起来而形成珠被。发育完善的胚珠为单室、单珠被和单个倒生的胚珠(图5C-E)。但是也有一个胎座上存在两个胚珠的情况,且两个胚珠发育进程存在差异(图5F、G)。随着珠心和珠被的继续发育,靠近珠孔一端的珠心表皮下细胞逐渐分化出了孢原细胞(图5J),经平周分裂,形成二分体。接着分裂生成1个线型的大孢子四分体(图5K-N)。四分体中,3个周缘细胞接近珠孔端,后期逐渐地退化消失完全,而远离珠孔端的的造孢细胞继续发育最终形成单核胚囊(图5K-T)。
2.4.2 雌配子体发育
单核胚囊在发育过程中吸收很多营养物质,所以体积逐渐增大,在整个珠心组织中占很大的空间位置(图6A)。随后发生一次有丝分裂形成2个子核,多数横向排列,鲜有纵向排列(图6B、C)。不久,2个子核向胚囊的两极移动,生成二核胚囊(图6D、E),又发生一次有丝分裂形成了四核胚囊(图6J),但是有时两端的子核分裂可能存在不同步现象或者其他状况,胚囊内可见3个核(图6H、I)。四核胚囊的加倍分裂,最终胚囊拥有8个细胞核。8个细胞核的去向不一,其中,胚囊两端的2个细胞核向中间靠拢,发育成含有2个极核的1个中央细胞。珠孔端的3个核最终分化成为1个较大的卵细胞和2个助细胞。合点端的3个核分化成3个反足细胞。最终,发育成为8核7细胞的成熟胚囊(图6X)。
大花铁线莲的卵细胞多数位于珠孔端(图6M、N),鲜见位于合点端(图6O)。已经成熟的卵细胞中细胞器的数量会减少,珠孔端的大液泡散开成为小液泡,细胞核会更加贴近合点端(图6K)。
2个助细胞紧紧相邻,与卵细胞呈三角鼎力之势(图6L)。助细胞代谢旺盛,且丰富的细胞器使得细胞质染色较深(图6P)。助细胞在珠孔端的细胞壁向细胞腔内分生成丝状器(图6Q)。
中央细胞的核是极核,有纵向和横向两种排列状态(图6R-S),中央细胞表现为一定的极性,常向珠孔端靠近(图6T-U),有时也靠向反足细胞(图6V)。
3个反足细胞在胚囊内一般呈三角形排列(图6W)。
3 结论与讨论
通过对大花铁线莲胚胎学的初步研究,可以发现,大花铁线莲花芽分化分为营养生长期、花芽分化初期、萼片原基分化期、雄蕊原基分化期、雌蕊原基分化期,共5个阶段。当植物的芽顶端形状改变或体积变大时,认为这是花芽分化开始的迹象。大花铁线莲在花芽分化过程中,生长锥由尖锐逐渐平坦,这与袁德义[11]等的观点十分相符。
大花铁线莲花药4室,花药壁由表皮、药室内壁、1~2层中层和1层绒毡层构成。绒毡层类型为腺质绒毡层。这与毛茛科的侧金盏[12]、金莲花和驴蹄草的观测结果相同[13]。一般认为,绒毡层细胞消失的过程就是为小孢子发育提供营养的过程[14]。大花铁线莲绒毡层是由内层的次生壁细胞发育而成,当花粉母细胞将近完成减数分裂时,绒毡层细胞开始逐渐解体,最终,花粉成熟时绒毡层完全消失或仅存一点残留。由此可以证明,绒毡层的状态满足小孢子发育过程中对营养物质的需求。
大花铁线莲的小孢子母细胞孢质分裂方式为连续型和同时型均存在,形成左右对称型和四面体型的四分体,成熟花粉为2-细胞型。Heslop-Harrison[15]认为,胞间连丝的存在令同一药室内的小孢子母细胞同时进行减数分裂。而大花铁线莲最初的小孢子母细胞多呈不规则型且存在胞间连丝,在后期的减数分裂过程中没有发现异常,较为同步,这与上述观点吻合。
大花铁线莲子房为单室,单胚珠、单珠被、倒生胚珠。大孢子母细胞是由子房腹缝线内表皮下的一些细胞分化得到的孢原细胞,经平周分裂发育成造孢细胞,造孢细胞进一步分化而得到的。这和Tamura[16]所描述的毛茛科植物大孢子母细胞的由来十分接近。大孢子母细胞经减数分裂生成直线型四分体,胚囊为蓼型发育,含有7细胞8核。这些表现与已报道的铁线莲的胚胎学特征颇为相似[17-18]。
大花铁线莲的一朵花上会有多株雌蕊,各个雌蕊之间的发育存在着差异。此外,雌蕊和雄蕊的发育表现不同步,存在雌雄异熟的现象,具体表现为雄蕊发育比雌蕊更早。这在核桃[19]、板栗[20]等植物中也出现过类似情况。植物的这种雌雄发育不同步的现象能有效延长授粉期[21]。
文中对东北地区大花铁线莲的花芽分化和花器官发育特性进行了较为详细的研究,发现大花铁线莲在生长过程中,花的外部形态特征和内部的配子体发育有相应的对照关系。通过对外部形态特征的观察,可以更好地掌握其内部雌雄蕊的发育进程,以便预测花期、人工授粉加强管理等。本试验使铁线莲属植物的雌雄配子体发育和大小孢子相关的胚胎学研究更加丰富,也为大花铁线莲花粉、胚珠组织培养、遗传育种及分子生物学提供可靠的理论依据。
