绣球菌子实体单组分多糖结构表征及其免疫活性
2021-11-22郝正祺刘靖宇孟俊龙常明昌冯翠萍
郝正祺,刘靖宇,2,孟俊龙,2,常明昌,2,冯翠萍*
(1 山西农业大学食品科学与工程学院 山西太谷 030801 2 山西省食用菌工程技术研究中心 山西太谷 030801)
多糖是一类由糖苷键聚合形成的生物大分子,主要来源于动植物及微生物,在生物体中发挥着信号传导、免疫调控和物质运输等作用[1]。真菌多糖具有广泛的生物活性,特别是在免疫调节方面,常被作为免疫调节剂和抗癌药物应用于临床治疗中[2]。有研究表明,真菌多糖的生物活性与其分子质量、分支程度、三螺旋结构、单糖组成和糖苷键类型有关,如高分子质量多糖比低分子质量多糖具有明显的抗肿瘤和抗氧化功效;巨噬细胞分泌NO 能力受多糖化学组成和糖苷键类型的影响等[3-5]。
巨噬细胞是连接机体非特异性免疫和获得性免疫之间重要的桥梁,它不仅吞噬并消化细胞残片和病原体,还可以激活淋巴球或其它免疫细胞,令其对病原体作出反应,在宿主的免疫应答过程中发挥着重要作用[6-7]。巨噬细胞常被作为评价活性多糖免疫调节特性的理想细胞模型。
绣球菌(Sparassis latifolia),又名绣球菇、绣球蕈,是一种珍稀名贵的食药用真菌,具有丰富的矿物质、氨基酸、维生素等营养成分,β-葡聚糖含量高达39.9%~43.6%[8-10]。绣球菌β-葡聚糖以带有1,6 分支的β-(1,3)-D-葡聚糖为主,分支频率约为3 个主链单位1 个分支,还有少量的β-(1,6)-D-葡聚糖和ɑ 型葡聚糖,具有多种生理及药理学功效[11],可抑制肿瘤诱导的血管生成及黑色瘤细胞的代谢,刺激外周血单核细胞及小鼠脾细胞产生细胞因子,提高动物的造血功能,同时绣球菌多糖还可提高炎症相关因子IL-12,IL-1β 等基因的表达量,促使树突状细胞的成熟[12-14]。目前对绣球菌多糖纯化后均一多糖组分的结构、免疫活性及其构效关系研究较少。本研究组采用聚能超声波辅助水提醇沉法提取绣球菌多糖,分离纯化后测定其分子质量、单糖组成、结构、表观形貌等,研究其免疫活性,探究其均一多糖组分结构表征与免疫活性之间的关系,为绣球菌多糖功能食品原料的开发利用提供理论依据。
1 材料与方法
1.1 试验材料
绣球菌子实体,由山西省太和食用菌栽培基地提供。
1.2 主要试剂
核糖、木糖、阿拉伯糖、甘露糖、半乳糖、葡萄糖、果糖等单糖标准品,DPPH、Tris、邻苯三酚,美国Sigma 公司;大孔树脂HZ-830、DEAE-52 纤维素粉末、Sephadex-G200 粉末、透析袋(8 000~14 000 u)、T-右旋糖酐(T-10,T-40,T-70,T-500,T-2000) 标品、DMEM 高糖培养基、青链霉素混合液、胰蛋白酶、中性红染料、脂多糖(Lipopolysaccharides,LPS)、MTT 细胞增殖及细胞毒性检测试剂盒、一氧化氮(NO)测定试剂盒,北京索莱宝科技有限公司;10×上样缓冲液、RNAiso Plus 试剂盒、Prime Script TM RT Master Mix(Perfect Real Time) 试剂盒、SYBRRPremix Ex TaqTM II (Tli RNaseH Plus)试剂盒,日本Takara公司;其它试剂均为国产分析纯级。
