GmSTK12基因转化大豆及对转化体生物量影响的研究
2021-11-11杨亚男黄荣梅
柏 锡,陈 云,杨 雪,焦 爽,杨亚男,黄荣梅
(东北农业大学生命科学学院,哈尔滨150030)
大豆作为粮食和经济作物在生产实践中应用广泛,作为常用豆制品、榨取豆油、提取蛋白质、酿造酱油原料,其体内所含氨基酸种类及比例与人体所需十分接近,易被消化吸收,是人类优质蛋白主要来源。由于环境影响,在遭受各种非生物胁迫时,其生长发育明显滞后,生理机能遭到破坏,导致大豆产量降低[1-3]。
丝氨酸/苏氨酸蛋白激酶是一类庞大的生物酶家族,参与蛋白质磷酸化途径,对真核生物生物学过程产生影响[4-5]。在植物中,蛋白激酶磷酸化过程被证实参与多种信号传导途径,目前已从拟南芥、小麦、玉米、水稻、番茄和大豆等植物中克隆出丝氨酸/苏氨酸蛋白激酶基因。蒋正宁等研究发现编码丝氨酸/苏氨酸蛋白激酶TaS/TK基因可提高小麦条锈病菌抗性[6];杨郁文等研究陆地棉发现编码丝苏氨酸蛋白激酶GhPK1基因参与盐胁迫反应[7];陶晓迎等研究发现盐藻丝氨酸/苏氨酸蛋白激酶基因DsSTPK显著提高盐藻耐盐性[8];Karl-Jo⁃sef等研究水稻发现编码丝氨酸/苏氨酸蛋白激酶SAPK4提高水稻耐盐性[9]。与此同时,Zhang等研究发现黄瓜中编码丝氨酸/苏氨酸蛋白激酶基因Cs⁃PID导致叶片形状发生改变,叶片形状是植物重要结构特征,受植物激素(特别是生长素)影响[10];晋育丹研究发现丝氨酸/苏氨酸蛋白激酶基因PID在调节拟南芥信号通路中,促进侧根细胞分裂和生长[11];魏述刚研究发现马铃薯StBL-SLG/PK是一个典型的G-type lectin丝氨酸/苏氨酸蛋白激酶,显著增强植株生长状况,如株高、根长、鲜重等[12]。
上述研究表明,丝氨酸/苏氨酸蛋白激酶类基因在植物抗逆、生长发育不同阶段发挥重要作用,但对大豆中丝氨酸/苏氨酸蛋白激酶类基因功能研究知之甚少。为此,本文将大豆中丝氨酸/苏氨酸蛋白激酶基因GmSTK12转化大豆,获得基因过量表达大豆材料,探究GmSTK12基因功能,阐明其对大豆植物生长发育影响,为培育大豆优良品种提供基因资源。
1 材料与方法
1.1 材料
本试验所用供试大豆品种为东农50;载体:pEASY-T3和pTF101.1;菌株:大肠杆菌DH5α和农 杆 菌EHA101;TransStart FastPfu DNA Poly⁃merase,反转录试剂盒(Easy Script One-Step gDNA Removal and cDNA Synthesis SuperMix)和荧光定量试剂盒(TransStart Top Green qPCR SuperMix)均购于北京全式金生物技术有限公司。
1.2 方法
1.2.1GmSTK12基因植物表达载体构建
