羰基还原酶的克隆、异源表达及其在(R)-2-羟基-4-苯基丁酸乙酯合成中的应用
2021-11-05庞凯南王鑫情朱佳妮来婉琪刘锐华邵飞鸿黄一帆
庞凯南,王鑫情,朱佳妮,来婉琪,刘锐华,邵飞鸿,黄一帆,罗 希
(台州学院 生物质资源研究所,浙江 台州 318000)
随着人们生活水平的提高,心血管疾病的发病率和死亡率逐年上升,已成为危害人类健康的头号杀手,其中高血压是导致患者死亡的主要原因之一[1]。普利类药物,如卡托普利、依那普利、贝那普利、赖诺普利和雷米普利等,能抑制体内血管紧张素转换酶活性,切断肾素-血管紧张素-醛固酮系统,阻断血管紧张素Ⅱ的形成,从而达到扩张血管、降低血压的作用,是目前市场上治疗高血压的主要药物[2-3]。(R)-2-羟基-4-苯基丁酸乙酯(R-HPBE)是制备普利类药物的关键手性中间体[4]。
近年来,一系列化学和生物过程被用于合成R-HPBE。化学法多采用过渡金属催化剂和多步溶剂萃取,产物分离纯化困难、反应条件苛刻、产物光学纯度差。而生物合成法具有立体选择性高、环境友好、反应条件温和等突出优势,更符合绿色合成要求,因而受到了广泛的关注[5]。微生物来源广泛、生长周期短且具有丰富的多样性,是廉价易得的生物催化剂,可催化2-氧代-4-苯基丁酸乙酯(OPBE)不对称还原制备R-HPBE。但微生物细胞内多种不同立体选择性还原酶共存且酶表达量低,使得产物光学纯度不高,产物累积浓度也普遍较低[6]。大肠杆菌(Escherichiacoli)易于操控,重组蛋白表达量约占细胞内总表达量的55%,常用作异源基因表达宿主[7]。已有多种不同来源的羰基还原酶在大肠杆菌中表达,高效催化OPBE不对称还原,获得光学纯R-HPBE。如Ni等[8]将来源于枯草芽孢杆菌(Bacillussubtilis)的羰基还原酶IolS与葡萄糖脱氢酶共表达,在水/正辛醇两相反应体系中催化OPBE不对称还原,获得光学纯R-HPBE,时空产率为660 g/(L·d)。Shen等[9]将来源于光滑假丝酵母(Candidaglabrata)的羰基还原酶CgKR2与葡萄糖脱氢酶耦联,在水相中催化合成R-HPBE,时空产率达到700 g/(L·d),对映体过量值(e.e.)>99%。