1.3 主要设备与仪器
超声波细胞破碎仪(Scientz-1200E),宁波新芝有限公司;中压特制玻璃层析柱 (2.6 cm×30 cm),山东化工研究院;恒流泵(DHL-ZB)、自动部分收集器(DBS-100A),沪西仪器厂;旋转蒸发仪(RV10),IKA 公司;冷冻干燥机(PL 3000),Thermo 公司;傅里叶变换红外光谱仪(Tensor27),Bruker 公司;紫外分光光度计(Cary 4000)、凝胶渗透色谱,Agilent 公司;离子色谱仪(ICS-3000),DIONEX 公司;超导核磁共振波谱仪(600 MHz),Bruker 公司;扫描探针显微镜(SPA-300 HV),日本精工;扫描电镜(JSM-6010),JEOL日本电子;凝胶色谱柱(TSK-GEL-4000PWXL),日本Tosoh 公司;CO2培养箱 (HF 90 / HF 240),Heal Force公司;荧光倒置显微镜(IX2-UCB),OLYMPUS 公司;多功能酶标仪 (Sepctramax i3X),Molecular Devices 公司;MyCyler PCR 仪,Bio-Rad 公司;荧光定量PCR 仪(QuantStudio 7 Flex),Thermo Scientific 公司。
1.4 试验方法
1.4.1 绣球菌多糖的制备、分离及纯化 绣球菌粗多糖参照文献[15]的方法制备。将绣球菌子实体样品置于35 ℃烘箱中干燥,切片后粉碎,过200目筛。称取一定量的绣球菌子实体干粉,按料液比1∶40(g/mL)加入纯水,聚能超声波1 200 W 处理20 min,将1%的纤维素酶、中性蛋白酶分别在pH 4 和pH 7,温度40 ℃条件下水解3 h。灭酶后100℃水浴提取3 h,水提液3 500×g离心20 min,取上清,旋蒸浓缩,加入3 倍体积的无水乙醇醇沉,4℃下静置过夜,3 500×g离心15 min 得到沉淀。沉淀分别用丙酮、乙醚洗涤、过滤后收集多糖。采用Sevag 法除蛋白,HZ-830 离子交换树脂脱色、除杂,冷冻干燥后得到绣球菌粗多糖样品,命名为SlPs。
取一定量的SlPs 溶解质量浓度为10 mg/mL的多糖溶液,用0.45 μm 一次性针头过滤器除杂,将5 mL 样品液缓缓加入DEAE-52 阴离子交换柱(2.6 cm×30 cm),用0.1 mol/L NaCl 溶液洗脱,调节恒流泵速度为1.0 mL/min,同时利用自动收集器每5 min 收集1 管(约5 mL)。采用苯酚-硫酸法跟踪检测,收集得到SlPs 洗脱液,浓缩,透析(3 500 u,24 h)去除NaCl,冷冻干燥得到多糖样品。取一定量经DEAE-52 纯化的多糖样品,用0.45 μm 滤器除杂后,将5 mg/mL 溶液缓缓加入Sephadex-100 凝胶过滤柱(2.6 cm×30 cm)进一步纯化,用蒸馏水洗脱,控制恒流泵使其流速为1 mL/min,每5 mL 收集1 管,苯酚-硫酸法追踪检测,直至吸光值出现为止,收集吸收峰的洗脱液,浓缩冻干后得纯化的多糖,命名为SlP。
