浙江省主栽水稻品种抗稻瘟病基因的分子检测
2021-12-02沈浙南邱结华解军辉时焕斌寇艳君
沈浙南 邱结华 解军辉 时焕斌 寇艳君*
(1 中国水稻研究所/水稻生物学国家重点实验室,杭州311400;2 三峡大学生命科学院,武汉430000;#共同第一作者;*通讯作者:kouyanjun@caas.cn)
稻瘟病是由稻瘟病菌(Magnaporthe oryzae/Piricularia oryzae)引起的真菌性病害,世界各稻区均有发生,一般导致水稻减产10%~30%,严重的甚至颗粒无收[1]。田间稻瘟病菌菌群毒力的多变性导致生产上推广使用的抗病品种常因发病严重而淘汰[2]。目前生产上对稻瘟病的防治主要有化学防治和抗病育种,培育抗病新品种是最绿色经济的方法[3]。
近年来,通过图位克隆等手段在水稻中鉴定了100 多个抗稻瘟病基因,其中36 个已被克隆[4]。这些水稻抗稻瘟病基因分布在除第3 染色体外的所有染色体上,在第6、11 和12 染色体上存在较多的抗稻瘟病基因簇。目前,大部分克隆的抗稻瘟病基因均有特异的分子检测标记[5]。这些分子检测标记的开发为水稻抗病分子育种奠定了良好基础。育种家结合分子检测标记可以快速鉴定水稻品种抗稻瘟病基因型,有效辅助抗病新品种的选育。黄乾龙等[6]利用 Pi2、Pi5、Pi9、Pikh、Pita、Pikm 和Pib 等7 个抗稻瘟病基因的分子标记对重庆73 个水稻骨干品种的抗病基因进行鉴定,解析这7 个抗病基因在水稻抗稻瘟病育种中的作用。陆展华等[7]利用 Pi1、Pi2、Pi9、Pib 和 Pita 等 5 个主效抗病基因的分子标记,结合叶瘟和穗瘟的抗性鉴定,对广东省70 个主栽品种抗稻瘟病基因的组成进行了解析。张文龙等[8]通过分子标记分析了云南省地方水稻品种稻瘟病抗性与抗稻瘟基因型的相关性,为云南省抗稻瘟病品种的选育提供了重要参考。这些研究表明,分子检测标记正逐步被育种家使用,为稻瘟病抗性育种提供了重要的辅助工具。
本研究在人工接种稻瘟病抗性鉴定的基础上,利用 Pi1、Pi5、Pi2、Pi9、Pia、Pizt、Pigm、Pib、Pik 和 Pikh 等 7个主效抗稻瘟病基因分子检测标记,对63 个浙江省主栽水稻品种的稻瘟病抗性进行综合分析,旨在明确浙江主栽品种的抗病基因型,并初步探讨了稻瘟病抗病基因的基因型与稻瘟病抗性之间的相关性,为浙江省水稻抗稻瘟病品种的选育提供参考。
1 材料与方法
1.1 试验材料
选取浙江省63 个水稻主栽品种为材料,其中,籼稻类型品种24 个、粳稻类型30 个、籼粳交类型9 个(表1),由中国水稻研究所黄世文研究员提供。感病对照品种为丽江新团黑谷和CO39,丽江新团黑谷由中国农业大学彭友良教授提供,CO39 为本实验室保存的品种。
1.2 稻瘟病菌的培养