虽然,到目前为止已有一些野生型微生物和羰基还原酶重组菌被应用于不对称合成R-HPBE,发现新型的、有潜力的生物催化剂依然是手性中间体合成研究的重要课题。新的生物催化剂的发现,不仅可为包括R-HPBE在内的手性中间体绿色制造提供更多选择方案,也可为应用蛋白质工程改造酶的催化功能和特性提供材料,以加深酶结构-功能关系的理解,满足工业化生产的需要。因此,本研究中,笔者筛选一株具有催化OPBE不对称还原制备R-HPBE活性的野生型微生物菌株,并通过基因挖掘获得关键酶基因,构建重组大肠杆菌并异源表达,用于全细胞催化合成R-HPBE,考察其反应工艺,为规模化生产提供数据。
1 材料与方法
1.1 材料与试剂
胰蛋白胨和酵母粉,Oxoid(英国);分子克隆和重组大肠杆菌构建所用酶和试剂盒,生工生物工程(上海)股份有限公司或TaKaRa公司;2-氧代-4-苯基丁酸乙酯、(R)-2-羟基-4-苯基丁酸乙酯等,百灵威科技有限公司;其他试剂均为市售分析纯。
野生菌筛选所需土样取自于浙江省内农田、果园、湖泊及化工厂等环境。
1.2 野生菌的筛选
称取不同土样各0.5 g,加无菌水10 mL悬浮,放至30 ℃恒温摇床中孵育1 h制备土壤菌悬液。吸取菌悬液1 mL分别加入LB培养基和豆芽汁培养基中,分别在30和28 ℃恒温摇床中培养至培养液浑浊。取1 mL富集培养液,梯度稀释。分别吸取106、107和108倍稀释的富集培养液50 μL,加入琼脂平板,玻璃涂布棒涂布均匀,并放至恒温培养箱培养(LB琼脂平板30 ℃,豆芽汁平板28 ℃)至长出显著单菌落。挑取单菌落,分别接种至新鲜LB培养基(30 ℃、180 r/min培养24 h),或豆芽汁培养基(28 ℃、180 r/min培养48 h)。在8 000 r/min、4 ℃条件下离心10 min,收集细胞并置于4 ℃冰箱备用。
催化反应在50 mL摇瓶中进行,称取0.05 g OPBE、0.05 g葡萄糖至装有不同菌种细胞的离心管中,加入10 mL磷酸缓冲液(100 mmol/L,pH7.0),充分悬浮后,于30 ℃、200 r/min摇床中反应12 h,取样后以2,4-二硝基苯肼显色法初筛[10],确定活性菌株后再采用气相色谱法复筛。
1.3 菌种鉴定
