瑞士乳杆菌H11发酵乳饮料的ACE抑制活性及代谢组学研究
2021-11-14王昊乾李伯海赵景娜刘文俊陈永福孙天松
王昊乾,李伯海,赵景娜,刘文俊,陈永福,孙天松
(内蒙古农业大学,乳品生物技术与工程教育部重点实验室,农业农村部奶制品加工重点实验室,内蒙古自治区乳品生物技术与工程重点实验室,内蒙古呼和浩特 010018)
活性乳饮料是指将原料乳经过益生菌发酵后得到的具有独特风味特点和有利于人体健康的乳制品[1]。能够用于活性乳饮料中的微生物需具有良好的益生功效,且能够耐受胃肠液并定殖于人体肠道保持活性,进而促进人体健康[2-3]。瑞士乳杆菌属于革兰氏阳性异型发酵乳酸菌,具有较强的蛋白水解系统,能够通过水解牛乳中的蛋白质产生多种代谢产物,如有机酸和多肽[4],这些代谢物有助于发酵乳制品质地和风味的形成[5]。在发酵过程中,瑞士乳杆菌通过蛋白酶水解产生大量的生物活性肽,赋予了发酵乳制品优良的益生特性[6-7]。研究表明,具有生物活性功能的发酵乳制品具有提高免疫力、加强老年人认知功能、抗突变、降血压和抗氧化等健康功效[8-9]。
高血压是威胁人体健康的慢性疾病之一,人体自身可通过多种途径来调节血压,血管紧张素转化酶(ACE)对维持血压平衡起关键作用[10],但药物治疗高血压会带来一系列的不良反应,而酶促水解和微生物发酵牛奶蛋白所释放出的降血压肽不会对人体产生副作用[11]。过去的十几年中,食品衍生肽为预防和治疗高血压方面提供了理论依据[12-13]。
目前市面上常见的发酵乳饮料,如日本的养乐多、伊利每益添和蒙牛优益C等发酵乳饮料基本选择干酪乳杆菌和副干酪乳杆菌作为发酵剂,例如蒙牛优益C选择使用副干酪乳杆菌Lc-01作为发酵剂[1,14],而使用瑞士乳杆菌在功能性饮料的开发和应用方面的研究较少[14]。因此,本文以商业发酵剂副干酪乳杆菌Lc-01作为对照组,以前期筛选的一株具有良好发酵及益生特性的瑞士乳杆菌H11为研究对象,旨在研究瑞士乳杆菌H11在活性乳饮料贮藏期间的ACE抑制活性、挥发性风味物质和代谢物与副干酪乳杆菌Lc-01之间的差异,为开发新型安全高效的益生菌乳饮料提供研究基础。
1 材料与方法
1.1 材料与仪器
瑞士乳杆菌H11 内蒙古农业大学乳酸菌菌种资源库提供,副干酪乳杆菌Lc-01 购自丹麦科汉森公司;血管紧张素转换酶(ACE,0.25 U/g)、马尿酸(HA)、马尿酰组胺酰亮氨酸(HHL) Sigma公司;三氟乙酸分析级、乙腈色谱级、甲醇色谱级 Fisher Chemicals公司;脱脂乳粉 新西兰恒天然公司。
