塞罕坝地区植被景观格局时空尺度效应1)
2021-12-03徐雯雯董雪婷张志东黄选瑞
徐雯雯 董雪婷 张志东 黄选瑞
(河北省林木种质资源与森林保护重点实验室(河北农业大学),保定,071000)
景观格局是由形状大小不同的景观斑块在景观空间上的排列,反映了景观的异质性,是诸多自然和人为因素在不同时空尺度上作用的结果,是景观生态学的研究重点之一[1-3]。植被资源作为陆地生态系统的重要组成部分之一,为人类社会提供重要的生态和经济服务价值。然而,气候变化和人为干扰极易破坏植被景观格局的整体性,使其破碎化加剧,导致生物多样性减少和生态系统服务价值降低[4-5]。
有关研究表明,景观异质性的量化分析与分析尺度密切相关,尺度选择直接关系到景观格局分析结果的可靠性[6-7],分析不同时空尺度下的植被景观格局演变特征,对于合理分析植被景观生态过程、有效监测环境及生物多样性变化具有重要的理论和现实意义[8]。在遥感及地理信息系统技术的基础上,采用景观格局指数法能够从斑块水平、类型水平和景观水平上量化景观格局信息[9-10]。然而,不同类型的景观指数对尺度变化的响应存在差异[11],按照景观指数的粒度效应可将其分为可预测响应型、阶梯变化型、波动变化型及无规律变化型[12]。基于空间粒度效应的景观格局研究成为目前的研究热点[13-14]。生态学中的空间尺度通常指空间幅度或粒度,对于空间数据或影像资料而言,空间粒度是指最大分辨率或像元大小[15]。空间粒度大小变化导致用于量化景观格局信息的指数随之发生变化,即景观指数在不同时空尺度下呈现不同的响应状态[12],辨识特征空间粒度是真实准确反映植被景观格局与生态过程自身运行特征的关键[16]。景观格局指数本身也存在一些明显的局限性,由于空间尺度、时间尺度、景观类型等不同,而呈现出不同敏感性和可预测性[17-18]。因此,通过多个指数的联合应用来量化景观格局演变与生态过程间的关系,能够有效的解释植被景观生态过程。
塞罕坝地区处于华北地区典型森林草原交错带,集森林、草原、草甸、沼泽和水体为一体的复杂多样的生态系统,能够提供极其重要的生态服务功能。然而,受国家政策和人类活动的影响[19],塞罕坝地区人工林面积持续增加,灌木林面积迅速减少,天然次生林及草地面积先减后增,导致该地区的景观破碎化程度加深,物种丰富度急剧下降[20]。分析各植被景观对时间和空间粒度变化的响应情况,能够更好地理解植被资源动态变化与生物多样性间的关系提供依据[21]。本文针对塞罕坝地区植被景观格局变化显著的特点,以3期遥感影像以及样地调查数据为基础,基于信息熵模型和景观指数方法,探讨塞罕坝地区4大植被类型(草地、天然次生林、灌木及人工林)的景观格局时空尺度效应,具体包括:在时空尺度下,4种植被景观格局尺度效应;景观指数在不同时空尺度下的可预测性;最大程度的保留景观格局信息量的空间粒度阈值。通过对景观格局时空尺度效应研究,为更科学合理的森林经营和监测以及优化植被景观分布格局提供科学依据。