1.4.2 结构鉴定
1) 均一性和相对分子质量的测定 采用高效凝胶渗透色谱法测定SlP 的平均分子质量,色谱柱为TSK-GEL-4000 PWXL (300 mm×7.8 mm),检测器为折光示差检测器,检测温度40 ℃,上样量20 μL,上样质量浓度2 mg/mL;0.6 mL/min蒸馏水洗脱。采用不同分子质量的T-右旋糖酐标品(T-10,T-40,T-70,T-500 和T-2000)建立标准曲线。标准曲线方程为lg Mw=-0.3826Rt+8.66,R2= 0.9945,式 中,Mw——分子质量(u);Rt——保留时间(min)。
2) 单糖组成的测定 称取10 mg 样品于安瓿瓶中,加入4 mL 2 mol/L 三氟乙酸,用酒精喷灯对安瓿瓶进行封口,在120 ℃油浴中水解6 h。完全反应后冷却至室温,减压条件下将水解物蒸发至干燥,用甲醇洗涤3~5 次,除去剩余的三氟乙酸。采用离子色谱分析单糖组成,通过Dionex ICS 3000 色谱系统分析,色谱柱为Dionex Carbopac PA 10(250 mm×4 mm)。将干燥的水解液溶解在1 mL 蒸馏水中,用10 倍梯度稀释法测定,柱温30 ℃,上样量25 μL,洗脱液为2 mol/L NaOH,流速0.45 mL/min。采用单糖标准品为阿拉伯糖、核糖、木糖、葡萄糖、果糖、甘露糖和半乳糖。
3) 特征吸收峰的测定 采用傅里叶红外光谱法测定多糖特征吸收峰。称取一定量的多糖样品和溴化钾粉末,按质量比1∶50 充分混合,研磨后压片,在波长400~4 000 cm-1的扫描范围内采集数据,通过软件对图谱进行分析。
4) 初级结构的鉴定 称取干燥的100 mg SlP,溶解在装有0.55 mL DMSO 的核磁管中,采用600 MHz 的核磁共振仪在室温下检测13C NMR,1H NMR。
5) 三螺旋结构分析 采用刚果红染色方法分析SlP 的三螺旋结构。配制方法:将2 mL 1 mg/mL SlP 溶液与91 μmol/L 刚果红溶液充分混合,逐滴加入1 mol/L NaOH,使其终浓度在0~0.5 mol/L 范围内增加,用紫外-可见分光光度计在波长400~700 nm 范围内扫描,在不同NaOH 浓度下检测多糖溶液与刚果红溶液的λmax。以NaOH 浓度为横坐标,多糖与刚果红混合溶液最大吸收波长λmax为纵坐标,绘制图像。
6) 表观形貌观测 将冻干的SlP 粉末粘到铜质样品台上,对样品表面喷金,用扫描电镜观察SlP 表面的结构。
7) 分子形貌观测 配制质量浓度为1 μg/mL SlP 溶液,采用0.45 μm 一次性滤器除杂、过滤,取5 μL SlP 溶液滴在新鲜剥离的云母片表面上,室温干燥过夜。室温下采用AFM 观测多糖分子在溶液中的形貌,用NanoScope 软件处理、分析试验图形。
1.4.3 免疫活性的测定