7697A-GC-7890B-MS-5977C气相色谱-质谱联用仪、Agilent1100液相色谱系统 Agilent公司;UPLC-Q TOF-MS超高效液相-四级杆-飞行时间质谱仪 Waters公司。
1.2 实验方法
1.2.1 发酵乳饮料的制备 将13%的脱脂奶粉和2%的葡萄糖溶解于45 ℃蒸馏水中,搅拌均匀,静置30 min[15]。在115 ℃下加热10 min后取出,分装至250 mL无菌瓶中,待其褐变后冷却至37 ℃,分别接种5×106CFU/mL瑞士乳杆菌H11和副干酪乳杆菌Lc-01,在37 ℃下发酵。当滴定酸度(Titer acidity,TA)达到200oT时停止发酵。最后将发酵乳与无菌蔗糖溶液按体积比25:75进行混合,冷藏28 d,在贮藏0、7、14、21、28 d时分别取样,用于分析测定。
1.2.2 体外ACE抑制活性的测定 样品前处理:将样品于4500×g,离心10 min,取上清液,用NaOH调节pH至8.3。取发酵乳饮料上清液和HHL溶液各100 μL混合,于37 ℃下培养2 min,再加入100 μL的ACE,将混合物在37 ℃下孵育40 min。在85 ℃水浴中加热10 min;将反应液注入400 μL的0.1 mol/L EDTA溶液中,使用0.22 μm的滤膜过滤待测。
使用RP-HPLC法测定酶解反应产物中马尿酸的含量。色谱柱为ZORBAX C18(4.6 mm×250 mm,5 μm,Agilent,USA),流动相:体积比为22%乙腈(含0.1%三氟乙酸)和78%去离子水(含0.1%三氟乙酸)溶液;流速为1.0 mL/min;检测波长为228 nm;柱温30 ℃;进样量20 μL。
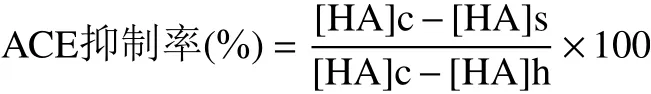
式中,[HA]c:空白对照组中马尿酸浓度;[HA]s:含有ACE抑制剂的待测样品中马尿酸浓度;[HA]h:HHL标品中马尿酸浓度。
1.2.3 ACE抑制肽(Val-Pro-Pro(VPP)和Ile-Pro-Pro(IPP))含量测定 样品前处理:将样品于4500×g,离心10 min,取100 μL样品与400 μL的乙腈混合,于12000×g离心10 min,样品于0.22 μm滤膜过滤后,上机检测。
