经典Wnt信号通路对人牙周膜干细胞成骨分化的影响
2021-12-02刘冬巧王天齐段小妮张淋坤张春香
刘冬巧,王天齐,段小妮,颜 艳,张淋坤,张春香
1.郑州市口腔医院正畸科(郑州450000);2.南开大学医学院口腔医学系口腔正畸学(天津300071)3.天津市口腔医院正畸科,天津市口腔功能重建重点实验室(天津300041);4.天津医科大学口腔医学系口腔正畸学(天津300070);5.天津市口腔医院中心实验室(天津 300041)
牙周膜干细胞(periodontal ligament stem cells,PDLSCs)是从牙周韧带中分离出来的一种新的间充质干细胞(mesenchymal stem cells,MSCs),在维持牙周组织内稳态、牙周组织再生和正畸治疗等方面发挥着重要作用。由于人PDLSCs有可能在体内形成牙骨质/牙周韧带(periodontal ligament,PDL)样结构,为牙周炎、牙合创伤或正畸矫治力损伤的牙周组织的重建和再生提供了新方法[1]。最近,有研究证明Wnt信号通路在骨再生过程中起着重要的作用[2-4]。目前已鉴定的Wnt家族蛋白有19种,根据其稳定细胞内β-catenin 的作用,分为两类:经典和非经典。经典Wnt 信号传导通路是通过β-catenin 介导的,它在经典Wnt 信号传导通路中起着关键作用。而非经典Wnt/Ca2+信号传导途径可激活Ca2+/钙调蛋白依赖性蛋白激酶II(CaMKII)[5],其信号传导不依赖于β-catenin。Wnt 分子是一个分泌蛋白家族,一些Wnt蛋白,例如Wnt1、3a、4、5、10b和13,在成骨细胞形成中起着重要作用[6]。在缺少Wnt 蛋白的情况下,β-catenin 与含有腺瘤性息肉病大肠杆菌(adenoma⁃tous polyposis coli,APC)、糖原合成酶激酶-3β(glyco⁃gen synthase kinase-3 beta,GSK-3β)、Axin 蛋白和酪蛋白激酶1(casein kinase 1)形成复合物。在这个复合体中,GSK-3β组成性磷酸化β-catenin,导致其泛素化,并被26S蛋白酶体降解。在Wnt刺激下,Wnt蛋白与卷曲(Fz)受体和低密度脂蛋白受体相关蛋白5/6(lowdensitylipoprotein receptor-related protein 5/6,LRP5/6)共受体的相互作用导致Axin/APC/GSK-3β活性的抑制[7]。这种复合体的破坏导致活性去磷酸化β-catenin 的稳定和核移位,进而激活淋巴增强因子-1(lymphoid enhancer-binding factor-1,LEF-1)/T细胞因子(T lymphocyte factor,TCF)转录系统,参与细胞周期进程和分化的调节[8]。
有研究表明Wnt/β-catenin 信号通路控制着干细胞向成骨细胞的分化[9-10]。已有研究发现,Wnt信号通路通过直接刺激Runx2 基因表达促进成骨作用,这一功能在成骨细胞形成中起重要作用。同样,小鼠体内Wnt10b的过度表达可以抑制骨丧失,并通过诱导Runx2使骨髓MSCs向成骨细胞系转移。LiCl刺激MSCs 作为Wnt 信号通路的激活剂,可以诱导骨髓MSCs分化为成骨细胞[11]。与之相反,Wnt通路抑制因子(dickkopfs1,DKK1)则可以与LRP5 结合并阻断Wnt/β-catenin信号通路,从而减少骨形成[12]。遗传学研究发现Wnt/β-catenin 信号通路在间充质组织发育过程中起重要作用,其中包括骨骼成熟和牙齿形成[13-14]。然而,经典Wnt/β-catenin 信号通路对骨髓MSCs 成骨分化的影响仍存在争议[10,15]。经典Wnt/β-catenin 信号通路与人PDLSCs 成骨分化的潜在关系尚不清楚。在本项研究中,我们试图研究经典Wnt/β-catenin信号对人PDLSCs成骨分化的调控。