1) 细胞培养 小鼠巨噬细胞RAW264.7,中国科学院上海生命科学研究院细胞资源中心。将细胞置于含10%胎牛血清和2%青霉素-链霉素的高糖培养基(DMEM)中,在温度37 ℃、湿度5%的CO2培养箱中培养24~36 h,达到对数期。
2) 对巨噬细胞RAW264.7 增值活力的影响采用MTT 法测定巨噬细胞增殖活力。将处于对数期的细胞接种于96 孔培养板上,密度为1×106细胞/mL,每孔100 μL。待细胞贴壁后,除空白对照外,分别加入31.25,15.63,7.81,3.92 μg/mL 和1.95 μg/mL 的SlP,每组3 次重复1 次,于温度37℃、相对湿度5%的CO2培养箱中培养24 h。培养结束后,弃培养基,每孔加入20 μL MTT 溶液,在温度37 ℃、相对湿度5%的CO2培养箱中培养24 h。4 h 后弃MTT 溶液,每孔加入150 μL DMSO,置摇床中轻轻振摇15 min,在波长490 nm 处用酶标仪测定OD 值。计算时以仅含多糖的孔为背景误差组,除去背景误差。
3) 对巨噬细胞RAW264.7 NO 分泌量的影响 将巨噬细胞RAW264.7 培养液调整至含量为1×106个/mL,接种到24 孔细胞培养板中,每孔500 μL,置于温度37 ℃、相对湿度5%的CO2培养箱中培养24 h。培养结束后,弃培养基。将含有7.81,3.92 μg/mL 和1.95 μg/mL SlP 的细胞培养液 (设为组I,II 和 III)、阳性对照组10 μg/mL LPS 和空白的细胞培养液加入培养板中,作用24 h 后收集细胞上清液。按照索莱宝一氧化氮(NO)测定试剂盒说明书,测定巨噬细胞RAW 264.7 细胞培养上清中NO 分泌量。
4) 对巨噬细胞RAW264.7 吞噬能力的影响将巨噬细胞RAW264.7 培养液调整至含量为1×106个/mL,接种到24 孔细胞培养板中,每孔500 μL,置于温度37 ℃、相对湿度5%的CO2培养箱中培养24 h。培养结束后,弃培养基。将含有7.81,3.92 μg/mL 和1.95 μg/mL SlP 的细胞培养液 (设为组I,II 和III)、阳性对照组10 μg/mL LPS 和空白的细胞培养液加入培养板中,作用24 h 后,加入0.075%中性红染料,置环境温度37 ℃,相对湿度5%的CO2培养箱内培养1 h,弃旧的培养基,向每孔中加入PBS 反复洗涤3 次。在每孔中加入细胞裂解液,室温反应15 min,在波长540 nm 处测定吸光值。吞噬指数(PI)= ODA1/ODA2,式中,A1、A2分别为样品组和空白组的OD 值。
5) 对巨噬细胞RAW264.7 免疫细胞因子基因表达量的影响 将巨噬细胞RAW264.7 以1 ×106个/mL 含量接种到24 孔培养板中,每孔500 μL。24 h 后换液,将含有7.81,3.92 μg/mL 和1.95 μg/mL SlP 的细胞培养液(设为组I,II 和III)、阳性对照组10 μg/mL LPS 和空白的细胞培养液加入培养板中,孵育24 h 后获得细胞悬液,用于检测 TNF-α、IL-1β、IL-6、IL-3、IL-10 和IFN-β mRNA 相对表达量。