色谱条件:色谱柱T3 column(2.1 mm×100 mm,1.8 μm);柱温为45 ℃;流速为0.4 mL/min;进样量为4 μL;流动相A为含0.1%甲酸的去离子水;流动相B为含0.1%甲酸的乙腈,梯度洗脱程序(B%):
0~1 min,3%B;1~5 min,3%~30%B;5~5.1 min,30%~90%B;5.1~7.1 min,90%B;7.1~7.2 min,90%~3%B;7.2~9.2 min,3%B。
1.2.4 风味物质的测定 萃取头老化条件:进样口温度250 ℃,老化20 min。
固相微萃取条件:将老化好的萃取头插入样品瓶中,50 ℃,300 r/min下吸附60 min,随后在进样口处250 ℃解吸附3 min。
气相色谱条件:升温程序:起始温度为35 ℃,保持5 min;以5 ℃/min升温至140 ℃,保持2 min;以10 ℃/min升温至250 ℃,保持3 min;汽化室温度250 ℃;载气为氦气(≥99.999%),流速1.0 mL/min;不分流进样。
质谱条件:电离方式为EI源,电子能量70 eV;发射电流100 μA;离子源温度为230 ℃;质量扫描范围(m/z)33~450。利用GC/MS工作站软件Masshunter自带NIST 11标准库自动检索各组分质谱数据,结合质谱裂解规律确定化学成分,利用面积归一化法计算出各组分相对峰面积百分比。
1.2.5 代谢差异物分析 色谱条件:使用BEH C18色谱柱(100 mm×2.1 mm,1.7 μm),柱温40 ℃、流速0.3 mL/min,进样量10 μL。最佳流动相由(A)0.1%甲酸水溶液和(B)0.1%甲酸乙腈溶液,梯度洗脱程序:0~6 min,5%~40%B;6~18 min,40%~85%B;18~18.5 min,85%~90%B;18.5~22 min,90%~90%B;22~22.5 min,90%~5%B;22.5~25 min,5%B。
质谱条件:采用ESI正离子(ESI+)模式进行扫描,离子源温度为100 ℃,脱溶剂气温度为350 ℃,脱溶气流量为800 L/h,锥气流量为50 L/h,毛细管电压为3 kV,采样锥电压为40 V,碰撞能量为6 eV,萃取锥电压为3 V,荷质比扫描范围50~1200 m/z。采用浓度为为200 ng/mL的亮氨酸-脑啡肽(556.2771 u)作为校正液,流速为30 μL/min。