1 材料与方法
1.1 细胞培养
根据以往方法分离和培养人PDLSCs[16]。人PDLSCs取自12~16岁因正畸减数拔牙受试者的前磨牙,要求前磨牙没有龋坏以及其他症状,并且患者知情同意。第2~4 代的PDLSCs 通过CD146 免疫磁珠试剂盒(Miltenyi Biotec,Bergisch Gladbach,Germa⁃ny)进行PDSLCs 的免疫磁珠分离。使用STRO-1,CD146,CD271 和Scleraxis 抗体,通过免疫细胞化学染色将分选的CD146+细胞鉴定为人PDLSCs[16]。为了研究PDLSCs的多向分化能力,对分离的细胞进行成骨和成脂分化能力的检测[16]。将人PDLSCs传代3~4代后用于后续实验研究。细胞在含有5%胎牛血清、10 nM 地塞米松、10 mM β-甘油磷酸酯和50 μg/mL L-抗坏血酸的成骨培养基中培养3 d,培养基分为添加和不添加氯化锂(GSK3β抑制剂)或豆蔻素(Wnt/β-catenin 抑制剂)。人PDLSCs 的收集和培养得到了南开大学天津市口腔医院伦理委员会的批准(2009026)。
1.2 细胞活力测定
将人PDLSCs接种于6孔板中,分别用豆蔻素或氯化锂处理3 d。使用Beckman Coulter Vi-Cell 测量三次细胞活力。
在细胞培养第5 d,使用豆蔻素或LiCl最后一次处理样本后2 h,用Trizol 试剂(Invitrogen)分别提取不同培养条件下人PDLSCs的总核糖核酸(ribonucle⁃ic acid,RNA)。然后用分光光度计测定所提取RNA的浓度和纯度,结果显示总RNA的纯度高达95%或者更高。RNA 分离后,使用SYBR-PrimeScriptTM逆转录聚合酶链反应试剂盒(中国,Takara),在每10 μL反应体积中使用500 ng RNA 进行反向转录脱氧核糖核酸(complementary deoxyribonucleic acid,cD⁃NA)合成,每个反应体积包含0.5 μL PrimeScriptTMRT酶混合物I,0.5 μL oligo dT引物,2.0 μL PrimeScriptTM缓冲液和0.5 μL 随机6 聚体。使用ABI PRISM 7300实时PCR 系统(Applied Biosystems,US)重复测量三次RT-PCR。根据GenBank 数据库合成了Runx2、ALP、Col-I和GAPDH的引物序列,如表1所示。本实验采用2-ΔΔCT方法计算基因表达水平。实验数据显示为相对于对照组的折叠变化。为保证扩增产物的纯度,绘制了每次PCR反应的熔融曲线。

表1 RNA提取及实时定量RT-PCR
1.3 Western blot分析
从培养液取出细胞后,用冷PBS洗涤2次,在缓冲液(Keygen total protein extraction kit,Keygen Bio⁃tech,China)中裂解20 min。然后将裂解后的混合物在4 °C 下14 000 r 离心15 min 后,收集上清液,用BCA蛋白检测试剂盒进行定量测定。每个样本为30~ 50μg 等量的细胞裂解物,将其放在10 %SDS-PAGE 凝胶上运行,然后转移到PVDF 膜(Bio-Rad)上。如前所述进行Western blot 分析[16]。封闭后,用抗β-catenin、Osterix、OPG、RANKL 和GAP⁃DH(Abcam,Cambridge,MA,USA)的一抗进行检测,然后加入辣根过氧化物酶偶联的二抗(中国北京中山生物有限公司)。之后使用化学发光试剂盒(Milli⁃pore,Billerica,MA,USA)观察免疫反应蛋白,使用ChemiDoc XRS 凝胶文档系统和Quantity One 软件(Bio-Rad,Hercules,CA,USA)测定条带强度。