根据Phytozome数据库提供的GmSTK12编码区序列,设计克隆基因引物,以大豆品种Williams82叶片cDNA为模板,应用TransStart FastPfu DNA Polymerase(北京全式金生物技术有限公司)作PCR扩增,获得GmSTK12基因CDS区,引物P1(见表1)。反应体系为50 μL,包含cDNA模板1 μL,上下游引物(10 μmol·L-1)各2.5 μL,FastPfu DNA Polymerase 0.5 μL,buffer 25 μL,ddH2O 18.5 μL。PCR反应条件为98℃预变性30 s;98℃变性10 s;60℃退火20 s;72℃延伸2 min,35个循环;72℃终延伸7 min。通过同源重组方法构建植物表达载体。

表1 引物序列Table 1 Primer sequences
1.2.2大豆GmSTK12基因过表达材料获得及鉴定
通过农杆菌介导的大豆子叶节遗传转化获得大豆过表达材料,提取阳性大豆植株和东农50叶片总DNA,以DNA为模板,使用Bar基因特异性引物作PCR扩增,引物P2(见表1)。根据扩增获得的GmSTK12基因序列,设计半定量和定量引物P3和P4(见表1)。以过表达大豆和东农50植株叶片180 ng·μL-1cDNA为模板,应用EasyTaqMix(康为世纪公司)作半定量PCR扩增。反应体系为20 μL,包含cDNA模板1 μL,上下游引物(10 μmol·L-1)各1 μL,EasyTaqMix 10 μL,ddH2O 7 μL。PCR反应条件为94℃预变性10 min;94℃变性30 s;60℃退火30 s;72℃延伸30 s,30个循环;72℃终延伸10 min。以过表达大豆和东农50植株叶片180 ng·μL-1cDNA为模板,Tua5为内参基因,引物P5(见表1)。应用全式金荧光定量试剂盒(Trans⁃Start TopGreen qPCR SuperMix)作实时定量PCR扩增。反应体系为15 μL,包含cDNA模板1 μL,上下游引物(10 μmol·L-1)各0.6 μL,2xTransStart Top⁃Green qPCR SuperMix 7.5 μL,ddH2O 5.3 μL。PCR反应条件为95℃预变性30 s;95℃变性10 s;60℃退火30 s,40个循环;溶解曲线,95℃15 s;65℃60 s;95℃1 s。每个反应设1次重复。利用公式2-ΔΔCt计算相对表达量。利用GraphPad Prism 5作图及显著性分析。
1.2.3 大豆GmSTK12基因生物学功能分析