1.3 数据处理及统计学分析
用SPSS 22.0进行Pearson相关性和方差分析;用Origin 8.5用于绘制图表;将UPLC/Q-TOF MS测定的原始数据经MassLynx 4.1进行数据处理,通过SIMCA 14.0和MetaboAnalyst v.5.0进行主成分分析、T检验分析和选择差异变量(P<0.05),与METLIIN、HMDB、KEGG、MASSBANK等生化数据库中的信息进行比较,对代谢差异进行分析。
2 结果与讨论
2.1 ACE抑制率的变化分析
已有研究表明,乳杆菌属的发酵乳中具有较高的ACE抑制活性,瑞士乳杆菌具有较强的蛋白水解系统,通过水解牛乳蛋白能够产生具有ACE抑制活性的肽,进一步水解产生氨基酸,满足了发酵乳制品的营养需求[16]。从图1可以看出,瑞士乳杆菌H11和副干酪乳杆菌Lc-01的发酵乳饮料在贮藏期间均显示出对ACE的抑制活性,这是由于益生菌的胞外蛋白酶将牛奶蛋白水解为寡肽,然后被胞内肽酶进一步水解为小肽和氨基酸[17]。而存在于肽的C端末位的疏水性氨基酸(例如Try、Phe、Trp、Ala、Ile、Val和Met)或带正电荷的氨基酸(例如Arg和Lys以及Pro)均具有较高的ACE抑制活性[18],例如,在C末端存在苯丙氨酸残基可能会增加肽的ACE抑制活性[10]。瑞士乳杆菌H11和副干酪乳杆菌Lc-01发酵乳饮料中的ACE抑制活性在贮藏过程中呈上升趋势,且瑞士乳杆菌H11比副干酪乳杆菌Lc-01显示出更高的ACE抑制活性,尤其是在贮藏21 d时瑞士乳杆菌H11表现出最高的ACE抑制活性(70.13%±2.83%),这可能是贮藏过程中pH和钙离子浓度的变化导致了蛋白质二次水解[17,19]。

图1 贮藏期间发酵乳饮料的ACE抑制率变化Fig.1 Change in ACE inhibition of fermented milk beverages during storage
2.2 VPP和IPP含量测定