在无菌条件下,将浙江省稻区分离的9 个保存菌株(由浙江省农业科学院提供)[9]接种至PA 培养基(酵母提取物1 g/L,乳糖2.5 g/L,蔗糖2.5 g/L,酸梅汁40 mL/L,琼脂粉20 g/L,调pH 至6.5),置于28℃恒温培养箱内活化3 d 左右。活化完成后接种到燕麦培养基(燕麦40 g/L,番茄汁150 mL/L,琼脂粉 20 g/L)。在 28℃的恒温培养箱内培养5 d。用灭菌水去除表面的菌丝,放入光照培养箱内产孢2 d 后,用含0.2% Tween 20 的蒸馏水洗孢子,配制成5×105个孢子/mL 的孢子悬浮液用于喷雾接种和注射接种。
1.3 品种稻瘟病抗性鉴定
1.3.1 叶瘟的鉴定
参试水稻品种种子用抗菌素402 浸种2 d 后,放入28℃的恒温培养箱中进行催芽,完成催芽后播种于育秧盘内,每个品种播种5~10 粒。待秧苗长至4 叶期时进行喷雾接种,接种后用PVC 膜密封保持湿度,黑暗培养24 h 后恢复光照,置于22℃条件下培养7 d 后调查发病情况。按国际水稻研究所(IRRI)0~9 级的分级标准(0 级,无病;1 级,抗;3 级,中抗;5 级,中感;7级,感;9 级,高感)记载叶瘟。
1.3.2 穗颈瘟的鉴定
参考李湘民等穗瘟接种方法[10],用医用注射器将0.5 mL 孢子悬浮液注入稻苞内,每穗接1 次。接种20 d后开始调查穗瘟发病情况,按国际水稻研究所0~9 级的分级标准(标准同叶瘟)记载穗瘟。
1.4 分子标记检测
CTAB 法提取水稻品种DNA:首先在2.0 mL 的离心管中,加入剪碎的水稻叶片,加入800 μL 的CTAB提取液,加入钢珠,放入植物组织破碎仪中60 Hz 研磨60 s,65℃水浴 30 min,每隔 10 min 上下颠倒混合,65℃水浴后取出室温冷却,加入等体积的氯仿,12 000 rpm离心10 min,吸取700 μL 的上清液于新的1.5 mL 的离心管中,加入2 倍体积的无水乙醇,-20℃放置30 min,12 000 rpm 离心10 min,去上清,倒置于吸水纸上,室温干燥过夜,加150 μL 去离子水,4℃保存备用。
PCR 扩增:抗稻瘟病基因分子选择标记引物见表1,引物由杭州有康生物公司合成,PCR 反应体系采用20 μL,其中 2×Mix 10 μL、F/R 引物各 1 μL、DNA 模板1 μL,加水 7.5 μL 补齐。PCR 程序为 95℃/3 min 预变性,94℃/30 s、55℃/45 s、72℃/30 s,35 个循环,后 72℃条件下5 min。
电泳检测:PCR 扩增产物采用3%的琼脂糖凝胶电泳,180 V 电泳45 min,伯乐凝胶成像仪拍照记录,基因型条带信息记录在Excel 中。
1.5 统计分析
采用SAS 9.2 和Excel 2016 进行统计分析,相关性分析采用SAS 程序corr,遗传聚类采用SAS 程序cluster,采用类平均法得到聚类树。
2 结果与分析
2.1 水稻品种对稻瘟病的抗性分析
本研究通过室内苗期和田间穗期人工接种,对浙江省63 个主栽品种进行稻瘟病抗性分析。结果(表2)显示,抗叶瘟品种42 个(占比66.7%)、感叶瘟品种21个,其中,高抗品种12 个、抗病品种28 个及中抗品种2 个;抗穗瘟病品种32 个(占比50.8%)、感病品种31个,其中高抗品种4 个、抗病品种13 个及中抗品种15个。这些结果显示,浙江省主栽品种整体对叶瘟抗性良好,但穗瘟抗性有待进一步加强。
