纤维素酶枯草芽孢杆菌表达系统及其发酵黄芪的研究
2021-11-06伏春雨王亭亭刘小芳
赵 情,伏春雨,王亭亭,刘小芳,彭 军
(山东农业大学动物科技学院,山东 泰安 271018)
纤维素是自然界存在的天然可再生资源,是植物细胞壁的主要成分。纤维素酶是由内切葡聚糖酶、外切葡聚糖酶、β-葡聚糖酶3种酶组成的能够降解纤维素的一组酶系总称[1],在畜禽饲料和环境处理上发挥着重要的作用。在饲料中加入适量的纤维素酶,通过对植物细胞壁的破坏,不仅降低了饲料中的粗纤维含量,增强粗饲料的营养价值,而且更有利于被畜禽吸收和转化;纤维素酶还可将农作物秸秆和粪污中的纤维素分解成葡萄糖,在缓解环境压力的同时又可以产生大量的有机肥料。同时,天然菌株产纤维素酶能力差或活性低等问题,限制了这些菌株的应用范围并且难以大批量生产。近几年来,工程菌的出现渐渐避免了这些问题。目前已在大肠杆菌中成功表达多种纤维素酶基因[2],但由于大肠杆菌为条件致病性菌,所以不适合广泛应用于生产实践研究中。枯草芽孢杆菌是世界公认的属于安全级别的微生物,以它作为基因工程菌,能够更便利地构建饲用级表达系统,其基因工程产品更安全,而且它对于大肠杆菌来说具有易于培养的优势,而且质粒、噬菌体都可作为克隆载体,没有明显的密码子偏好性,转录翻译系统十分严密,可以避免形成错误折叠的蛋白质和包涵体,能够将蛋白直接分泌在培养基中,更有利于蛋白的表达[3]。
已有研究[4]发现,从地衣芽孢杆菌GXN151中筛选到3个表达纤维素酶活性的不重叠的独立克隆,其中Cel9A属于表达内切葡聚糖酶的一段基因,其结构前体的N末端第21-456氨基酸残基形成家族9糖基水解酶催化功能域,第480-565氨基酸为家族3碳水化合物结合组件,无明显的信号肽特征。该段基因在基因进化上有一定的保守性,这种基因结构对降解天然纤维素有非常重要的作用。
黄芪多糖是黄芪中最重要的天然有效成分。近几年的研究发现,黄芪多糖药理作用十分广泛,主要有增强免疫系统功能、抗炎症、抗氧化、抗病毒、抗肿瘤、降血糖及延缓衰老等作用,而且还可以作为免疫增强剂配合疫苗使用,增强免疫效果[5]。作为饲料添加剂使用,可以促进动物生长。因此,对多糖的提取和活性研究成为了新的热点,传统多糖的提取方法有水煎煮法、碱水提取法、碱醇提取法、超声波提取法、微波提取法等。生物酶法提取黄芪中的多糖是在传统的溶剂提取方法的基础上,根据黄芪细胞壁的结构组成,利用酶反应所具有的高度专一性等特点,将细胞壁的组成成分水解或是降解,破坏细胞壁的结构,使有效成分充分暴露出来,溶解、混悬于溶剂中,从而达到提高黄芪多糖提取率的目的。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒 从NCBI查找纤维素酶编码基因Cel9A的序列,基因合成部分由生工生物工程有限公司完成;枯草芽抱杆菌WB800N用作枯草杆菌表达宿主,购于北京华越洋生物。