1 研究区概况
塞罕坝地区位于河北省最北部,地处内蒙古高原与冀北山地交界处(42°4′~42°36′N,116°53′~117°39′E),海拔高度1 010~1 939.6 m。属于寒温带半干旱半湿润大陆性季风气候,年平均气温-1.4 ℃,极端最高和最低气温分别为30.9 ℃和-43.2 ℃;年降水量490 mm。土壤类型为坝上地区以风沙土为主,伴随有草甸土和沼泽土,接坝地区以棕壤和灰色森林土为主[22]。乔木树种主要有华北落叶松(Larixprincipis-rupprechtii)、樟子松(Pinussylvestrisvar.mongolica)、云杉(Piceaasperata)、白桦(Betulaplatyphylla)、蒙古栎(Quercusmongolica)等;灌木主要有山刺玫(Rosadavurica)、沙棘(Hippophaerhamnoides)、山杏(Armeniacasibirica)、胡枝子(Lespedezabicolor)等;草本植物主要有地榆(Sanguisorbaofficinalis)、金莲花(Trolliuschinensis)、老芒麦(Elymussibiricus)、披针叶苔草(Carexlancifolia)等。
2 研究方法
2.1 数据来源与处理
本研究选取塞罕坝地区1989、2000、2016年每年春、夏、秋、冬四个季节的Landsat TM/ETM+/OLI卫星遥感数据(空间分辨率30 m)作为基础数据,以2002年的SPOT5卫星影像(分辨率2.5 m)、2012年航空影像(分辨率0.5 m)、DEM高程数据(分辨率30 m)以及当地林场二类调查矢量数据为辅,进行图像预处理和训练样区选取。参考国际地圈生物圈计划(IGBP)中的土地利用类型分类标准,结合塞罕坝地区生态环境特征及研究需要,将土地利用类型分为7类:草地、天然次生林、灌木、人工林、城镇及建设用地、水体及裸地。
遥感影像处理过程主要包括预处理、训练样区选取、分类和分类后处理4个步骤。在ENVI5.1软件中进行辐射定标、大气校正、几何精校正、影像镶嵌及裁剪;采用具有较高分类精度的随机森林分类算法[23-24],在ENVI5.1软件中结合enMAP-BOX工具包进行遥感影像分类,利用基于影像原始波段、纹理特征数据、地形数据以及植被指数选取的287个分类特征变量构建随机森林分类模型;采用ENVI5.1中Majority/Minority分析进行分类后处理,随机选取30%验证样本进行精度评价,结果表明3期影像分类整体精度和Kappa系数均超过90%,能够满足研究需要[25](见图1)。