将目标菌株培养物适当稀释后涂布于富集培养基固体平板上,培养至产生单菌落后观察形态。电子显微镜观察细胞形态。16S、18S rDNA 的分子生物学鉴定:提取活性菌基因组DNA,送生工生物工程(上海)股份有限公司测序鉴定。
1.4 羰基还原酶的克隆和重组基因工程菌的构建
以羰基还原酶(CR)氨基酸序列(GenBank:PTN20075.1)为探针,通过NCBI数据库中BLAST功能进行比对分析,找出来源于Kazachstanianaganishii菌种的相似序列。以K.naganishii基因组为模板,CR-F 5’-CCATGGGCATGTCCGTTT TTATCTCAGG-3’(NcoⅠ),CR-R 5’-CTCGAGTAGGTTCCCTTCGTGTTTC-3’(XhoⅠ)为引物进行PCR反应,克隆酶基因。PCR反应体系(总体积100 μL):10×Taq DNA Polymerase Buffer 10 μL(Mg2+),引物CR-F和CR-R各1 μL(50 μmol/L),dNTP mixture 1 μL(10 mmol/L),基因组DNA 1 μL,Taq DNA Polymerase 1 μL,去离子水85 μL。PCR程序:94 ℃预变性5 min,经过35个循环(94 ℃ 0.5 min,56 ℃ 0.5 min,72 ℃ 1.5 min),72 ℃延伸10 min。琼脂糖凝胶电泳鉴定后,胶回收目标酶DNA,双酶切获得黏性末端插入表达载体pET-28a,重组质粒用热击法转入E.coliBL21(DE3)感受态细胞,构建羰基还原酶基因工程菌。
1.5 羰基还原酶重组菌诱导表达条件优化
接种E.coli/pET-28a-kncr至含有50 μg/mL卡那霉素的LB液体培养基,37 ℃培养10 h,转接(4%,体积分数)至有50 μg/mL卡那霉素的LB液体培养基中,37 ℃培养至菌液OD600为0.8,加入乳糖,在28 ℃、150 r/min下诱导,分别对诱导剂乳糖的用量(2、4、6、8、10和12 g/L)和诱导时间(2、4、6、8和10 h)进行优化。离心收集菌体。
生物量测定:取诱导完成的发酵液1 mL,稀释10倍,在600 nm下测定吸光值。细胞超声破碎后离心,上清液进行SDS-PAGE凝胶电泳分析。