1.1.2 主要试剂 限制型内切酶、T4 DNA连接酶,购自于TaKaRa公司;引物为上海瑞博生物科技有限公司合成;低分子量蛋白质标准购自上海实生公司;卡那霉素、氯霉素购自上海生工生物公司;胶回收试剂盒购自上海华舜生物公司;SDS-PAGE试剂盒购于上海雅酶生物科技有限公司;Western Blot一抗、二抗购自康为世纪生物科技有限公司;DAB染色液;CMC营养琼脂;革兰氏碘染色液(2g碘化钾和1g碘溶于300 ml蒸馏水);发酵培养基。
1.2 方法
1.2.1 CeL9A目的基因 目前研究纤维素酶较多的细菌主要是纤维素粘菌属、生孢纤维粘菌属、纤维杆菌属和芽孢杆菌属。真菌则多是木霉属、曲霉属、青霉素、根霉属、漆斑霉属等,真菌在产纤维素酶的过程中,大多会产生霉菌毒素,而且不易去除,如果大量应用于饲料中会给家畜的健康带来损害。所以本研究选取芽孢杆菌属的地衣芽孢杆菌中的一段纤维素酶基因Cel9A作为扩增目的基因。
查阅数据库获得地衣芽孢杆菌纤维素酶Cel9A基因序列(GenBank:AY445620)[4],分别在序列上游和下游添加限制性内切酶位点BamHI和SmaI,在下游终止子后添加组氨酸标签:CATCATCACCATCACCAT。
1.2.2 构建重组质粒 选取地衣芽孢杆菌中的Cel9A基因作为扩增目的基因,基因合成由生工生物工程有限公司完成,命名为PUC57-Cel9A。然后,双酶切克隆质粒PUC57-Cel9A、表达质粒PHT43并进行胶回收(表1),将两胶回收片段用T4DNA连接酶于16 ℃ 过夜连接(表2)。

表1 双酶切反应体系
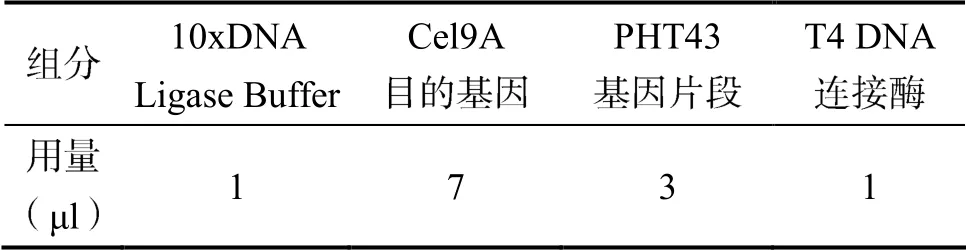
表2 连接反应体系
1.2.3 构建重组菌 采用传统的CaCl2化学转化方法[6]将PHT43-Cel9A转化到B.subtilisWB800N感受态菌株中,将转化菌放入含有Amp抗生素的LB肉汤中过夜培养。对浑浊的菌液提质粒并用PCR进行验证,引物序列为:PHT43-R:5'-CGATATTAGCCTCGTATGTT-3';PHT43-F: 5'- -3',反应条件见表3。

表3 PCR反应体系
1.2.4 重组菌诱导表达 将重组菌1:100扩大培养后在LB营养肉汤中培养至OD600=0.8,加入诱导剂IPTG,37 ℃、200 rpm进行诱导。将诱导好的菌液离心取上清,样品与buffer按照1:4的比例混合后沸水浴10min,SDS-PAGE检测蛋白表达。
Western Blot鉴定蛋白的反应原性,其中一抗为鼠抗His单克隆抗体,二抗是羊抗鼠单克隆抗体,两种抗体根据说明书用抗体稀释液进行稀释使用。将经过聚丙烯酰胺凝胶电泳分离的蛋白质样品转移到固相载体上,以固相载体上的蛋白质或多肽作为抗原,与对应的抗体起免疫反应,再与第二抗体进行反应,经过底物显色以检测电泳分离的特异性目的基因表达的蛋白成分。