2.2 水稻品种的抗稻瘟基因型鉴定
利用表1 中的特异引物对63 个水稻品种进行分子检测,结果表明,参试的63 个水稻品种中,检测到6个抗病基因的品种2 个,检测到5 个抗病基因的品种12 个,检测到4 个抗病基因的品种20 个,检测到3 个抗病基因的品种18 个,只含有1 个抗病基因的品种1个,此外,有2 个品种未检测到本研究中的抗病基因(表2)。在63 个水稻品种中,分布频率最高的基因是Pizt 和Pib,有45 个水稻品种鉴定到Pizt 和Pib 基因,有43 个水稻品种鉴定到Pi2 和Pi5 基因,鉴定到Pikh、Pia、Pi1 及 Pik 的水稻品种分别有 41、37、32 和 30 个,分布频率最低的基因是Pigm 和Pi9,均只有2 个品种鉴定到。这些鉴定结果说明,浙江省主栽品种中Pizt、Pib、Pikh、Pia、Pi1 和 Pik 抗病基因利用频率高,而 Pigm和Pi9 利用频率低。

表1 本研究所用到的引物信息

表2 63 个水稻品种抗稻瘟病基因检测结果

续表2
2.3 抗稻瘟病基因间的相关性分析
对浙江省63 个主栽品种抗性表型和10 个抗稻瘟病基因之间的相关性分析显示,品种的叶瘟抗性和穗瘟抗性之间相关系数为0.255,达到显著水平(P=0.046)(表3)。叶瘟抗性与Pi1 和Pib 基因之间的相关性系数分别为0.302 和0.610,达到显著水平。穗瘟抗性与Pi9、Pi1 及Pia 基因之间相关性达到显著水平,穗瘟抗性与Pib 基因相关系数为0.493,达到极显著水平(P<0.001)。结果说明浙江省主栽品种中Pi1 和Pib 基因在叶瘟和穗瘟抗性过程中起着重要作用。
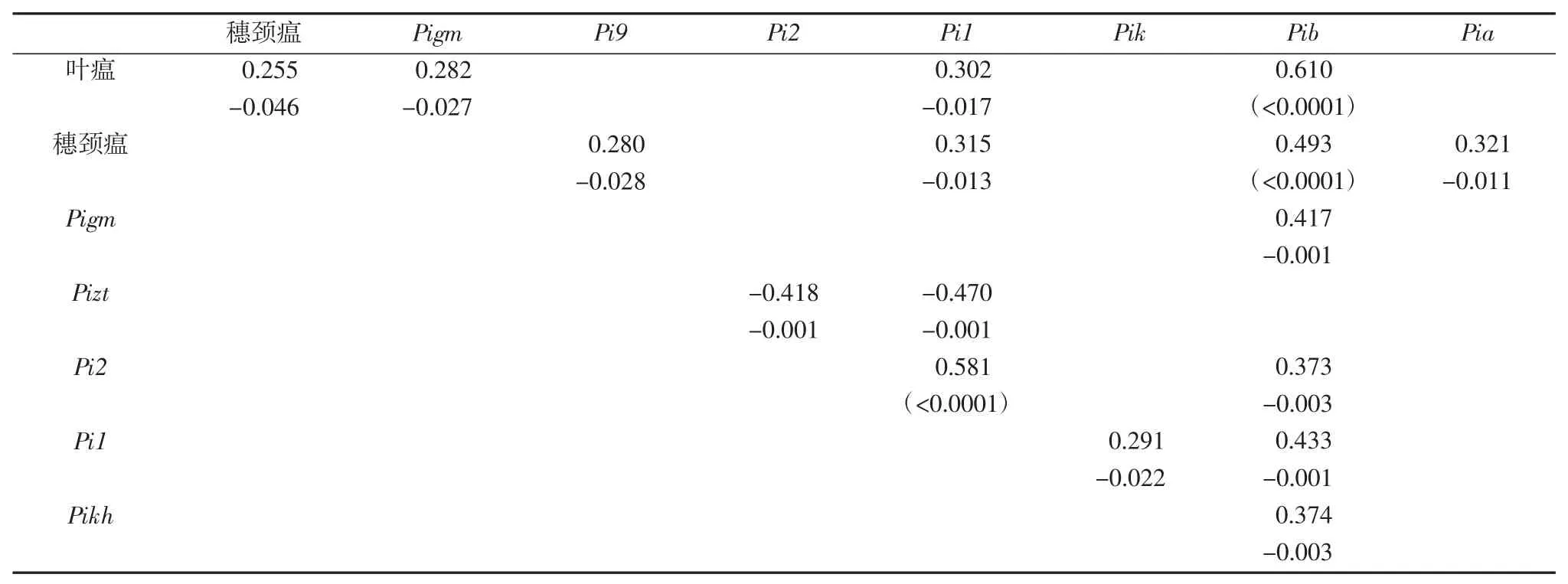
表3 抗性反应与抗病基因以及抗病基因之间的相关性
在 10 个抗稻瘟基因之间,Pigm 与 Pib、Pizt 与 Pi2、Pizt 与 Pi1、Pi1 与 Pib、Pikh 与 Pib 之间的相关性极显著,Pi1 与Pik 相关性显著。Pi1 和Pib 多与其他抗病基因的相关性达到显著及以上水平(表3),说明Pi1 和Pib 抗病基因与其他基因一起聚合对水稻抗病育种具有重要作用。
2.4 抗病基因间的遗传聚类分析
对浙江省63 个主栽品种的叶瘟指数、穗瘟指数和10 个抗稻瘟基因的聚类分析显示,在分类距离为1 时可以将63 个水稻品种分出3 大类:JD036 和JD048 为一类,JD052 和JD062 为一类,其余为一类,在分类距离0.9~1 之间又可以将第三大类细分为三类(图1)。通过聚类分析所分出的类群中,同一类群中的品种抗性类似。这为以后品种改良选择提供了方向,同类中的基因重组有利于水平抗性的提高,类群外的基因重组可以提高广谱抗稻瘟病性。
2.5 抗病基因的频率分布