酶活测定反应体系(200 μL):0.25 mmol/L NADH,1 mmol/L OPBE,100 μL粗酶液溶于磷酸钾缓冲液(100 mmol/L,pH7.0),用酶标仪测定OD340的变化值。酶活定义:在30 ℃、pH7.0条件下,反应消耗1 μmol NADH所需的酶量定义为一个酶活单位(1 U)。
1.6 全细胞催化反应条件优化
羰基还原酶重组菌细胞,OPBE和葡萄糖溶于10 mL磷酸钾缓冲液(100 mmol/L)。分别对pH(6.0、6.5、7.0、7.5和8.0)、温度(25、30、35、40和45 ℃)、葡萄糖(5、10、15、20、25和30 g/L)、细胞用量(0.5、1.0、1.5、2.0和2.5 g)、底物质量浓度(5、10、15、20、25和30 g/L)进行优化,于200 r/min反应30 min后取样,乙酸乙酯萃取后气相色谱检测。
1.7 全细胞催化反应时间进程
催化反应体系50 mL,在筛选后的最优条件下进行,磁力搅拌转速设为500 r/min,流加1 mol/L Na2CO3溶液维持反应液pH恒定。每1 h取样,气相检测。
1.8 检测方法
HPBE浓度和R-HPBE对映体过量值(e.e.)的测定参照文献[11]的方法。
2 结果与讨论
2.1 菌种筛选与鉴定
微生物是酶的天然宝库,与基于数据库的基因挖掘法相比,从野生型微生物体内获得所需新酶的几率更高。选取农田、果园、湖泊和化工厂等腐殖质较多、微生物物种丰富的土壤作为土样,经富集培养及分离纯化,共获得236株野生菌。通过2,4-二硝基苯肼显色法初筛和气相色谱复筛,获得19株形态各异的野生型菌种。其中,编号为DY99的菌株能催化不对称还原OPBE反应,获得R-HPBE 2.47 g/L,显示了较优催化活力,且产物e.e.也较高,达到91.3%(图1(a))。在豆芽汁固体培养平板上,DY99菌落呈白色、圆球形、凸起、表面光滑、边缘整齐、不透明状(图1(b))。经显微镜观察,DY99细胞呈卵圆形,少数细胞正进行出芽生殖(图1(c)),初步判定为酵母细胞。为进一步确定该菌种属,将PCR扩增获得的18S rDNA测序获得序列并输入GenBank数据库中,利用BLAST软件将该序列与NCBI收录的DNA序列进行比对,并通过系统进化树分析,结果见图2。由图2可知:菌株DY99与Kazachstanianaganishii同源性最高(100%,18S rDNA序列GenBank:AB016512.1)。结合形态特征,DY99确定为Kazachstanianaganishii。