采用TRIzol 试剂提取总RNA,定量后用RT-PCR 反转录试剂盒反转录为cDNA。在 GenBank 中找到小鼠巨噬细胞中βactin、TNF-α、IL-1β、IL-6、IL-3、IL-10 和IFN-β的基因序列,通过Primer 5.0 软件设计试验所需引物,由深圳华大基因有限公司合成,其引物序列如下:TNF-α 上游引物5′-TATGGCTCAGGGTC CAACTC -3′ ,下游引物 5′ -GCTCCAGTGA ATTCGGAAAG-3′;IL-1β 上游引物5′-GAGCC CATCCTCTGTGTCTC-3′,下游引物5′-AGCT CATATGGGTCCGACAG-3′;IL-6 上游引物5′-CCGGAGAGGAGACTTCACAG-3′,下游引物5′-TCCACGATTTCCCAGAGAAC-3′;IL-3 上游引物5′-GACCTGAAGGGCACATGAGA-3′ ,下游引物5′-CAAGTGCCAGTGAGTTGCAG-3′;IL-10 上游引物5′-CATTG CATACG GGACAGAACT-3′,下游引物5′-ACTGTTTGAGGGCCACTTCAT -3′ ;IFN-β 上游引物5′-CTCCATTACCATCCGCCTCA-3′,下游引物5′-GCCAGCAGCCTTACGAGATA-3′;β-actin 上游引物5′-AGCCATGTACGTAGC CATCC-3′,下游引物5′-CTCTCAGCTGTGGTG GTGAA-3′。PCR 扩增条件:95 ℃30 s;95 ℃3 s,60 ℃30 s(40 个循环);72 ℃30 s,95 ℃15 s,60 ℃1 min,95 ℃15 s。
1.5 统计分析
2 结果与分析
2.1 SlP 均一性和相对分子质量
如图1所示,SlP 高效凝胶渗透色谱图表现为单一、对称峰,说明SlP 是均一多糖。根据标准曲线方程,测得SlP 分子质量为1.04×104u。

图1 分子量分布Fig.1 The distribution of molecular weight
2.2 单糖组成
如图2所示,SlP 主要由葡萄糖构成。
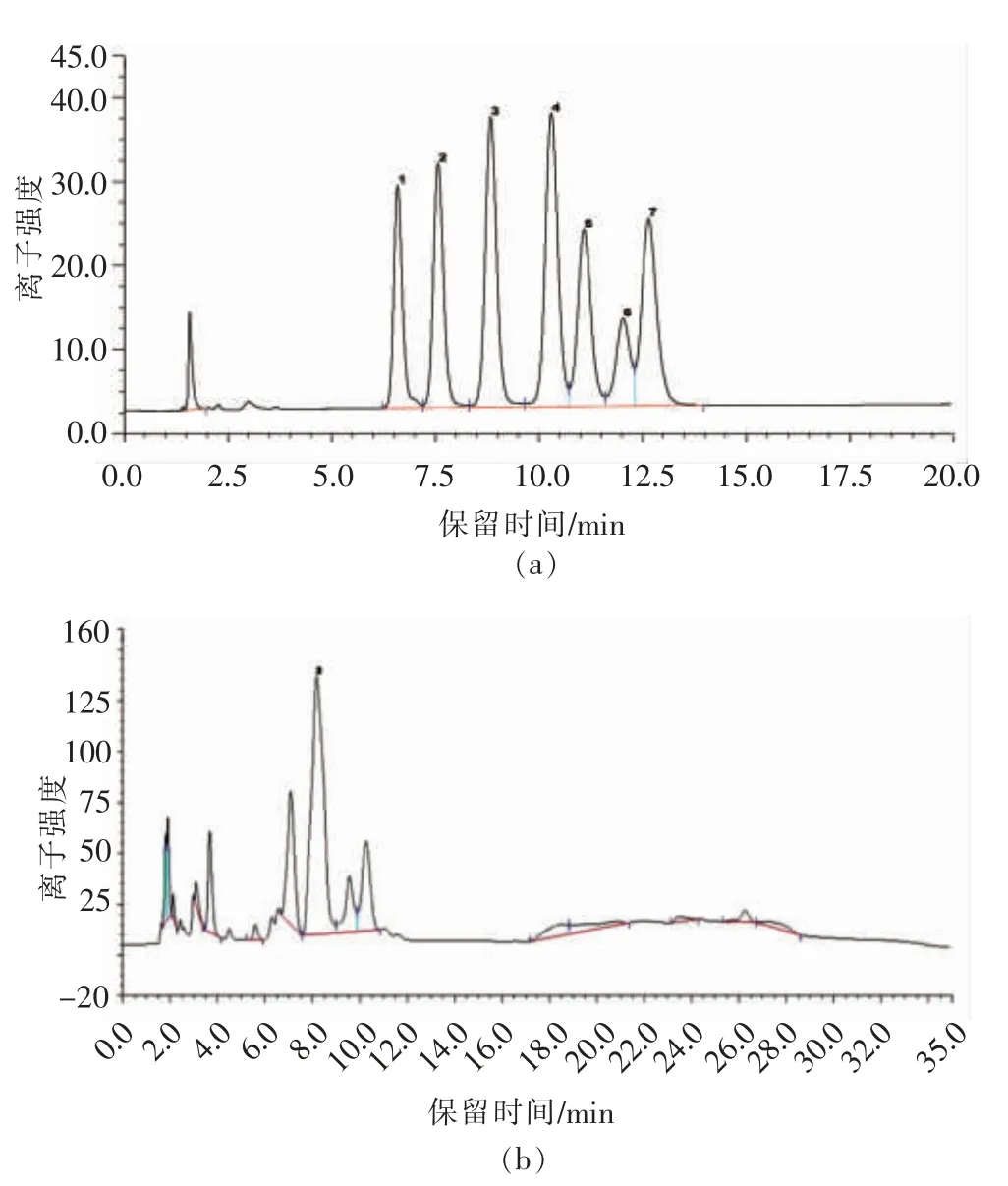
图2 离子色谱图Fig.2 Ion chromatography (IC) chromatograms
2.3 红外光谱分析
如图3所示,SlP 具有典型的多糖吸收峰。在3 425.36 cm-1出现的强吸收峰由多糖分子间游离羟基和氨基官能团的O-H 和N-H 伸缩振动引起;在2 925.60 cm-1出现的中强尖峰由糖链亚甲基间的C-H 伸缩振动引起;在1 641.85 cm-1出现的强吸收峰由多糖分子中-COOH 或-CO-中C=O伸缩振动引起;在1 416.05 cm-1和1 374.38 cm-1处有吸收峰,由C-H 弯曲振动引起;而在1 150.20 cm-1和996.23 cm-1附近出现的中等吸收峰则分别由C-O-H 的拉伸振动和C-O-C 糖苷键的拉伸振动的重叠环振动引起。