不同抗病基因在籼稻和粳稻两类材料中分布存在差异(图 2)。抗病基因 Pia、Pik、Pi1、Pi2、Pizt、Pikh、Pi5及Pib 在浙江省主栽品种中分布频率整体分布较高,而Pigm 和Pi9 分布频率分布较低。在38 个粳稻材料中,Pib 的频率分布最大为 0.76,其次是 Pia、Pik、Pi1、Pi2、Pizt、Pikh、Pi5、Pi9 和 Pigm,分布频率分别为 0.66、0.55、0.47、0.29、0.26、0.13、0.05 和 0.03。在 25 个籼稻材料中,分布频率最大的也是Pib 为0.64,其次是Pi5、Pia、Pikh、Pi1、Pik、Pizt、Pi2 和 Pigm,分布频率分别为0.60、0.48、0.44、0.36、0.32、0.32 和 0.04。Pi9 在籼稻分布频率为0。抗病基因Pi5 和Pikh 在籼稻和粳稻中分布频率差异较大。
3 讨论
稻瘟病在水稻的不同生育时期、不同部位均可以侵染,其中,穗瘟对水稻产量威胁最大。叶瘟可以作为指标之一来预测穗瘟的发生,对防治穗瘟发生具有指导意义。本研究对63 个水稻品种的叶瘟和穗瘟进行相关性分析,发现叶瘟和穗瘟间存在显著的相关性。叶瘟和穗瘟抗性反应一致的材料占比达70%(表2)。张品辉等[17]利用贵州麻江县10 年叶瘟和穗瘟的数据发现,叶瘟和穗瘟存在极显著水平的回归关系。陈福如等[18]用人工接种研究叶瘟与穗瘟的关系发现,叶瘟和穗瘟的抗性存在一定的正相关性。这些结果表明,一定程度上可以用室温人工鉴定的叶瘟结果来预测田间穗瘟发生情况,这为稻瘟病抗性鉴定效率的提高提供了参考。
抗病育种是控制稻瘟病最绿色、经济和安全的方法,抗病基因的有效利用是抗病育种的关键。了解当前浙江省主栽品种中抗病基因的构成及与抗性水平的相关性,可为抗病品种的合理布局及提高抗病育种的可预见性提供依据。本研究以63 个浙江省主栽品种为材料 , 利 用 Pi1、Pi5、Pi2、Pi9、Pia、Pizt、Pigm、Pib、Pik 和Pikh 等10 个抗病基因的分子标记,分析这些抗病基因在主栽品种中的分布规律以及其对叶瘟和穗瘟抗性的贡献。Pigm 和Pi9 在63 个材料占比比较少,检出率只有3.2%,其他抗病基因检出率都在50.0%左右。Pigm、Pi1 及Pib 与叶瘟抗性存在显著及以上的正相关性,Pi9、Pi1、Pib 和Pia 对穗瘟抗性存在显著及以上的正相关性,暗示着使用这些抗病基因进行水稻新品种培育将有效降低稻瘟病的发生。
抗病基因间复杂的互作关系使得育种家进行多个抗病基因聚合时需要注意所聚合基因的主效性和正向性[19]。本研究发现,Pigm 与 Pib、Pi2 与 Pi1、Pi2 与 Pib、Pi1 与 Pik、Pi1 与 Pib 以及 Pikh 与 Pib 之间是正向显著相关,而Pizt 与Pi2、Pizt 与Pi1 是负向极显著相关,这些结果为未来多个抗病基因聚合提供了重要参考。Pi9对穗瘟抗性具有重要贡献,Pigm 对叶瘟具有重要的贡献,而Pi9 和Pigm 在本研究63 个主栽品种中检出率比较低,说明Pigm 和Pi9 基因在浙江省抗性育种中还有利用空间。关注这2 个基因的分子选择育种可能有助于进一步提高新品种的抗病性。