图1 1989、2000与2016年塞罕坝地区土地利用类型图
2.2 空间粒度变换与景观指数选取
粒度效应研究是把精微尺度上的观察、试验和模拟结果外推到较大尺度,即把景观类型栅格图进行尺度上推(粗粒化)[26-27],利用ArcGIS10.2软件中的空间分析模块,运行Resample命令将研究区的土地利用类型图通过栅格聚合的方式转换为不同粒度的栅格数据。本文空间粒度变换范围从30 m×30 m到300 m×300 m之间,以30 m为间隔,最终生成空间分辨率分别为30、60、90、120、150、180、210、240、270、300 m共10幅不同粒度的景观类型栅格图。目前描述景观格局的指数很多,参考国内外类似研究成果中的指数选择以及研究区的景观特征[28-29],本文选取了6个类型水平指数以及3个景观水平指数(见表1),在Fragstats4.2软件中计算景观指数。

表1 选取的景观指数
2.3 基于信息熵的最优尺度选择
景观格局对尺度的依赖性主要体现在不同景观有不同的格局特征,且对尺度变化的响应也不同,若尺度过小则造成景观格局信息量过大而掩盖部分重要信息,相反则造成缺失细节信息[30]。因此针对特定景观需要选择适宜分析粒度,即最佳粒度,便于对景观格局进行准确有效地分析。信息熵作为信息论中用于衡量信息量的指标已被广泛应用到量化景观格局的研究[31-32],它描述的是信源的不确定性,是信源中所有目标的平均信息量,表示为随机变量的概率分布函数。利用信息熵对不同粒度下的植被景观格局信息进行衡量,其基本定义为:
式中:H表示信息熵值;n是所有事件数量;Pi表示随机事件i的概率。
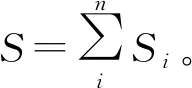
3 结果与分析
3.1 时间尺度效应
在时间尺度上,通过判别随时间推移3个时期的景观指数响应曲线变化趋势是否具有一致性,可将指数分为两类:类型Ⅰ,景观指数响应曲线变化趋势具有一致性,可预测性强(见图2、图3);类型Ⅱ,景观指数响应曲线变化趋势不一致,可预测性较弱(见图4)。
由图2、图3可知,类型Ⅰ包括类型水平上边缘密度、斑块密度、景观形状指数、聚集度指数以及景观水平上蔓延度指数、香浓多样性指数、香浓均匀度指数,以上指数的粒度效应曲线在3个时期中的变化趋势具有一致性。边缘密度和斑块密度随时间推移在4种植被景观上的变化趋势具有可预测性,但在4种植被类型间的变化趋势不同;边缘密度和斑块密度在草地上呈均先增后降的“n”字型变化趋势,在天然次生林上均呈先降后增的“v”字型变化趋势,在灌木上呈稳定下降趋势;人工林的边缘密度不断增加,斑块密度逐渐降低。