挑选无病害、表面光滑、无破损东农50和过量表达GmSTK12基因大豆种子,待种子萌发后,将其放置于温度24℃,相对湿度60%,光照/黑暗为16 h/8 h人工气候室中培养。分别取过表达及野生型V3和V6期植株各30株,使用毫米位直尺测量过表达及野生型(Wild type,WT)相同部位大豆叶片长宽比、叶面积、根长,并统计主根及侧根数量。测定大豆生育期内植株鲜重、植株干重、根鲜重、根干重,使用清水冲洗植株表面杂物,滤纸吸干表面称量鲜重,105℃下30 min,80℃下烘干至恒重,称量干重。使用叶绿素测定仪(型号:TYS-B)测定植株叶片叶绿素含量,每个试验均使用3个生物学重复和3个技术重复。
1.2.4 大豆GmSTK12基因过表达植株农艺性状统计
对在人工气候室中种植东农50和过量表达GmSTK12基因T1和T2代植株作常规农艺性状考种。每个品种随机选取5株,收获后调查每株株高、主茎节数、有效分枝数、单株荚数、单株粒数、百粒重,取5株平均值作为各性状表型数据。
2 结果与分析
2.1 大豆GmSTK12基因过表达材料获得及鉴定
如图1A所示,通过同源重组方法构建pTF101.1-GmSTK12植物表达载体。利用农杆菌介导的大豆子叶节遗传转化法对东农50转化,对获得植株完全展开嫩叶作草铵膦涂抹阳性苗鉴定(草铵膦浓度为160 ng·μL-1)培养,获得GmSTK12基因过量表达大豆材料T1代阳性植株7株,分别命名为:FTDY-1、FTDY-4、FTDY-5、FTDY-11、FT⁃DY-13、FTDY-14、FTDY-15(见图1B、E)。
心脏瓣膜二尖瓣位于左房室口周缘,与乳头肌相连,其主要作用是维持心室血液的正常循环,防止左心室血液回流[6]。瓣叶、瓣环、乳头肌或左心室中任何一个部分出现结构改变,均可导致二尖瓣关闭不全,严重影响患者的健康安全[7-8]。目前,临床上普遍采用单纯的CABG治疗轻度二尖瓣关闭不全,有利于改善左心室体积。而对于重度二尖瓣关闭不全通常需要行CABG联合MVR治疗[9-10]。
将所获得T1和T2代植株取材,分别提取东农50和过量表达GmSTK12基因植株DNA鉴定标记基因Bar是否成功导入受体大豆。由图1C、F可看出,7份材料中均有Bar基因,且转基因材料中GmSTK12基因表达量高于受体品种东农50。同时,研究东农50和过量表达GmSTK12基因植株转录水平上测定目的基因是否表达,通过提取东农50和过量表达GmSTK12基因植株RNA作反转录cDNA,分析目的基因转录水平。由图1D、G可看出,qRT-PCR检测结果中T1代材料FTDY-11中GmSTK12基因表达量上调倍数达到300倍以上,T2代材料FTDY-11中GmSTK12基因表达量上调倍数也达到200倍以上。材料FTDY-1、FTDY-4、FT⁃DY-13、FTDY-14、FTDY-15中GmSTK12基因表达量上调倍数也超过100倍。表明已成功获得Gm⁃STK12基因过量表达的大豆材料,下一步将选取表达量较高的FTDY-4、FTDY-11、FTDY-15开展下一步试验,分析GmSTK12基因生物学功能。