瑞士乳杆菌通过水解牛乳中的蛋白质释放大量的肽,例如抗高血压肽VPP和IPP以及血管紧张素转化酶(ACE)抑制肽,其中ACE是控制高血压疾病的有效治疗靶点。从图2可知,在贮藏期间,瑞士乳杆菌H11的发酵乳饮料中的VPP和IPP含量显著高于Lc-01(P<0.05),且Lc-01的乳饮料在贮藏期间未检测到IPP且只检测到少量的VPP,这也是瑞士乳杆菌H11发酵乳饮料中ACE抑制活性高于Lc-01的原因。Chen等[6]证明了瑞士乳杆菌H9制备的发酵乳中具有86.4%±1.5%的体外ACE抑制活性,并且具有较高的VPP(2.409±0.229 μmol/L)和IPP(1.612±0.114 μmol/L)浓度。研究发现除了VPP和IPP外,通过水解蛋白质所产生的生物活性肽,例如αs1-CNf(90~94)(RYLGY)、αs1-CNf(143~149)(AYFYPEL)和αs2-CNf(89~95)(YQKFPQY)等多种活性肽也具有降血压的功能[20-21]。
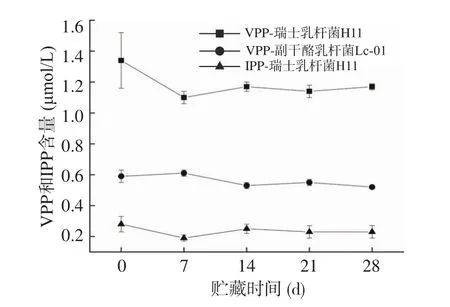
图2 贮藏期间发酵乳饮料中VPP和IPP含量的变化Fig.2 Changes in VPP and IPP content of fermented milk beverages during storage
2.3 基于SPME-GC-MS技术的风味物质分析
发酵乳饮料中具有多种挥发性风味化合物,其决定了发酵乳饮料的独特风味,采用SPME-GCMS技术对2组发酵乳饮料进行风味物质的测定,瑞士乳杆菌H11的发酵乳酸菌饮料中共检测54种挥发性风味物质,其中酮类物质6种,醇类物质13种,醛类物质7种,酸类物质10种,酯类物质8种,其他物质10种,对照样品中共检测73种挥发性风味物质,其中酮类物质13种,醇类物质16种,醛类物质6种,酸类物质12种,酯类物质6种,其他物质20种。为了研究2组样品之间的挥发性风味物质的差异以及贮藏时间对挥发性风味物质的影响,选择在贮藏过程中均含有且相对含量较高的7种风味物质,包括2-庚酮、2-壬醇、2-壬酮、2-十一烷酮、乙酸、2-十一烷醇和十一醛,各物质相对含量见表1。
由表1可以看出,瑞士乳杆菌H11和对照菌株Lc-01样品中的2-庚酮、2-壬酮相对含量均较高,在4 ℃贮藏28 d后分别为43.84%、12.39%和26.29%、15.80%。酮类物质产生是由于氨基酸的热降解、游离脂肪酸氧化和美拉德反应形成的,研究表明2-庚酮能够赋予发酵乳水果味[22],2-壬酮能赋予发酵乳果香、清香及奶油气息[23]。