1.4 统计学分析
本实验数据均使用SPSS 17.0(SPSS Inc,Chicago,IL,USA)进行处理,计量资料用平均值±标准差()表示,多个样本均数比较采用单因素方差分析(one-ANOVA)进行组间比较。P <0.05 为有统计学意义。
2 结果
2.1 细胞活力测定
豆蔻素的结构如图1A 所示。我们用Beckman Coulter Vi-Cell 检测了豆蔻素和LiCl 对细胞活力的影响。另外在进行六组数据的单因素方差分析前先进行整体分析,结果显示各组之间细胞活力的差异无统计学意义(F=2.170,P>0.05)。如图1B 所示,各组数据无统计学差异(P>0.05),即豆蔻素、二甲基亚砜(DMSO)和LiCl没有表现出任何明显的细胞毒性作用。

图1 豆蔻素结构以及细胞活力检测结果
2.2 豆蔻素通过促进β-catenin 降解来抑制Wnt/β-catenin的信号转导
为了检验二甲基亚砜(DMSO)对实验是否有干扰,我们比较了DMSO 组和正常对照组的蛋白质表达情况。如图2所示,β-catenin和Osterix在两组间的表达无明显差异(P>0.05),说明DMSO对实验的影响可以忽略不计。用抗β-catenin抗体进行免疫印迹试验分析,观察豆蔻素对细胞内β-catenin水平的影响。如图3 所示,β-catenin 水平降低。GSK-3β导致β-catenin的降解,而LiCl是一种GSK-3β的抑制剂,我们用豆蔻素和LiCl 来培养牙周膜干细胞来排除GSK-3β的影响。Western blot 分析结果显示,DMSO组和正常对照组有统计学差异(P <0.05),豆蔻素可降低细胞内β-catenin的表达。

图2 DMSO组和正常对照组的蛋白表达情况
2.3 经典Wnt通路对PDLSCs成骨分化的促进作用
LiCl 是Wnt/β-catenin 信号通路的激活剂,提高了β-catenin 的表达(图3)。通过评估成骨细胞特异性转录因子Osterix 蛋白表达的变化,以剂量依赖的方式支持Wnt信号在hPDLSCs中的成骨潜力(图3)。在进行五组数据的单因素方差分析前先进行整体分析,结果显示各组之间细胞活力的差异有统计学意义(F=6.270,P <0.05)。Western blot 分析结果表明,LiCl 培养3 d 后,OPG 的表达增加(P <0.05),RANKL 表达无明显变化(P >0.05),因此OPG/RANKL比值增加(图4)。为了证实LiCl促进成骨的作用,我们采用了实时RT-PCR的方法,以ALP、Col-I和Runx2 作为其他成骨靶基因的例子。经过LiCl 处理后,各组信使核糖核酸(messenger ribonucleic ac⁃id,mRNA)水平均呈剂量依赖性升高(图5)。上述结果表明,经典Wnt 通路促进PDLSCs 向成骨细胞分化。

图3 各组Western blot分析结果
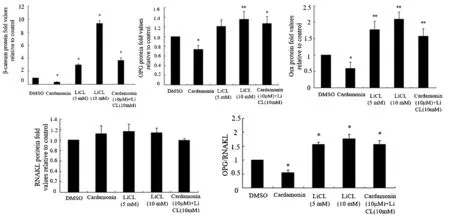
图4 各组蛋白表达情况
2.4 豆蔻素对PDLSCs成骨分化的抑制作用
为了确定经典Wnt/β-catenin 信号通路是否对PDLSCs的成骨有正向调节作用,将这些细胞与豆蔻素培养,豆蔻素可促进细胞内β-catenin 的降解,并抑制经典的Wnt/β-catenin信号传导。对五组数据进行单因素方差分析前先进行整体分析,显示各组之间的表达量差异有统计学意义(F=8.18,P <0.05)。最终的结果表明豆蔻素能降低Osterix 和OPG 的表达,导致OPG/RANKL 比值降低(图4)。同时豆蔻素也降低了成骨因子ALP、Col-I 和Runx2 的mRNA 水平。此外,豆蔻素减弱了LiCl诱导的成骨分化刺激作用,提示豆蔻素通过典型的Wnt/β-catenin信号通路抑制成骨分化(图5)。

图5 各组mRNA表达情况