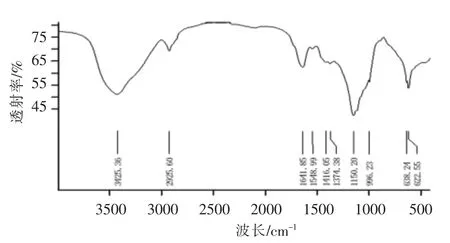
图3 红外光谱图Fig.3 FT-IR spectrum
2.4 核磁共振波谱分析
由图4可知,在1H NMR 波谱中的异头氢信号在δ 4.51~4.96 ppm 范围出现5 个强吸收峰,表明SCP-1 糖苷键中含有β 构型;从13C-NMR 谱图可看出,异头碳区域的共振信号峰出现在δ 103.49×10-6和δ 103.45×10-6处,进一步表明为SlP 为β构型葡聚糖。
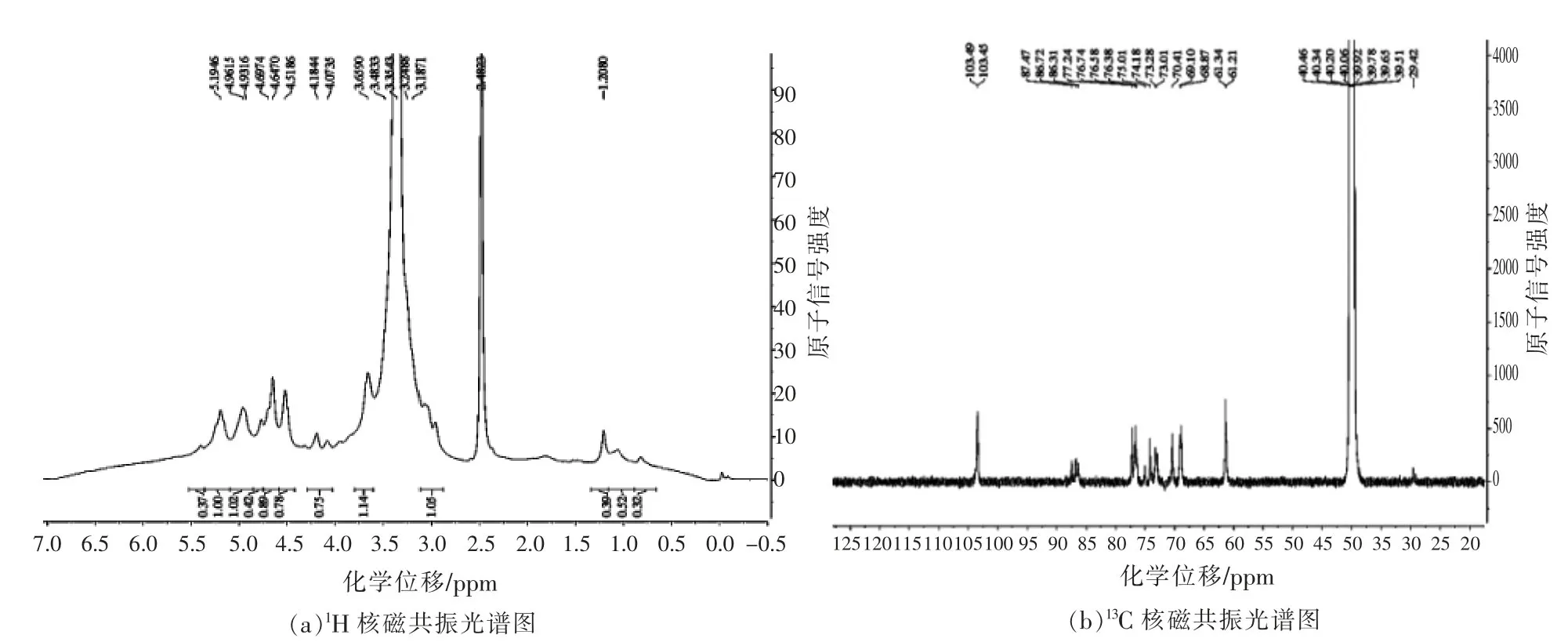
图4 1H 和13C 核磁共振光谱图Fig.4 1H and 13C NMR spectra
2.5 SCP-1 三螺旋结构分析
如图5所示,与刚果红相比,在0.04 mol/L NaOH 条件下,刚果红+SlP 溶液的λmax发生明显的变化,出现红移现象,说明SlP 与刚果红形成复合物,呈三螺旋结构存在;当NaOH 浓度超过0.04 mol/L 时,溶液λmax减小,络合物开始分解,螺旋结构开始解离;当NaOH 浓度接近0.28 mol/L 时,络合物完全分解,螺旋完全解开。由此说明在弱碱性条件下SlP 具有三螺旋结构。
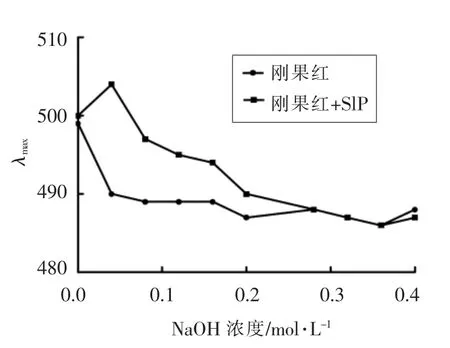
图5 不同浓度NaOH 下刚果红与SlP混合液的λmax 变化图Fig.5 The maximum absorption wavelength of various SlP-Congo red mixture at different concentrations of NaOH
2.6 分子形貌
如图6所示,在扫描范围5 μm×5 μm 下观察SlP 分子间呈现无规则的线团和岛屿状结构,链间缠绕且具有分支状结构;扫描范围800 nm ×800 nm 下,SlP 出现大小、形状不一的球形聚集体,这些聚集体的形成可能是因氢键作用,SlP 的多股单链间相互纠缠缔合所致。SlP 的单链高度和直径分别分布于31.48~42.61 nm 和0.28~1.75 nm范围。

图6 SlP 原子力显微镜Fig.6 AFM images of SlP
2.7 表观形貌
扫描点镜下SlP 表观形貌如图7所示。100 倍下,SlP 呈现大小、形状不一无规则的碎片状;放大300 倍,SlP 碎片一面光滑,一面粗糙。

图7 扫描电镜镜图Fig.7 SEM images
2.8 免疫活性分析
2.8.1 对巨噬细胞增殖活力的影响 从图8可知,SlP 质量浓度在1.95~31.25 μg/mL 范围内,SlP可促进RAW264.7 巨噬细胞增殖。当SlP 质量浓度在7.81 μg/mL 时,促进细胞增殖能力最强。
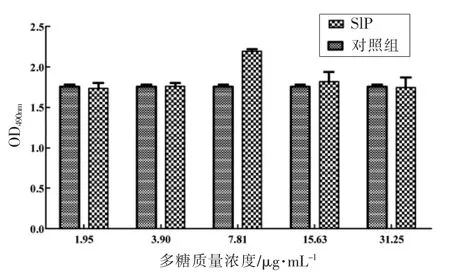
图8 SlP 对巨噬细胞RAW264.7 增殖的影响Fig.8 Effect of SlP on macrophage proliferation