1.2.5 生长曲线及质粒稳定性 将重组菌株接种于LB液体培养基中,37 ℃,200 rpm振荡培养,每隔2 h使用分光光度计进行OD600值的测定,并绘制生长曲线。
将过夜培养的重组菌按1:100的比例转接到LB液体培养基中培养12 h,之后再按照1:100的比例转接到新的LB液体培养基中,每转接一次作为一代,按照此方法连续进行12次转接后,用PCR方法对重组菌质粒的稳定性进行检测[7]。
1.2.6 重组菌的酶学性质 (1)DNS法测定重组菌的酶活性:纤维素酶能将羧甲基纤维素降解成寡糖和单糖。具有还原性末端的寡糖和有还原基团的单糖在沸水浴条件下可以与DNS试剂发生显色反应。反应液颜色的强度与酶解产生的还原糖量成正比,而还原糖的生成量又与反应液中纤维素酶的活力成正比。因此,通过分光比色测定反应液颜色的强度,可以计算反应液中纤维素酶的活力。
试验原理:还原糖与3,5-二硝基水杨酸进行反应,被还原成棕红色的氨基化合物,在一定范围内还原糖的生成量与反应液的浓度是成正比例关系的,利用分光光度计在540nm处测得吸光度值,得出还原糖的量。
葡萄糖标准曲线的绘制:精确称取1.0 g葡萄糖,完全溶解于100 ml 0.1 mol/L的乙酸-乙酸钠中。吸取不同体积的葡萄糖溶液以配制成不同浓度的葡萄糖标准溶液。吸取2.0 ml的葡萄糖标准溶液(表4),加入到刻度试管,再加入2.0 ml水和5.0 mlDNS试剂。电磁振荡3 s后沸水浴加热5 min,用自来水冷却至室温后定容至25 ml。以葡萄糖浓度(mg/ml)为横坐标,以对应的吸光度OD540值为纵坐标,绘制葡萄糖标准曲线。
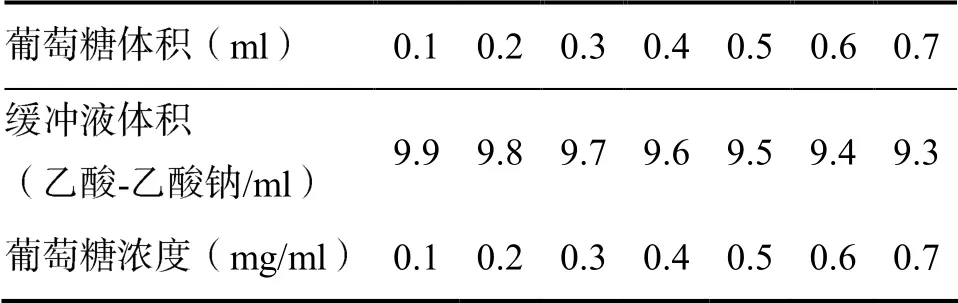
表4 葡萄糖标准溶液配置
采用DNS法对产纤维素酶的重组菌进行酶活性检测[8],以羧甲基纤维素钠(CMC-Na)为底物,取粗酶液2 ml,再加入2 ml的CMC-Na(已经过37 ℃ 平衡),电磁振荡3 s,37 ℃ 保温30 min,再加入5.0 ml的DNS试剂,电磁振荡3 s,以终止酶解反应,沸水浴加热5 min后用自来水冷却至室温,加水定容至25 ml,电磁振荡3 s。在特定波长(540 nm)下测定CMC酶活。CMC酶活单位为每分钟每毫升酶液催化纤维素水解生成1.0 µg葡萄糖所需的酶量为一个酶活力单位U,即1 U/ml。
(2)重组菌的酶活性检测-透明圈:将10 µl重组菌(已在LB肉汤中复苏)放入打孔的CMC营养琼脂中,37 ℃ 倒置培养24 h。用革兰氏碘染色液[9]染色3~5 min后观察透明圈。
1.2.7 重组菌的分解发酵作用 将诱导好的菌液以3 % 的接种量接种到含有黄芪粉的发酵培养基中,37 ℃,200 rpm震荡培养。测定不同培养时间对应粗多糖的含量(苯酚-硫酸分光光度测定法)[10]。