瑞士乳杆菌H11发酵乳饮料中的醇类和醛类物质含量在贮藏期间显著低于副干酪乳杆菌Lc-01(P<0.05),醛源自乳脂中不饱和脂肪酸的自动氧化,因为它们具有相对活跃的化学特性,很容易被还原成酸性化合物或醇类[24]。酸是甲基酮、醇、内酯和酯类物质的前体,对乳制品中的气味产生非常重要[25],在贮藏期间,瑞士乳杆菌H11发酵乳饮料中的己酸含量较高,其为乳制品提供类似于醋的强烈风味[26]。刘晓娇等[27]采用GC-MS技术对活性乳酸菌饮料的香气产物分析表明,主要的风味物质包括丁酸乙酯、己醛、2-庚酮、3-丁烯-1-醇、乙酸、2-壬酮等,其中2-庚酮和2-壬酮相对含量最高,分别为37.68%和17.86%,乙酸含量为4.97%,与本试验结果相近。上述结果可得知,L.helveticusH11能够赋予发酵乳饮料香气,具有较高的应用价值。
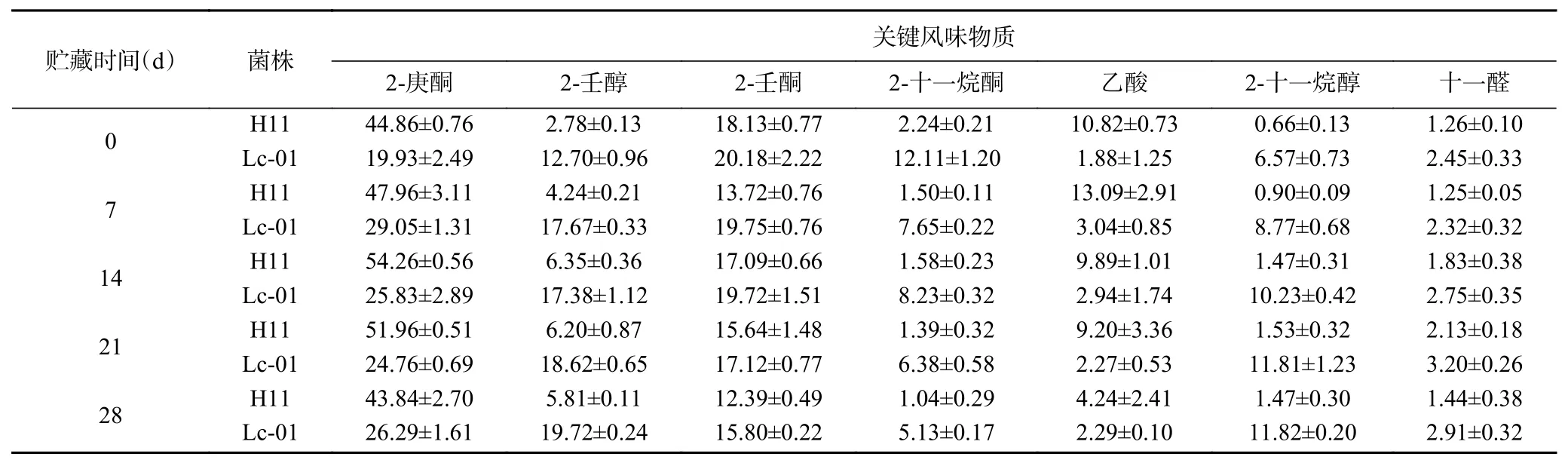
表1 贮藏期间发酵乳饮料中的风味物质的变化Table 1 Changes in flavour substances of fermented milk beverages during storage
2.4 基于代谢组学的代谢物差异分析
使用代谢组学分析贮藏过程中代谢产物的变化,从而预测产品的营养价值[28]。通过UPLC/QTOF MS/MS监测了瑞士乳杆菌H11和副干酪乳杆菌Lc-01两种发酵乳饮料在不同时间点的代谢谱变化,寻找两种发酵乳饮料之间的差异代谢物,在瑞士乳杆菌H11和Lc-01中分别检测到333和309种代谢物。为了可视化两组在不同时间点的代谢动力学,进行了PCA分析(图3)。两种发酵乳饮料分别形成两个区域,随着贮藏时间的延长,代谢谱发生了变化,于14 d后保持稳定。这表明在贮藏期间,瑞士乳杆菌H11和副干酪乳杆菌Lc-01仍具有代谢活性,使发酵乳饮料中的蛋白质发生二次水解,产生了更多的生物活性物质。
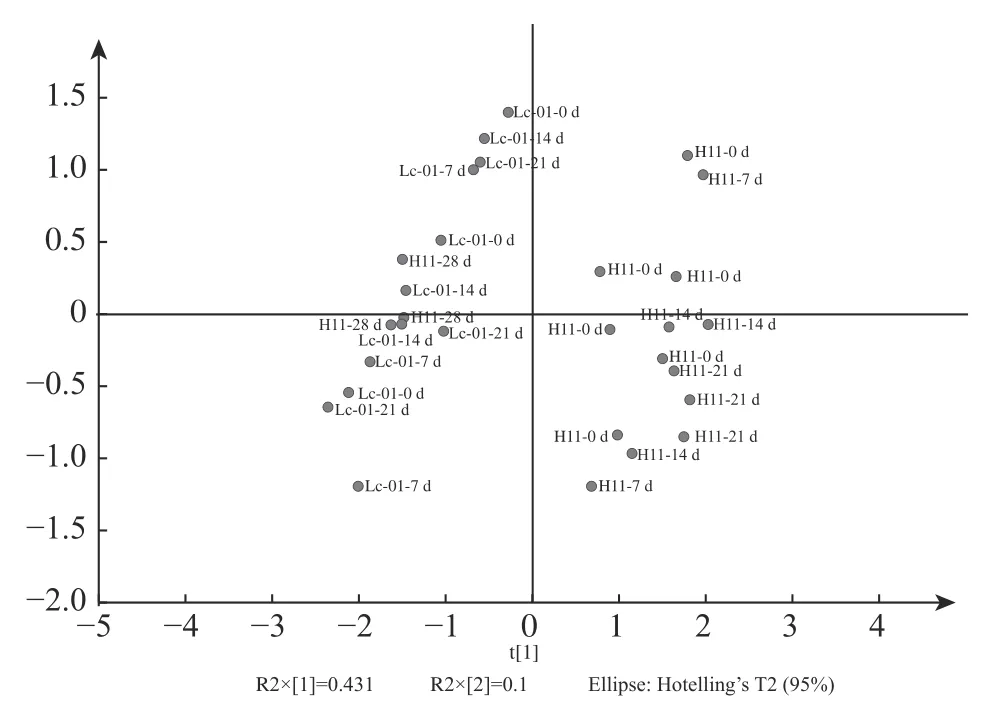
图3 贮藏期间两种发酵乳饮料代谢差异物的变化Fig.3 Changes in metabolic differentials of two fermented milk beverages during storage