聚集度在草地及灌木中呈现相似的变化趋势,在2016年出现最低值,在天然次生林和人工林上呈增长趋势。形状指数在草地上不断增加,在天然次生林、灌木及人工林上总体呈降低趋势。蔓延度指数可以反映景观中不同类型斑块间的团聚或延展趋势,其值越大表明景观中有连通度较高的优势斑块类型存在。研究期间,研究区的蔓延度指数逐渐增加,表明研究区景观斑块之间的连通性增强。香浓多样性指数及香浓均匀度指数可以评价景观类型的多样性及景观类型分布均匀度,二者随时间推移呈降低趋势,表明研究区的景观多样性及斑块分布均匀度降低。因此,在时间尺度上,研究区人工林的聚集程度增强,破碎化程度降低,整体上,天然次生林和草地的破碎化程度增加,灌木破碎度有所下降。
由图4可知,类型Ⅱ中包括最大斑块指数、面积加权平均形状指数的粒度效应曲线随时间推移其变化趋势不一致。最大斑块指数值的大小可以用来确定景观中的优势斑块类型,随时间推移,其值在草地及灌木上呈下降趋势,且波动幅度趋于平缓,在天然次生林及人工林上呈上升趋势,且增幅显著。面积加权的平均形状指数能够度量斑块形状的复杂程度,其值在草地及灌木上呈逐渐降低趋势,表明其斑块形状趋于规则简单,天然次生林及人工林大体上均呈增加趋势,表明其斑块形状复杂性增强。
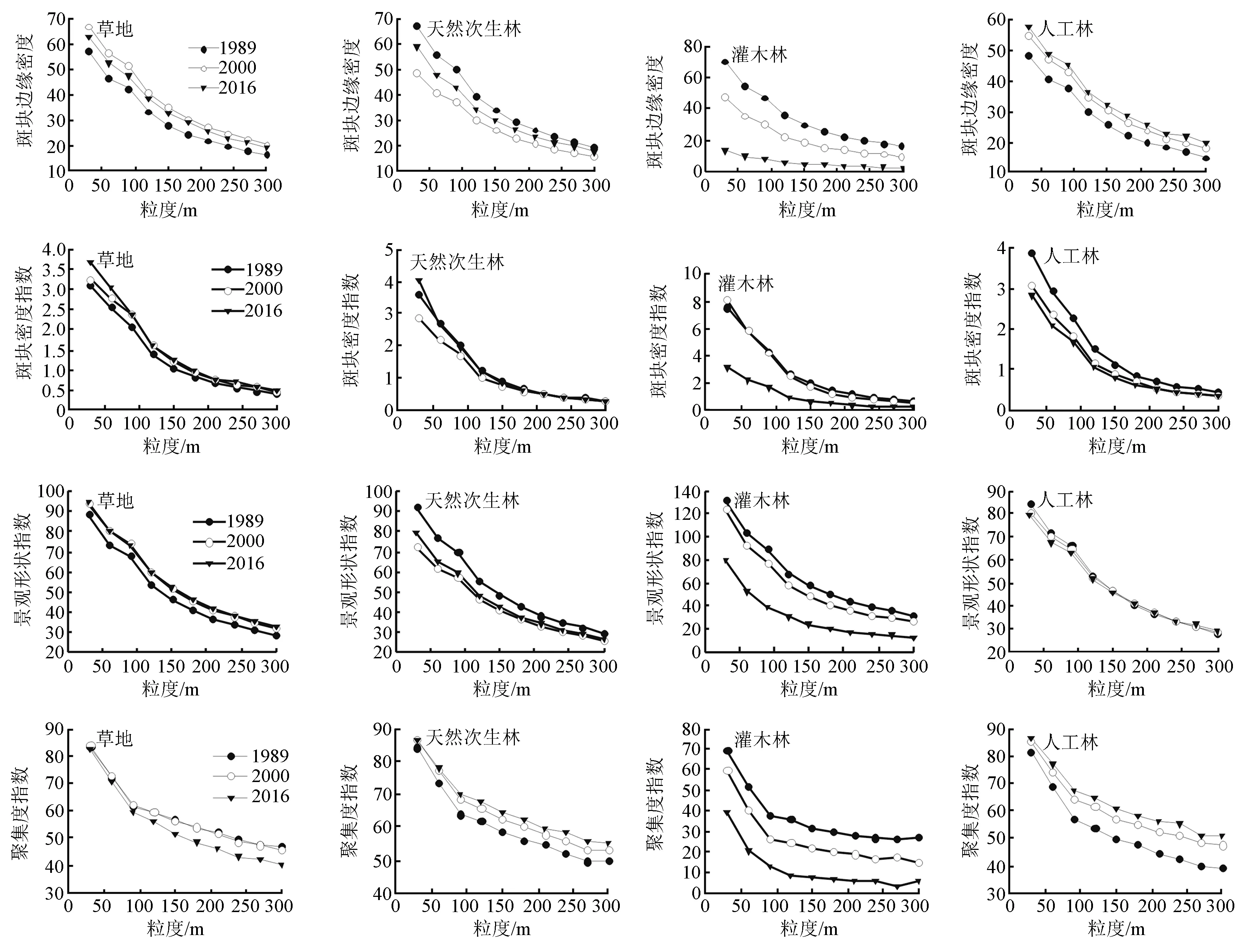
图2 研究区4种植被景观类型水平指数多尺度响应(类型Ⅰ)
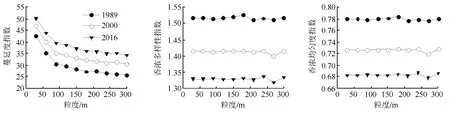
图3 研究区4种植被景观景观水平指数多尺度响应(类型Ⅰ)

图4 研究区4种植被景观类型水平指数多尺度响应(类型Ⅱ)
3.2 空间尺度效应
在空间尺度上,根据各景观格局指数随空间粒度增加的变化趋势,将景观指数分为三类:第1类指数随空间粒度变化呈单调下降的趋势,可预测性强(见图2、图3)。第2类指数随空间粒度变化呈波动变化趋势,变化趋势不规则,可预测性较差(见图4)。第3类指数基本不随空间粒度增加而变化,不具有可预测性(见图3)。
由图2、图3可知,第1类景观指数包括类型水平指数(边缘密度、斑块密度、景观形状指数、聚集度指数和景观水平指数(蔓延度指数)。随粒度增加,这类景观指数在4种植被景观上均呈单调下降,具有明显的转折点,且第一尺度域为30~90 m。在第一尺度域内,景观格局具有粒度效应的特征信息,考虑到计算景观格局特征信息的有效性以及计算效率,需在第一尺度域内选择中等偏大的粒度[15,33],即最佳空间粒度为90 m。