2 结果
2.1 重组质粒的构建
对克隆质粒PUC57-Cel9A分别用限制性内切酶BamH I和Sma I进行双酶切,电泳检测酶切产物。对1930bp处的电泳条带进行胶回收后与表达载体PHT43进行连接,经PCR鉴定为阳性,结果如图1。
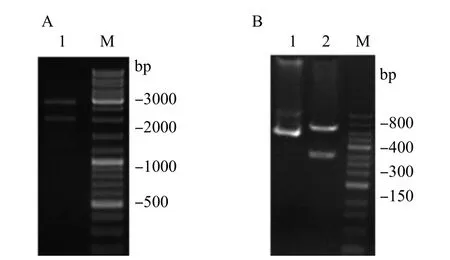
图1 A:PUC57-Cel9A质粒鉴定;B:双酶切鉴定
2.2 重组菌的构建
通过基因重组技术成功构建了表达纤维素酶的重组芽孢杆菌WB800N/PHT43-Cel9A。基因Cel9A经测序鉴定成功克隆到启动子和信号肽下游,引物为PHT43的通用引物,PCR鉴定结果如图2。

图2 WB800N/PHT43-Cel9A通用引物PCR鉴定
2.3 重组菌的诱导表达
2.3.1 聚丙烯酰胺凝胶电泳(SDS-PAGE) 用IPTG对重组菌进行诱导,对不同时间段的诱导菌进行取样处理。SDS-PAGE检测蛋白表达情况,其中第一条泳道为Marker,第二条泳道为空质粒菌作为对照,第三、四、五条泳道分别为诱导8 h、10 h、12 h的菌液。可以明显的看出重组枯草芽孢杆菌在诱导8h后表达分泌量较高。
2.3.2 生长曲线及质粒稳定性测定 将培养在LB肉汤中的重组菌每隔4 h进行取样,在紫外分光光度计下测定其OD600的值,并绘制生长曲线,如图3。重组菌在16~24 h时间段内生长较快,并在24 h时OD600达到最大值,然后随着时间的增长OD600逐渐下降。
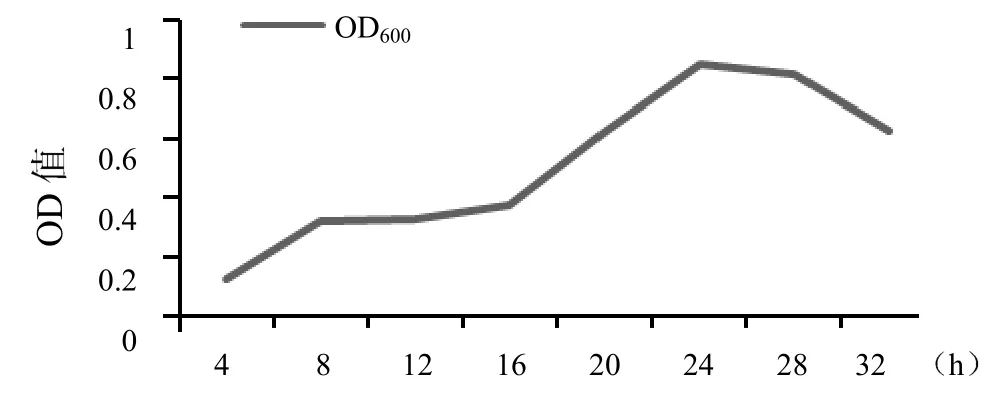
图3 重组菌生长曲线
将重组菌放入不含抗生素的LB肉汤中培养,每12 h进行转接一次,12 h算作一代,并取第2代、4代、6代、8代、10代、12代的菌液提质粒并进行PCR鉴定,结果显示经过12代后重组菌的质粒稳定,如图4。
图4 重组菌质粒稳定性
2.3.3 重组菌的酶学性质 (1)DNS法测定酶活性:葡萄糖标准曲线方程是Y=3.3821X-0.2973(R2=0.9777),通过标准葡萄糖曲线算出重组枯草芽孢杆菌的酶活力为43U/ml。试验中所用的葡萄糖标准曲线绘制结果如图5。
图5 葡萄糖标准曲线
(2)酶活性测定-透明圈:革兰氏碘与纤维素形成蓝黑色复合物,但不与水解纤维素形成,在产生纤维素酶的微生物菌落周围形成清晰且明显的区域。对已培养24 h的CMC板染色3~5 min,观察透明圈,将重组枯草芽孢杆菌与原枯草芽孢杆菌的透明圈进行对比,明显的看出重组枯草芽孢杆菌的透明圈较大,具体结果如图6。
图6 酶活性鉴定(透明圈)
2.3.4 发酵分解作用 纤维素酶对天然植物多糖的细胞壁具有分解作用,所以利用该作用提取多糖,将重组枯草芽孢杆菌与地衣芽孢杆菌分别发酵黄芪多糖。重组枯草芽孢杆菌在12 h时多糖的提取率较高,达到5.2 %,而地衣芽孢杆菌相应的多糖提取率明显低于重组枯草芽孢杆菌,结果如图7。