瑞士乳杆菌通过细胞膜蛋白(CEP)将牛奶中的蛋白质分解为寡肽,再通过细胞内肽酶进一步水解成游离氨基酸,形成挥发性和非挥发性化合物,从而赋予发酵乳饮料特殊的风味[29]。当VIP值>1.5,Fold Change(差异倍数)>2和P<0.01时,在瑞士乳杆菌H11和对照发酵乳饮料中鉴定出16种代谢差异物,包括12种下调的代谢产物和4种上调的代谢产物(表2),主要为氨基酸、肽和有机酸;研究发现,通过LC-MS和GC-MS两种代谢组学技术对发酵乳制品进行研究,发现主要的代谢产物以氨基酸、短肽、脂类和有机酸为主[29]。通过对比KEGG数据库,发现瑞士乳杆菌H11和副干酪乳杆菌Lc-01之间的代谢差异物主要涉及氨基酸代谢、三羧酸循环以及磷酸戊糖途径,由于牛乳中的必需氨基酸成分较少,瑞士乳杆菌缺乏16种氨基酸合成基因,是高度营养缺陷型菌株,通过自身复杂的蛋白酶水解系统水解牛乳蛋白产生相应的氨基酸来维持生长发育,这可能是瑞士乳杆菌H11发酵乳饮料中四种游离氨基酸(4-氨基丁醛、L-脯氨酸、L-苏氨酸和L-苯丙氨酸)浓度在发酵和储存期间高于副干酪乳杆菌Lc-01的原因[21,30];另外,有研究发现由于瑞士乳杆菌自身复杂的水解系统,在发酵过程中能够有助于改善发酵产品的风味和质地,同时产生的生物活性肽有益于人体健康[31-32]。
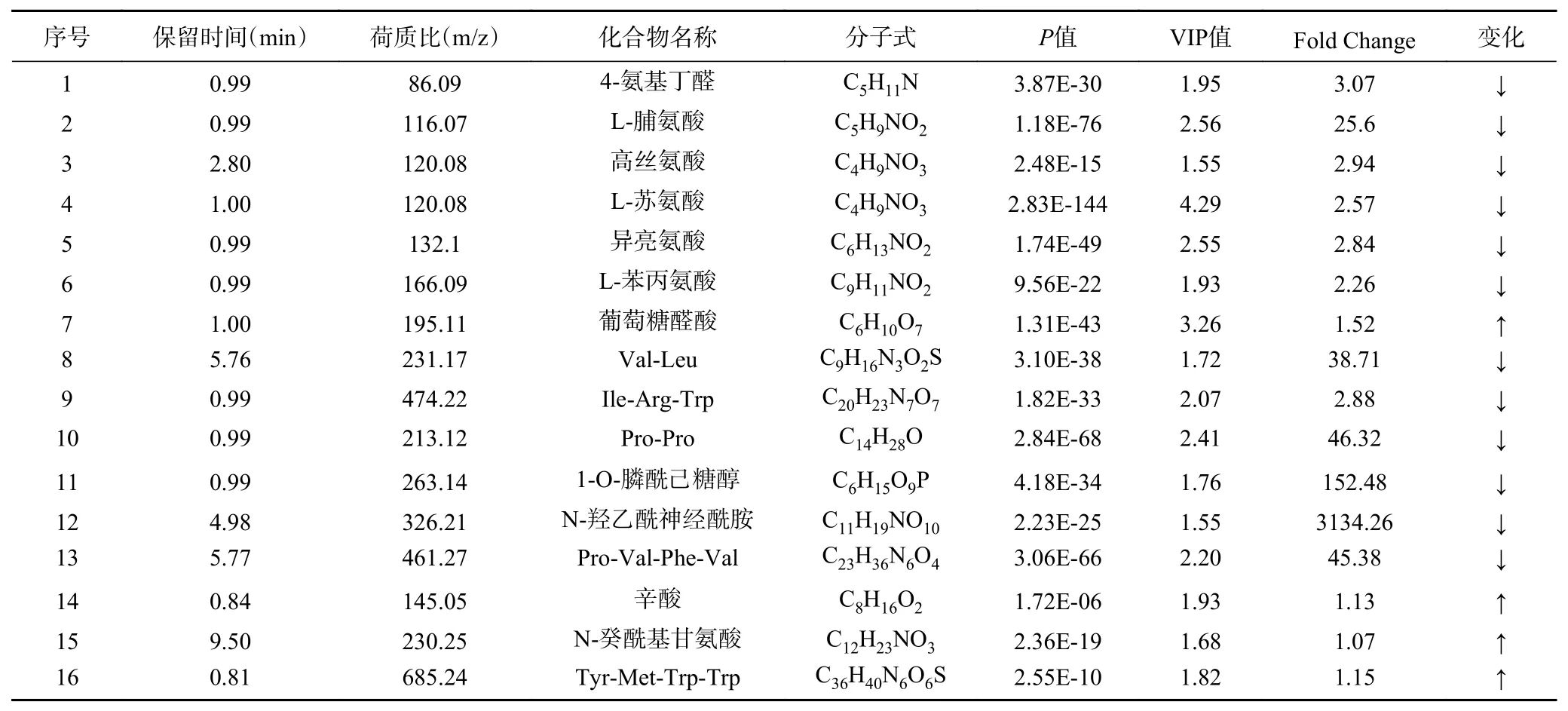
表2 瑞士乳杆菌H11和副干酪乳杆菌Lc-01发酵乳饮料在贮藏期间共有代谢产物变化Table 2 Changes in shared metabolites of L. helveticus H11 and L. paracasei Lc-01 fermented milk beverages during storage
3 结论
本文通过液相色谱、SPME-GC-MS和UPLCQ-TOF MS等代谢组学技术对瑞士乳杆菌H11和副干酪乳杆菌Lc-01发酵乳饮料的ACE抑制活性、代谢差异物和挥发性风味物质进行分析。结果表明,瑞士乳杆菌H11的ACE抑制活性比副干酪乳杆菌Lc-01高60%以上,ACE抑制肽VPP和IPP含量也显著高于Lc-01(P<0.05)。瑞士乳杆菌H11和副干酪乳杆菌Lc-01的主要代谢差异物为氨基酸、肽和有机酸,瑞士乳杆菌H11制备的活性乳饮料酸味更强且有奶油香气。综上所述,瑞士乳杆菌H11在降血压功能性发酵乳饮料的开发具有广阔的发展前景。