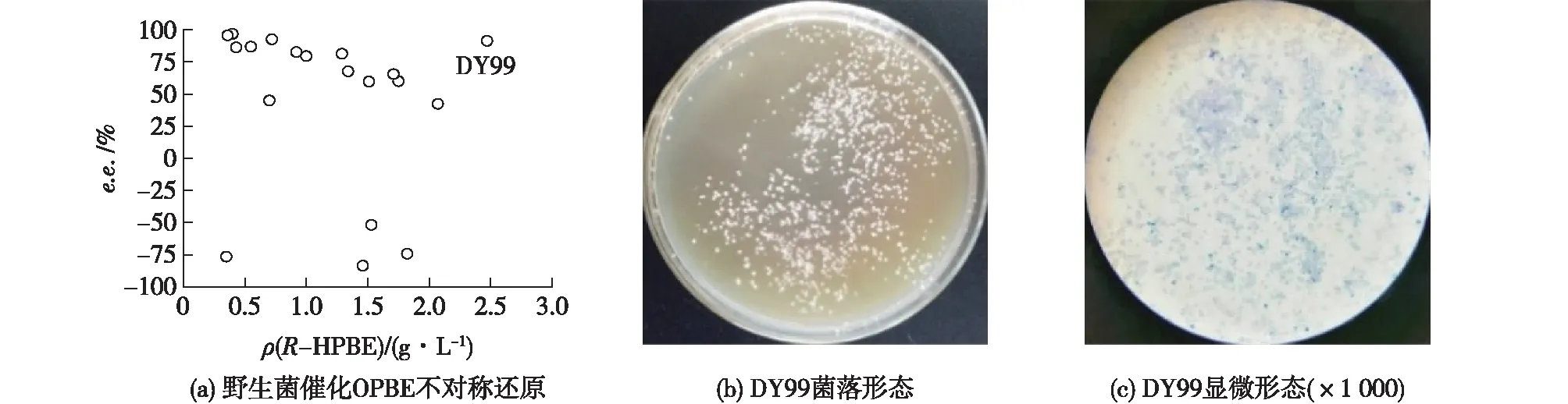
图1 菌种筛选
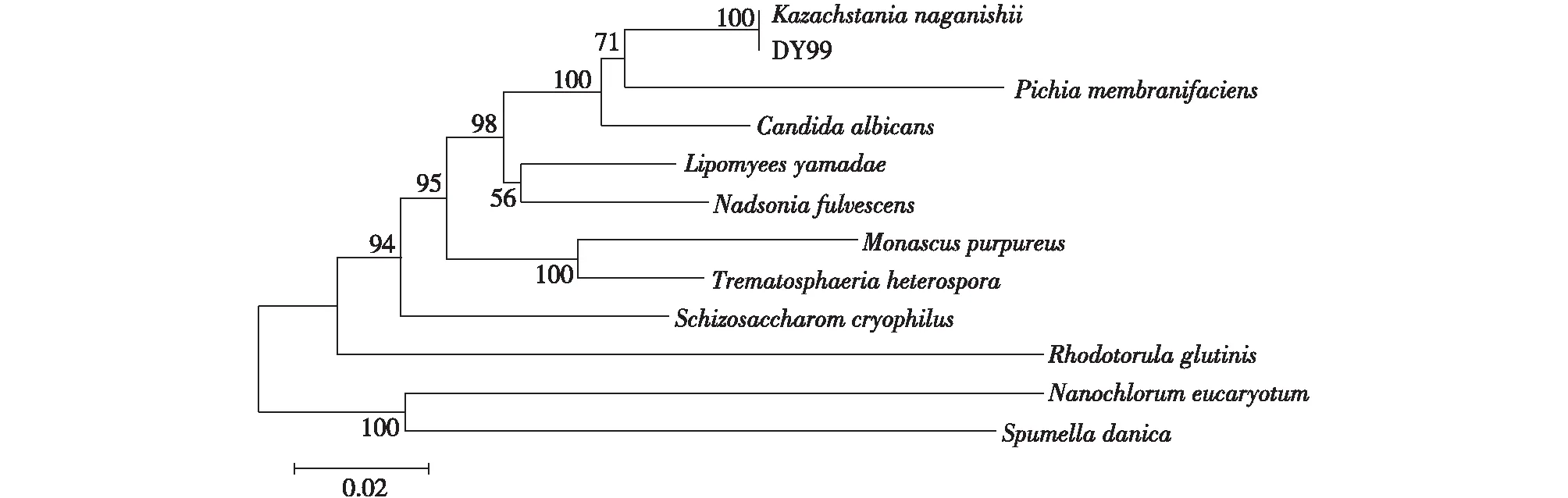
图2 系统进化树
2.2 羰基还原酶基因的克隆与重组工程菌构建
挖掘到1株推定的羰基还原酶,其氨基酸序列(NCBI号:XP_022464010.1)与探针酶相似度为59.18%,与文献[8-9]报道的具有较高R-HPBE产率的羰基还原酶IolS和CgKR2相比,氨基酸序列相似度分别为11.9%和12.2%。
以K.naganishii基因组为模板,采用设计的特异性引物进行RCR扩增,将获得的DNA产物分析后,插入表达载体转入宿主进行表达,并用电泳分析鉴定,结果见图3。由图3可知:通过PCR得到约1 000 bp条带(图3(a)),与目标基因一致(1 049 bp);将扩增获得的基因插入pET-28a表达载体(图3(b)),并将构建的重组表达质粒用热击法转化大肠杆菌BL21(DE3)感受态细胞,获得的重组大肠杆菌E.coli-pET28-kncr经乳糖诱导后,SDS-PAGE凝胶电泳分析(图3(c)),获得约4.0×104蛋白条带,与目标蛋白CR预测分子量一致(3.846×104)。由此可知,重组CR菌构建成功。
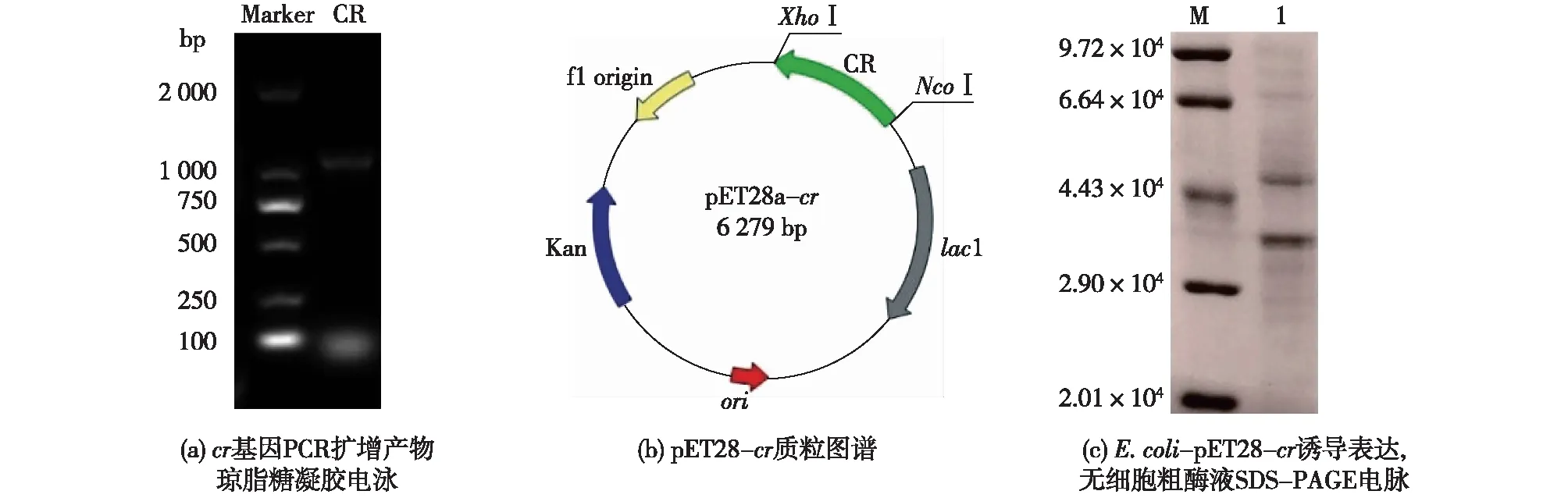
图3 E. coli-pET28-kncr重组菌构建
2.3 羰基还原酶重组菌诱导表达条件优化
为获得最佳诱导条件,对诱导剂(乳糖)浓度和诱导时间进行了优化,结果见图4。由图4可知:增加乳糖有利于细胞的增殖,在较低乳糖(0~6 g/L)条件下,乳糖的增加有利于酶的合成,发酵液总酶活也随之增加,但当乳糖超过6 g/L,酶蛋白可溶性表达量逐渐降低,导致发酵液的总酶活下降。采用6 g/L乳糖为诱导剂时,生物量和总酶活均随时间的延长而增加,但在诱导12 h后,酶蛋白可溶性表达量开始下降,导致总酶活降低。由此可知,在最佳诱导条件(28 ℃、150 r/min、6 g/L乳糖诱导12 h)下,最高总酶活为330.61 U/L。