2.8.2 对巨噬细胞NO 分泌量和吞噬能力的影响 从图9可知,与空白对照组相比,10 μg/mL LPS作用RAW264.7 巨噬细胞的12 h,其NO 分泌量、吞噬能力极显著增强 (P<0.01);SlP 质量浓度在1.95~7.81 μg/mL 范围,能够促进噬细胞NO 分泌量、吞噬指数随质量浓度的提高而增大,当多糖质量浓度为7.81 μg/mL 时,NO 分泌量、吞噬指数最大;与LPS 阳性对照组相比,SlP 作用巨噬细胞后,其NO 分泌量、吞噬指数明显低于LPS 组,差异极显著(P<0.01)。
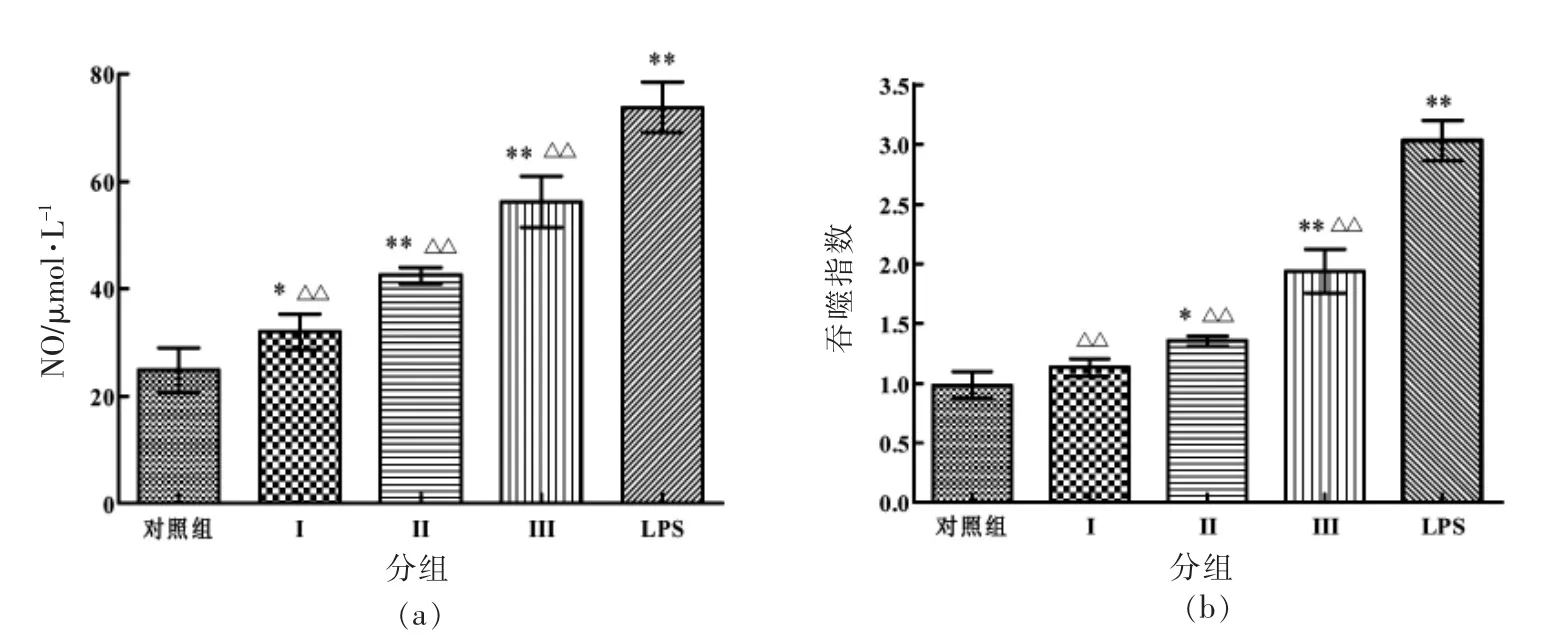
图9 SlP 对巨噬细胞NO 分泌量和吞噬能力的影响Fig.9 Effect of SlP on NO secretion and phagocytic capacity of macrophages
2.8.3 对巨噬细胞免疫细胞因子mRNA 表达量的影响 从图10可知,与空白对照组相比,10 μg/mL LPS 作用RAW264.7 巨噬细胞12 h,TNF-α mRNA、IL-1β mRNA、IL-6 mRNA、IL-3 mRNA、IL-10、IFN-β mRNA 表达量明显升高,差异极显著(P<0.01)。SlP 在质量浓度1.95~7.81 μg/mL 范围内,能提高巨噬细胞内细胞因子mRNA 表达量,随着质量浓度的升高,细胞因子mRNA 表达量逐渐增大,当质量浓度为7.81 μg/mL 时,差异极显著(P<0.01);与阳性对照组相比,LPS 作用后巨噬细胞的细胞因子mRNA 表达量明显高于多糖,差异极显著(P<0.01)。
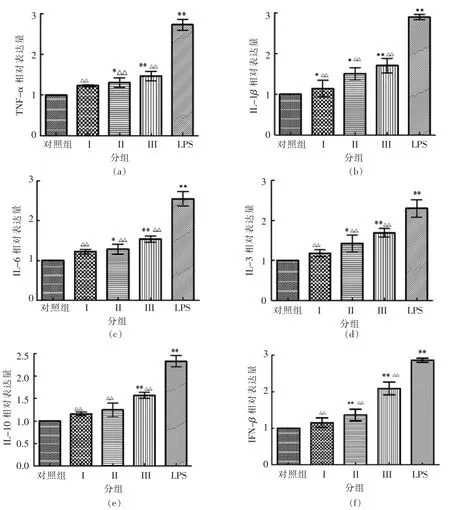
图10 SlP 对巨噬细胞RAW264.7 免疫细胞因子mRNA 表达量的影响Fig.10 Effect of SlP on immunity cell cytokine mRNA expression in RAW264.7 macrophages
3 讨论
多糖对宿主免疫系统的调节作用并非直接作用于机体,主要是通过激活NK 细胞、T 细胞、B 细胞和巨噬细胞依赖性免疫应答系统来间接实现的[16]。巨噬细胞是机体中重要的免疫细胞,在维持机体免疫反应以及非特异性防御中发挥着重要的作用。巨噬细胞能够杀死病原菌和肿瘤细胞,释放出刺激其它免疫细胞的细胞因子,并在细胞表面携带抗原,将其呈递给T 细胞,参与抗原的呈递[17]。β-葡聚糖是一种常见的生物免疫反应调节剂,可与巨噬细胞表面模式识别受体结合,启动免疫反应机制,增加活化巨噬细胞数量。本试验中SlP主要由β-葡聚糖构成,一定浓度的SlP 能够促进巨噬细胞增殖,显著增强巨噬细胞的吞噬能力。