图1 GmSTK12过量表达大豆植株分子鉴定Fig.1 Molecular identification of GmSTK12 overexpressing soybean plants
2.2 GmSTK12基因过量表达对大豆叶片长宽比和叶面积的影响
在温室内应季盆栽播种GmSTK12基因过量表达大豆材料FTDY-4、FTDY-11、FTDY-15和对照品种东农50各30株开展试验,在调查植株初期性状时并未发现大豆植株有明显差异,但当植株生长至V6期时发现过量表达GmSTK12基因植株叶面积发生明显改变,推测GmSTK12基因可能调控植株叶片大小,故调查植株V3、V4、V5、V6不同生长阶段叶长、叶宽及叶面积并作统计(V4、V5数据见图2)。由图3可知,植株生长VC至V3期时,两者叶片大小无显著差异(见图3A~C),且由图2数据显示,V4、V5期时植株叶片大小也无明显差异,而待植株生长至V6期时,过量表达GmSTK12基因植株叶片长宽比及叶面积开始高于对照品种。经统计分析发现,T1代过量表达植株叶面积约为对照品种叶面积2倍,且植株叶片长宽比差异显著,同时由图3可知,T2代植株叶面积及叶片长宽比与T1代一致,均显著高于对照品种,且T2代植株与T1代相比,表型更明显(见图3D~F)。结果表明,Gm⁃STK12基因可改变大豆植株叶片大小。
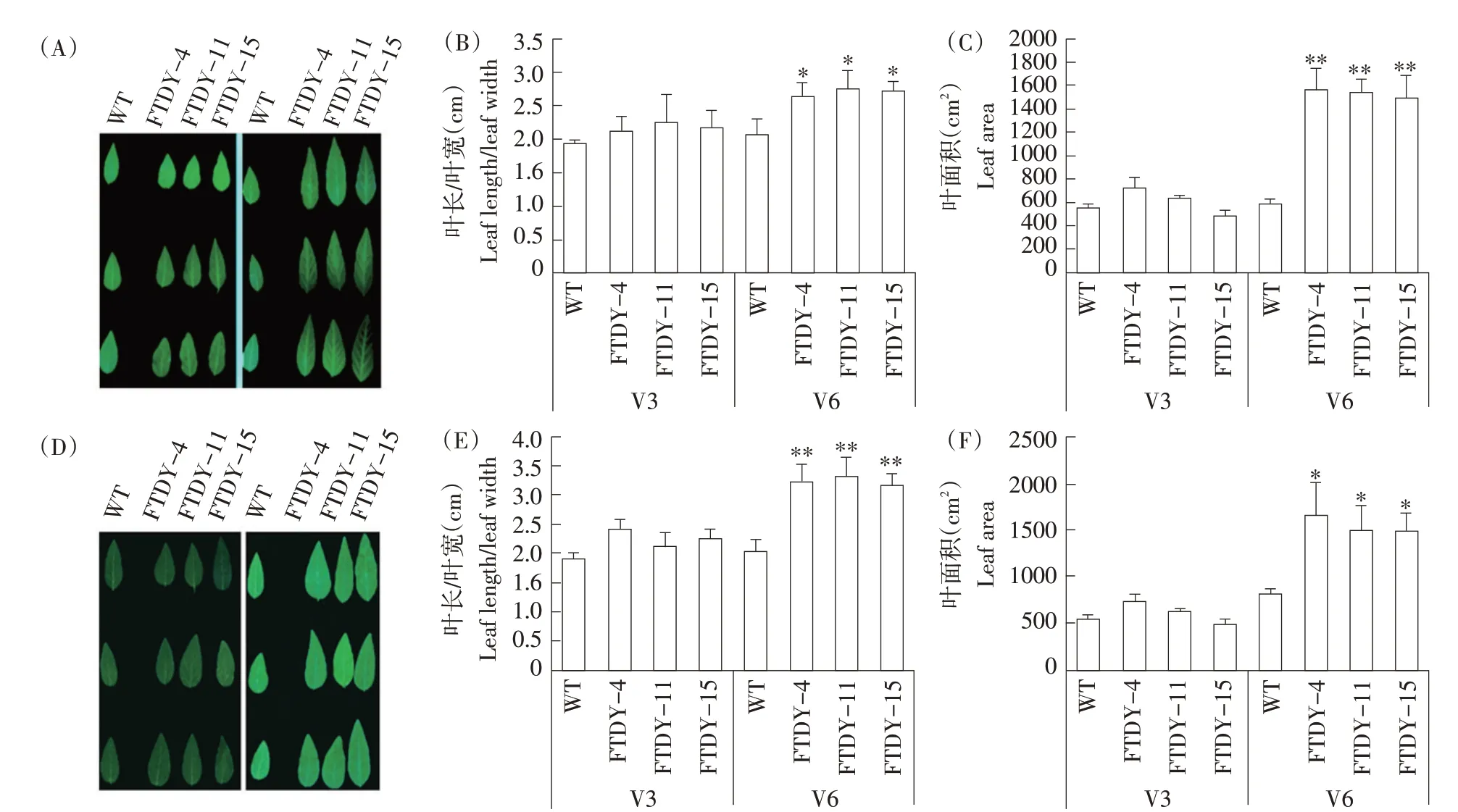
图3 大豆GmSTK12基因过量表达对T1、T2代植株叶片大小的影响Fig.3 Effects of soybean GmSTK12 gene overexpression on leaf size of T1 and T2 generation plants
2.3 GmSTK12基因过量表达对大豆根部的影响
为进一步分析GmSTK12基因功能,研究过量表达GmSTK12基因对大豆地下根部的影响,因此选择3份GmSTK12基因过量表达大豆材料FTDY-4、FTDY-11、FTDY-15和对照品种东农50植株数量各30株,分别测量其根长及侧根数量。
由图4A~C可看出,T1代GmSTK12基因过量表达植株根长与对照品种相比显著增长,约为对照品种植株根长2倍,表明过量表达GmSTK12基因植株根长增加,更利于吸收地下水分及矿质元素,植株生长旺盛;同时,过量表达植株侧根数量也明显高于对照品种植株,约为对照品种植株侧根数量1.5倍,由图4D~F可看出,T2代植株根长大于对照品种植株,侧根数量也多于对照品种,但与T1代植株相比,根长略长,而侧根数量相对较少。结果表明GmSTK12基因可使大豆植株根部形态发生改变。结果表明,过量表达GmSTK12基因根部生长情况较对照品种东农50更具优势。