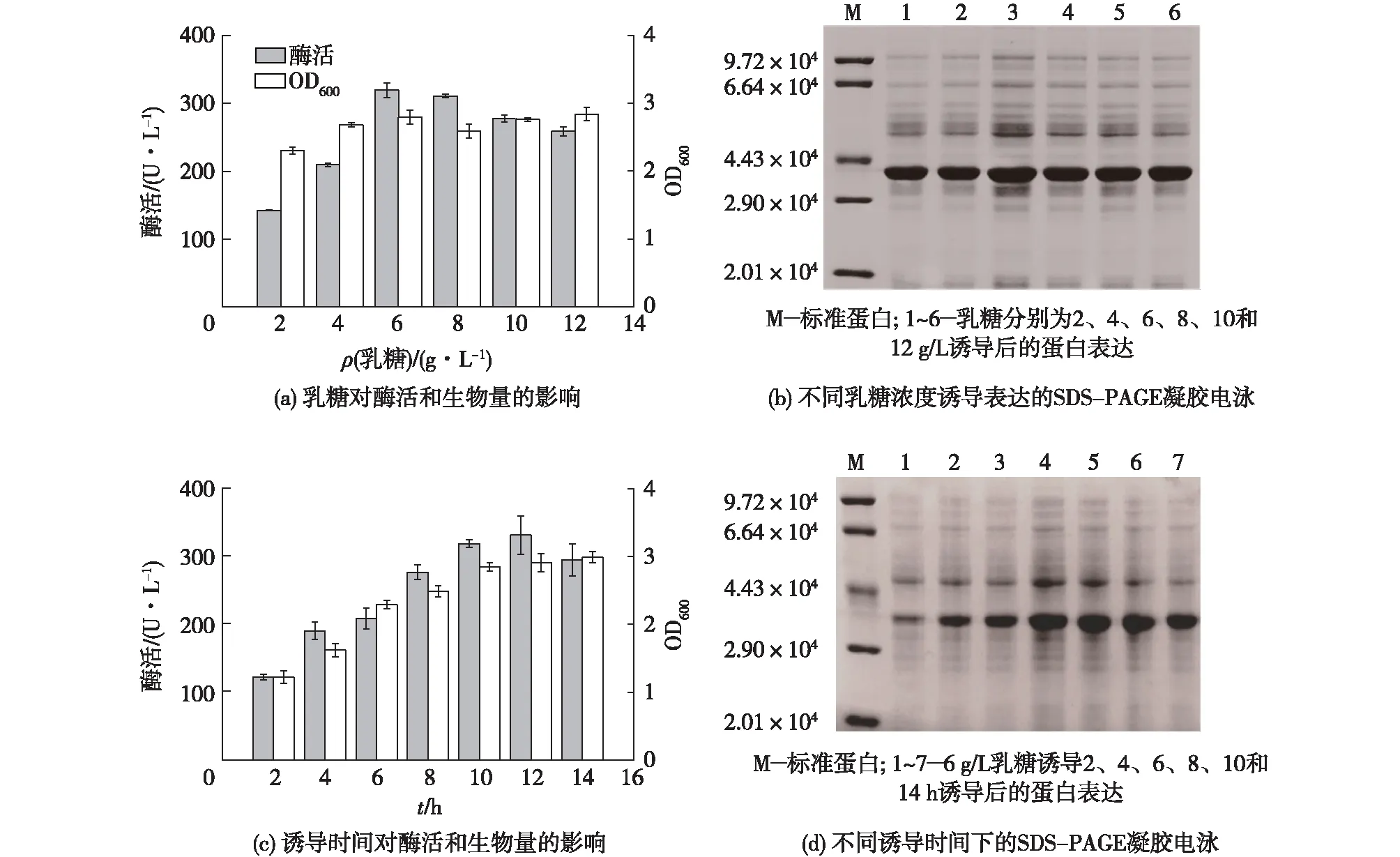
图4 诱导条件优化
2.4 催化反应条件优化
pH的变化会引起酶分子构象、酶和底物的解离状态发生变化,进而影响酶与底物的结合,使酶促反应速率发生变化。采用不同pH的磷酸钾缓冲液作为反应介质,在10 mL反应体系中,添加OPBE 10 g/L,葡萄糖5 g/L,重组菌细胞20 g/L(以干质量计),在30 ℃、200 r/min条件下研究pH对不对称还原反应的影响,结果见图5(a)。由图5(a)可知:随着pH的升高,产物累积浓度也逐渐增大,至pH7.5时最大,达到1.13 g/L;继续增大pH导致产物累积浓度下降。在所研究的范围内,pH的改变未引起酶立体选择性的显著变化,不同pH条件下产物的e.e.均大于99%。由此确定,反应最适pH为7.5。
在一定范围内,温度的升高有助于酶分子与底物分子的有效碰撞,从而使酶促反应速率加快;而另一方面,随着温度的升高,酶分子变性加剧,活性酶分子减少,反应速率降低。酶促反应最适温度是这两方面因素平衡的结果。在10 mL反应体系中,添加OPBE 20 g/L,葡萄糖5 g/L,重组菌细胞10 g/L(以干质量计),以100 mmol/L磷酸钾缓冲液为反应介质,控制pH为7.5,温度25~45 ℃,转速200 r/min进行不对称催化反应,结果见图5(b)。由图5(b)可知:当温度为30 ℃时,产物累积浓度达到峰值,继续升高温度,酶变性失活加剧,产物累积浓度变小。当温度为45 ℃时,反应液由澄清透明逐渐变浑浊,有白色黏稠固体产生,产物累积质量浓度仅为0.49 g/L。由此可见,羰基还原酶重组菌催化OPBE不对称还原获得R-HPBE最适反应温度为30 ℃。由于野生菌细胞中含有不同选择性的还原酶,因此以野生菌细胞为催化剂还原OPBE生成产物的光学纯度不高,采用基因工程技术克隆单个还原酶,构建重组菌并异源表达酶蛋白来催化不对称还原反应,可获得光学纯的产物。