3 讨论
人牙周膜组织源性间充质干细胞(PDLSCs)具有克隆形成能力强、增殖能力强等特点,可用于干细胞介导的治疗和组织工程。正畸牙齿的移动是由于外加的机械力而引起的组织重建的结果。PDLSCs则在正畸牙齿的移动过程中和过程后发挥着动态作用。组织再生和重建的过程受不同的信号通路控制。来自几个团队的最新研究阐明了经典Wnt/β-catenin信号通路在骨骼发育和出生后骨量维持中的重要性[2]。作为经典Wnt信号通路协助受体,人类的LRP5如果发生功能缺失的突变,就会导致骨质疏松症-假性胶质瘤综合征,在这种综合征中,人们表现为低骨密度和骨骼脆性。相反,激活人类LRP5 的突变发生会导致患者骨密度增加[17]。几项对于基因敲除小鼠的优等研究也证实,经典的wnt/β-catenin 信号通路对于骨骼发育和出生后骨量的维持是必不可少的[2-3]。
Wnt信号通路通过控制间充质干细胞或成骨细胞的功能进而参与成骨的过程。细胞和分子水平的研究表明,β-catenin 和Tcf 蛋白共同调节成骨细胞对于OPG的表达,OPG是破骨细胞分化的主要抑制剂。在βcat(ex3)ob 成骨细胞中,OPG 的表达明显增加,而作为主要破骨细胞分化因子的RANKL的表达则略有增加[18]。在矿化培养基中分化5 d 和10 d 后,β-catenin 成骨细胞中OPG mRNA 的表达与对照组相比降低,而RANKL的表达增加[3]。Wnt/β-catenin信号通路会促进MSCs(骨髓间充质干细胞)向成骨细胞分化[10]。用LiCl刺激MSCs,作为Wnt信号通路的激活剂,可以诱导MSCs向成骨细胞分化[11]。相反,Dick⁃kopfs1(DKK1)抑制Wnt/β-catenin 信号转导,可与LRP5 结合,阻断Wnt/β-catenin 信号转导,抑制BM⁃SCs 的成骨分化,减少骨的形成[12]。这些重要的发现表明,Wnt/β-catenin 信号通路的激活可能有助于改善骨或牙齿的组织工程。在牙齿发育过程中,人们发现经典Wnt信号通路作用于牙齿形态形成的多个阶段[19]。虽然Wnt 信号通路在牙周发育和动态平衡中的作用尚不清楚,但越来越多的数据表明,Wnt信号通路的激活是牙周组织(如牙骨质、牙周膜和牙槽骨)再生所必需的。然而,经典Wnt 信号通路与PDLSCs成骨分化之间的潜在关系尚不清楚。
在本项研究中,通过在人体PDLSCs 中加入GSK-3β抑制剂LiCl来模拟Wnt的作用,并检测经典Wnt信号通路是否对这些细胞的成骨分化有积极的影响。经过LiCl 的处理可升高PDLSCs 在成骨诱导培养基中的成骨特异性基因和蛋白的表达。LiCl 可促进OPG的表达,但不改变RANKL的表达,从而增加OPG/RANKL的比值。这一结果表明,使用可以上调经典Wnt信号通路的药物可以促进PDLSCs的成骨分化。成骨转录因子包括Runx2 和Osterix 是调节成骨细胞定位和间充质细胞成骨分化所必需的[20]。与以前的研究一致,本研究表明LiCl 以剂量依赖的方式提高Osterix、Runx2、ALP 和Col-I 的水平。以前的研究表明,Wnt 信号通路上调β-catenin 蛋白水平足以促进Runx2的表达,从而导致成骨细胞分化。反之,作为一种新型的Wnt/β-catenin信号的小分子抑制剂,不论有没有LiCl,豆蔻素能以剂量依赖性下调细胞内β-catenin 的表达[21],从而抑制成骨因子的表达,但不会改变RANKL 的表达。豆蔻素会减弱LiCl的一些积极作用,这表明豆蔻素可通过经典Wnt 信号通路抑制hPDLSCs的成骨分化。Cho等[22]的研究表明,剂量≤10 μM 的豆蔻素可抑制65%~ 70%的β-catenin 活性,而不影响细胞活力。也有学者指出豆蔻素引起ALP、OCN、OPN、BSP、CAP、CEMP1等成骨基因表达下降,从而导致相应的蛋白合成减少[23]。