图4 大豆GmSTK12基因过量表达对T1、T2代植株根长及侧根数量的影响Fig.4 Effects of soybean GmSTK12 gene overexpression on root length and lateral root number of T1 and T2 generation plants
2.4 GmSTK12基因过量表达对大豆植株地上及地下部分生物量的影响
研究发现,过量表达GmSTK12基因对大豆植株叶片大小及根部形态均有显著影响,为进一步确定过量表达GmSTK12基因对大豆植株地上及地下部分生物量是否也有影响,测定T1、T2代过量表达GmSTK12基因植株鲜重、干重、根干重、根鲜重。由图5A~H可看出,T1、T2代过量表达Gm⁃STK12基因植株鲜重和干重略高于对照品种,而根干重和根鲜重显著高于对照品种,均约为对照品种植株2倍。结果表明,过量表达GmSTK12基因增加大豆植株生物量,说明过量表达GmSTK12基因有助于植株营养代谢、生命活动旺盛。
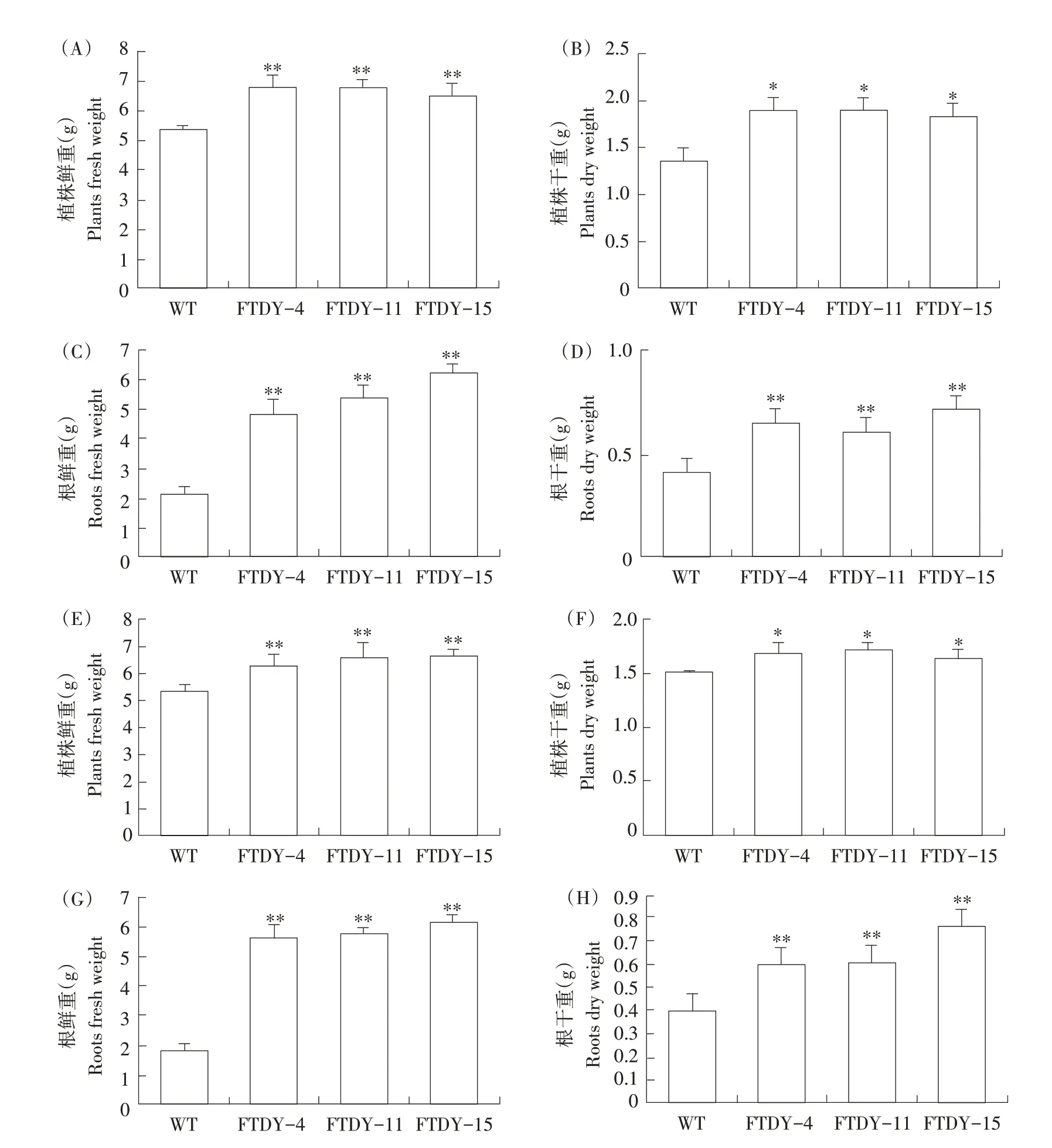
图5 GmSTK12基因过量表达对大豆植株生物量的影响Fig.5 Effects of GmSTK12 gene overexpression on soybean plant biomass
2.5 GmSTK12基因过量表达对大豆植株叶绿素含量的影响

图6 GmSTK12基因过量表达对大豆植株叶绿素含量的影响Fig.6 Effects of GmSTK12 gene overexpression on chlorophyll content of soybean plants
2.6 GmSTK12基因过量表达对大豆植株产量性状的影响
通过分析植株地上叶片面积及地下部分主根长度、侧根数量及叶绿素含量发现,过量表达Gm⁃STK12基因增强植株地上及地下生长发育状况,主根增长以及叶绿素含量增加优势使过量表达Gm⁃STK12基因植株地上部分叶面积增加,增强其光合作用合成有机物。因此进一步分析GmSTK12基因是否可通过增强大豆植株生长发育,进而提高大豆产量。
对农艺性状考种,由图7可看出,T2代过量表达GmSTK12基因植株主茎节数、有效分枝数、株高与对照品种相比无显著差异,但单株荚数、单株粒数、百粒重均显著高于对照品种。结果表明,GmSTK12基因通过调控主根长度增加以及地下部分生物量改变,提高对水分及矿质元素含量吸收,增强植物营养物质,且地上V6期叶片面积增加提高植物叶绿素含量,并有助于植物光合作用,增强营养生长期营养物质储备,以此改善生殖生长期地上部分对大豆产量的调控。