巨噬细胞受到刺激后,可以分泌大量细胞因子,如TNF-α、IL-1β、IL-6、IL-3、IL-10、IFN-β 等,在细胞免疫调节及抑制肿瘤生长等方面发挥着重要的作用。TNF-α 是由激活的巨噬细胞产生的多效性调节因子,在炎症反应启动中发挥着重要的作用,能够提高嗜酸性粒细胞和中性粒细胞功能,刺激超氧化物产生,释放溶酶体酶,介导其它细胞和炎症因子的表达,调节免疫和代谢功能,维持机体平衡[18]。IL-1β 由Th2 型T 辅助淋巴细胞、肥大细胞和B 细胞产生,能够抑制单核巨噬细胞产生促炎细胞因子,促进合成IL-1 受体拮抗剂,间接抑制IL-1 的功能,是一种多效应的炎症因子[19]。IL-6 是宿主对感染和受损组织反应的主要介质,能促进T 细胞发育,刺激B 细胞产生免疫球蛋白,参与炎症反应,在内分泌和造血系统调节过程中发挥作用[20]。IL-3 由活化的T 细胞产生,在NK 细胞以及T 淋巴细胞中表达,能与其它细胞因子联合作用,诱导和扩增树突状细胞产生,对肿瘤抑制具有一定的作用[21]。IL-10 可以抑制炎症细胞的黏附,阻止促炎症细胞因子合成及分泌,降低致炎作用的细胞因子的数量,减轻炎症反应[22]。IFN-β 可以调节T 淋巴细胞和B 淋巴细胞的免疫功能,增强IgG 受体的表达,有利于T 淋巴细胞、B 淋巴细胞的激活,巨噬细胞对抗原的吞噬以及NK 细胞对靶细胞的杀伤,增强机体免疫调节能力[23]。本试验中,一定浓度的SlP 能够提高RAW 264.7 小鼠巨噬细胞中细胞因子TNF-α、IL-1β、IL-6、IL-3、IL-10、IFN-β mRNA 表达量,并呈剂量的依赖性。SlP 对巨噬细胞产生细胞因子的调控可能与其表面膜识别受体有关,SlP 与巨噬细胞表面膜受体如TLR 2 和TLR 4 受体、CR 3 受体、MR 受体等相互作用将信号传递给巨噬细胞,细胞信号通过级联放大作用及不同信号途径之间相互交联作用,调控免疫反应基因的转录,诱发相应蛋白的活性以及表达量的变化,发挥免疫调节作用。关于SCP-1 和巨噬细胞之间特定的免疫调控机制,将将在后续试验中研究。
此外,本研究发现SlP 通过调节巨噬细胞免疫活性来促进NO 的产生。作为一种高效且多样化的生物调节剂,NO 在巨噬细胞的非特异性免疫过程中发挥着重要的作用[24]。NO 不仅能够抑制、杀死肿瘤细胞和致病微生物,还能对其它细胞因子的分泌起到一定调控作用。巨噬细胞产生NO主要是由于一氧化氮合酶 (NOS) 的催化作用引起,诱导型NOS(iNOS)是由巨噬细胞等效应细胞合成,直接参与机体免疫细胞防御及炎症反应。SlP 与巨噬细胞膜表面受体结合,促进细胞内iNOS 表达量的上调,激活细胞的非特异性免疫,诱导细胞内NO 分泌量增加。
4 结论
从绣球菌子实体多糖中纯化的SlP 是一种由葡萄糖构成的β-葡聚糖,分子质量集中在1.04×104u,具有三螺旋结构,表观呈现大小、形状不一、无规则的碎片状,分子间呈现无规则的线团和岛屿状,链间缠绕且具有分支状结构。SlP 能够促进RAW264.7 巨噬细胞增殖,提高巨噬细胞的吞噬能力、NO 分泌量以及细胞内TNF-α、IL-1β、IL-6、IL-3、IL-10、IFN-β 免疫因子mRNA 的表达量。SlP 可作为一种良好的功能食品原料应用于食品工业中。