图7 纤维素酶重组枯草芽孢杆菌、地衣芽孢杆菌 发酵黄芪相应的多糖产量
3 讨论
由于纤维素酶和纤维素酶生产菌能将一些粗饲料如玉米芯、麦糠、稻草和麦秆等转化为脂肪、菌体蛋白和糖等其他一些营养物质,所以它在饲料加工业中的应用十分广泛。纤维素酶发酵分解后不仅降低了饲料中的粗纤维含量,增强粗饲料的营养价值,而且对补充动物的内源纤维素酶、消除抗营养因子、促进养分的消化吸收有重要作用。
黄芪是人们经常食用的中药材,民间流传“常喝黄芪汤,防病保健康”的顺口溜,由此可见,对于黄芪人们经常采用水煮的方式来提取其中的有效成分。近年来,随着中药材的广泛应用,国内外学者开始对黄芪的化学成分、药理活性感兴趣并做了大量的研究,发现黄芪中含有多糖、蛋白质、氨基酸、黄酮等成分,其中多糖成分是主要的生物活性成分之一[11]。黄芪多糖主要由葡萄糖、鼠李糖、阿拉伯糖和半乳糖组成,其诸多功能中以免疫调节功能最为突出[12]。同时,由于人们对化学合成药物毒副作用及残留的认识提高,黄芪多糖以兽药原料、针剂、口服液、饲料添加剂等多种形式,被广泛地应用于畜禽的保健和临床治疗中[13]。
目前,黄芪多糖提取主要采用传统的水提醇沉、有机溶剂、超声波辅助提取法,但黄芪多糖的提取率普遍较低,且有机溶剂用量大[14]。酶解提取主要是利用纤维素酶破坏细胞壁的致密结构,加速药用有效成分溶出,提高得率,由于其操作简单、条件较温和,不需要大型设备,在成分结构与活性保持方面具有一定的优势,正逐渐被天然药物工作者所采用。
采用纤维素酶提取黄芪多糖,进而提高多糖得率。张嘉怡等[15]通过正交试验得出纤维素酶提取工艺的优化条件:料液比1:12(g/ml),处理时间90 min,处理温度100 ℃,水提3次,最终得出多糖提取率4.76 %,相较于传统热水浸提法提高了54.05 %。魏凤玉等[16]采用纤维素酶解法提取黄芪多糖,采用扫描电镜观察其酶解后的黄芪残渣表面结构,发现纤维素酶只破坏细胞壁,降低内部多糖扩散的阻力,因此提高了黄芪多糖的传质速率和有效扩散系数。闫巧娟等[17]通过对黄芪原糖、传统工艺药渣和酶处理药渣进行扫描电镜观察,结果表明,纤维素酶处理后的药渣,其内部附着物较少,网状结构较清晰。
为了适应市场对纤维素酶的产量要求,不少学者已开始研究创制纤维素酶生产菌。Mingardon[18]等使来源于真菌里氏木霉的8个纤维素酶基因cbh1、cbh2、eg1、eg2、eg3、eg4、eg5、bgl2,成功在大肠杆菌中获得表达。
本研究利用基因重组技术构建枯草芽孢杆菌纤维素酶表达系统,显著提高了纤维素酶的产量和黄芪多糖的提取效率,降低了其使用成本,为黄芪以及农产品秸秆等天然植物多糖在畜牧业中的大量推广利用提供技术支撑。