瞬时受体电位通道(TRP)参与了疼痛信号转导通路的激活或调节。有研究表明豆蔻素可以通过抑制TRPV1 离子通道发挥显著的镇痛作用[24]。为了避免此项干扰作用,提示我们后续实验可以采用特定siRNA 或者其他Wnt 通路抑制剂,用以佐证豆蔻素在Wnt 信号通路中的作用。细胞内β-catenin 水平受多种途径控制:GSK-3β依赖途径、PKCα依赖途径和Siah依赖途径。在GSK-3β依赖途径中,GSK-3β与腺瘤性息肉大肠杆菌和轴蛋白形成破坏复合物,磷酸化β-catenin的末端丝氨酸/苏氨酸残基,通过泛素依赖机制促进其降解[25]。β-catenin的N-末端丝氨酸/苏氨酸残基也会被PKCα磷酸化,也会导致泛素依赖的β-catenin 降解[26]。在Siah 依赖的途径中,Si⁃ah-1与腺瘤性息肉病结肠腺瘤性结肠炎的羧基末端相互作用,从而吸附泛素化复合体,促进β-catenin的降解[27]。在本研究中,豆蔻素能诱导细胞内β-catenin在LiCl(已知的GSK-3β抑制剂)存在的情况下发生降解。这些结果表明,豆蔻素胞内降解β-catenin 的途径可能是通过Siah 途径和/或依赖PKCα途径,而不是通过GSK-3β途径。经典Wnt/β-catenin信号通路通过一系列机制增加骨量,包括干细胞更新,刺激成骨细胞复制,诱导成骨,抑制成骨细胞和骨细胞的凋亡[25]。Heo 等[28]的研究表明,在成骨转录因子的刺激下,经典Wnt 信号通路会导致PDLC 分化为成骨细胞系。Wnt/β-catenin 信号通路会促进祖细胞向成骨细胞分化[9]。与之前的一些研究结果一致,本研究发现Wnt/β-catenin信号通路促进了PDLSCs 的成骨分化。与之相矛盾的是,有多篇文章表明Wnt/β-catenin 信号通路在体外会抑制MSCs的成骨分化和矿化[15,29]。例如,一项研究表明,wnt/β-catenin 信号通路会负向调节牙髓干细胞向成牙本质细胞的分化[14]。Liu 的研究指出,β-catenin 的激活抑制了非经典的Wnt/Ca2+途径,导致炎症微环境中PDLSCs 的增殖增加,但成骨分化减少[30]。Nemotot等[31]观察到Wnt信号通路会抑制成牙骨质细胞分化并促进细胞的增殖。鉴于祖细胞成骨分化调控的复杂性,我们要阐明Wnt/β-catenin 信号通路在调控MSCs成骨分化中的确切机制是非常困难的。在这方面,我们的研究结果与前面的一些研究结果是一致的[11,13-14],但与其他研究人员也有不同之处[14-15,29]。我们认为这种差异在于Wnt靶基因转录的基因特异性激活因子和/或抑制因子存在于细胞环境中,这或许取决于细胞类型和/或分化阶段[32]。在未来,开发一个模型系统来测试规范的Wnt 信号对人体PDLSCs 增殖和成骨分化的影响将会是很有意义的。综上所述,本文报道的结果首次证明Wnt/β-catenin 信号可以促进PDLSCs 的成骨分化。我们的研究结果表明,对经典Wnt 信号通路进行药理学调节可以提高hP⁃DLSCs 的成骨能力,为牙周组织工程提供了一种可行的策略。据了解,正畸治疗后复发的风险一直会持续到牙槽骨改建完成,因此我们的研究结果启发我们提出,通过调节Wnt/β-catenin 信号对PDLSCs 成骨分化的影响,正确调控正畸牙移动后的牙槽骨改建,可以预防移动牙齿的复发。LiCi是Wnt通路激活剂,本研究的局限性在于只检验mRNA 的表达不足以证明经典Wnt信号通路对人牙周膜干细胞的成骨分化具有正向调节作用,后续应增加实验增强结论的正确性。
4 结论
经典Wnt 信号通路对人PDLSCs 的成骨分化有正向调节作用,为牙周组织工程中经典Wnt/β-catenin信号通路对人牙周膜干细胞成骨分化的药理学调控提供了一种可行性策略,同时也为促进正畸治疗过程中的牙齿移动以及随后的牙周组织改建提供了思路。