由图4可知,第2类景观指数包括最大斑块指数及面积加权的平均形状指数。最大斑块指数在天然次生林、灌木及人工林上呈波动增加趋势,面积加权的平均形状指数在4种景观类型中呈波动下降趋势,这类景观指数随粒度增加的波动趋势不规则,可预测性较差,但在30~90 m区间上变化趋势相对规则,因此最佳空间粒度为90 m。
由图3可知,第3类景观指数为景观水平上香浓多样性指数以及香浓均匀度指数。它们在空间粒度上变化趋势相似,在小尺度上变化不明显,在大尺度上有了较小幅度的变化。同时,本文通过选择第1类景观指数进行数学模型拟合来进一步验证景观指数对空间粒度变化响应的有效性,并发现其拟合度均较为理想(见表2)。

表2 景观指数曲线拟合表
3.3 时空尺度效应
基于上文对景观指数时空尺度效应的分析,将分析结果汇总可知(表3),在时空双重尺度下,均具有可预测性的指数为边缘密度、斑块密度、形状指数、聚集度指数及蔓延度指数。
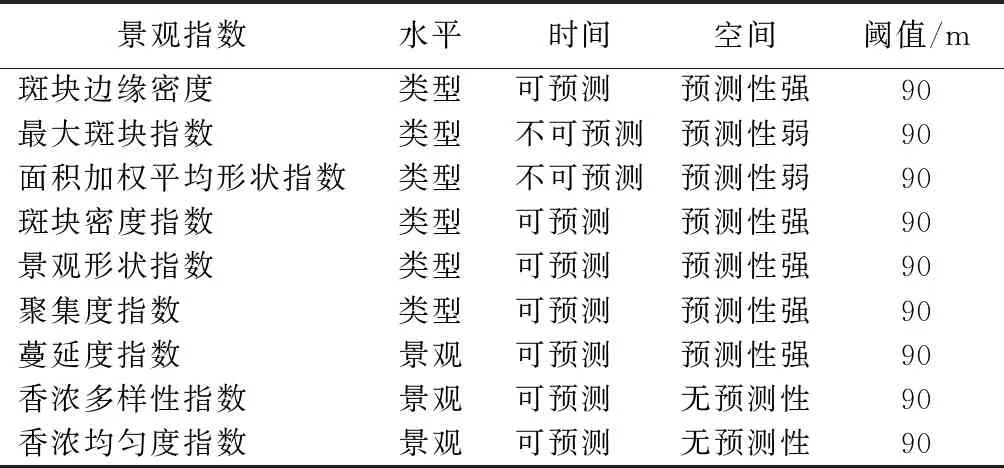
表3 景观格局指数时空尺度效应以及空间相关性
3.4 基于信息熵景观格局最优尺度选择
由图5可知,以2016年植被格局变化为例来探讨信息熵对空间尺度的响应,信息熵值在60~90 m区间上开始剧烈上升,粒度增至90 m后又急速下降,表明一些重要景观格局特征信息开始丢失,因此,本文认为60~90 m为能够最大程度的保存信息丰富度的阈值区间。
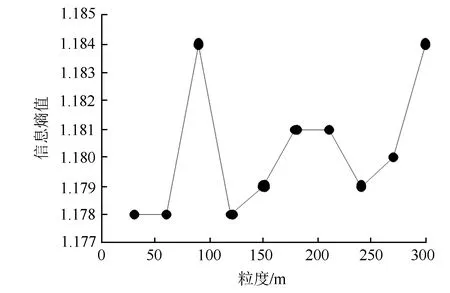
图5 不同空间粒度景观格局信息熵值的变化
4 讨论
自然因素与人为活动共同影响着景观格局[34],塞罕坝地区拥有北半球最大人工林林场,能够提供的生态服务价值不容忽视[35],1989—2016年塞罕坝地区植被资源在人为活动及自然因素共同影响下发生了显著变化[19]。通过分析塞罕坝地区植被景观格局时空尺度效应发现,量化景观格局特征的景观指数具有尺度依赖性,且各景观指数在时空尺度上的可预测性不同,这与研究区的生态过程以及人类干扰有较大关系[36]。
在时空尺度下,均具有较强可预测性的指数包括边缘密度、斑块密度、形状指数、聚集度指数及蔓延度指数。
在时间尺度上,能够表示草地破碎度的指数(如斑块密度、形状指数、聚集度指数)均增加,表明研究区草地破碎化随时间推移加剧,这与张宏斌等[37]对呼伦贝尔草地景观格局时空演变格局变化的研究结果相似,原因是草地受到与人类干扰相关的造林、旅游开发等活动的影响导致破碎化加剧。天然次生林的斑块密度增加,而边缘密度及形状指数减少,说明塞罕坝地区天然次生林的破碎化程度在研究的期间内不断加剧,原因是人工林入侵,导致其整体性被打破;人工林的斑块密度及形状指数不断减少,边缘密度及聚集度增加,这是由于近27 a人工林大面积增加使其破碎度降低[19-20]。