图7 GmSTK12基因过量表达对大豆植株产量性状的影响Fig.7 Effects of GmSTK12 gene overexpression on yield characters of soybean plants
3 讨论
丝/苏氨酸蛋白激酶基因是一类庞大信号分子,在作物生长发育中具有重要应用价值,其功能涉及作物生殖生长、株型、植株生物量及胁迫耐受性等方面。本研究克隆大豆中丝/苏氨酸蛋白激酶基因GmSTK12,利用农杆菌介导的大豆子叶节遗传转化法获得转基因株系,研究过量表达植株地上部分表型发现,过量表达GmSTK12基因植株叶面积明显大于对照品种植株(见图3C、F),与Zhang等在黄瓜中发现编码丝/苏氨酸蛋白激酶基因CsPID导致叶片增长结果一致[10],为探究其叶面积增加对植株光合作用影响,同时测定过量表达Gm⁃STK12基因植株和对照品种东农50叶片中叶绿素含量,过量表达植株叶绿素含量显著高于对照品种(见图6A、B),推测过量表达植株可能通过叶绿素含量增加吸收更多光能增强植株光合作用,合成有机物,使过量表达GmSTK12基因植株在营养生长期优于对照品种东农50;同时研究植株地下部分表型结果发现,其根长与侧根数量均显著优于野生型,研究发现水稻中编码丝/苏氨酸蛋白激酶基因OsASR6过表达品系使植株叶片增大,花序抽穗、角果和种子增多,且在根部生长中影响显著,与茎生物量增加1.7倍相比,根生物量增加4倍[13],与本试验结果一致,从而推测GmSTK12基因可能通过增加根部生物量(根干重,根鲜重等),增强植株吸水及保水能力,提高地下部分对水分及矿质元素的吸收,通过地下部分向地上部分运输营养物质,使植株地上部分也优于野生型植株,如植株鲜重和干重显著高于野生型(见图5A~H)。与此同时,过量表达植株侧根数量增多,根毛也随之增加,根毛是由根尖表皮的生毛细胞(Trichoblasts)发育而来,根毛形成显著增加根表皮细胞表面积,有助于根对土壤水分营养吸收和与土壤微生物的相互作用,促进植物生长发育[14-15]。通过统计植株生殖生长期产量性状发现,过量表达GmSTK12基因植株株系单株粒数、单株荚数及百粒重显著高于野生型,说明GmSTK12基因还具有提高大豆产量的功能,这一功能可能与植株根生长状况、叶面积大小及叶绿素含量均存在一定相关性,但GmSTK12基因对于大豆生长发育影响的具体机制还需进一步研究。
目前大豆中有关丝/苏氨酸蛋白激酶对植株生长发育的功能研究尚未明确,因此本研究以东农50为受体采用农杆菌介导的遗传转化法获得GmSTK12过表达植株,分析其全生育期表型及生物量。验证大豆中编码丝/苏氨酸蛋白激酶基因GmSTK12具有增加大豆植株生物量及产量的功能,为进一步探讨大豆中丝/苏氨酸蛋白激酶基因GmSTK12生理机制提供重要线索。