在空间尺度方面,边缘密度、斑块密度、形状指数、聚集度指数以及蔓延度指数在4种植被景观中均随着粒度增加而减少。这是由于在特定斑块类型下,随着粒度增加小型斑块将被合并为大型斑块,分散的小型斑块在合并过程中甚至消失[14],这会造成斑块间边界消失、斑块数量及多样性减少,同时斑块形状趋于简单规则。聚集度指数及蔓延度指数可以反映景观类型内部的团聚程度及连通性,其值越小,表明景观由许多离散的小斑块组成,相反,其值越大表明景观由连通度较高的大斑块组成[38],二者随粒度增加而降低,说明由于斑块不断融合,斑块间的相邻距离增大,空间分布更为分散,破碎化加剧[13]。通过分析景观指数对景观格局时空变化的预测情况,发现研究区具有明显的粒度阈值,重要的临界域粒度为90 m,当粒度大于90 m时,景观格局信息量将会迅速丢失。因此,综合考虑景观格局粒度效应以及信息熵结果,本文认为90 m为分析研究区植被景观格局的最佳空间粒度。
植被景观格局的变化具有一定的生态效应,如引起生物多样性、栖息地可用性及土壤侵蚀等变化[39]。塞罕坝地区植被资源结构单一,以人工林为主导,且天然次生林、草地破碎化程度高,尤其是空间粒度大于90 m时,景观格局发生显著变化,这将对当地物种栖息地的选择与保护产生负面影响[40],如对栖息地范围波动敏感的鸟类造成威胁[41]。为此,应采取积极措施减少人为干扰,应做好“封山禁牧”,以最大程度减少人为活动对草地的干扰,通过“封山育林”对天然次生林进行恢复和保护,加速天然次生林的形成;在外围建立缓冲区以减少人类活动对核心区天然次生林的干扰,在栖息地之间建立生态廊道,加强生态斑块间的连通性并促进物种间的交流和辐射,对于保护当地生物多样性具有重要意义[14,40]。
尺度效应是景观格局研究的基本内容,景观类型分类标准、遥感影像分辨率大小、空间粒度推绎范围、推绎步长、空间数据聚合方法及景观指数自身算法的差异等都是致使景观指数粒度效应不同的原因,本文探讨了不同景观指数的时空尺度效应,对指数选取、空间粒度推绎及格局分析具有参考价值。
5 结论
1989—2016年间,塞罕坝地区人工林大面积增长导致景观格局向非均衡化方向发展,人工林景观破碎化有所好转,但草地和天然次生林等天然植被景观格局破碎化程度较高。塞罕坝地区4种植被景观的空间尺度效应相似,景观指数表现为随粒度增加而下降,而4种植被景观的时间尺度效应不同,随时间推移人工林及灌木林破碎化程度降低,草地及天然次生林破碎化增加;研究选取的景观指数在时空尺度上表现出不同的可预测性,在时空尺度上均具有较高尺度效应的指数包括边缘密度、斑块密度、景观形状指数及聚集度指数;研究区植被景观格局的最佳分析粒度为90 m。