烟酰胺辅酶是氧化还原反应中的关键影响因素。葡萄糖可作为氢供体,促进细胞内辅酶再生。在10 mL反应体系中,添加OPBE 10 g/L,葡萄糖5~30 g/L,重组菌细胞20 g/L(以干质量计),以100 mmol/L磷酸钾缓冲液为反应介质,控制pH为7.5,温度30 ℃,转速200 r/min进行不对称催化反应,结果见图5(c)。由图5(c)可知:当葡萄糖为10 g/L(与OPBE浓度相同)时,产物累积质量浓度达到最大值1.56 g/L,e.e.>99%;再增加葡萄糖用量,反应液黏度升高,传质阻力增大,反而使R-HPBE浓度下降。由此可见,当葡萄糖与底物按1∶1(质量比)添加,不对称还原反应速率最大,能获得最大产物累积量。
在10 mL反应体系中,添加OPBE 10 g/L,葡萄糖10 g/L,重组菌细胞10~50 g/L(以干质量计),以100 mmol/L磷酸钾缓冲液为反应介质,控制pH为7.5、温度30 ℃、转速200 r/min进行不对称催化反应,结果见图5(d)。由图5(d)可知:E.coli-pET28-kncr细胞生物量为20 g/L(以干质量计)时,R-HPBE累积量最大,为最适细胞生物量。增大细胞生物量也会引起反应体系的黏度增加,使反应速率缓慢下降,导致产物积累量减少。
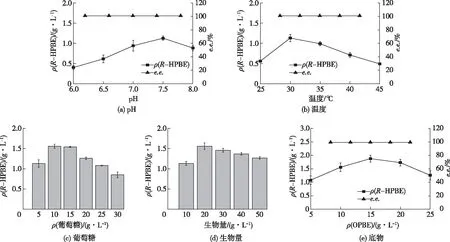
图5 全细胞催化OPBE不对称还原反应条件优化
在已获得的最优条件下,考察了底物浓度对反应的影响,结果如图5(e)所示。由图5(e)可知:在较低底物用量(5~15 g/L)时,底物用量的增加有利于反应速率的提升,产物累积浓度增加;最适底物为15 g/L,在上述最适条件下,经历0.5 h反应后,产物累积质量浓度为1.88 g/L,e.e.>99%,但是进一步提高底物浓度,反应速率降低,表现出明显的产物抑制。
2.5 全细胞催化不对称还原反应时间进程
反应在100 mL三颈圆底烧瓶中进行,反应体系50 mL,含15 g/L OPBE、15 g/L 葡萄糖、20 g/LE.coli-pET28-kncr细胞和50 mL磷酸钾缓冲液(100 mmol/L,pH7.5),恒温磁力搅拌器控制温度为30 ℃、转速500 r/min,流加1 mol/L Na2CO3使pH维持在7.5,反应12 h考察全细胞催化不对称还原反应的时间进程,每1 h取样进行气相色谱检测,结果见图6。由图6可知:随着反应时间的延长,底物OPBE不断消耗,产物R-HPBE不断累积,当产物达到11 g/L时,底物消耗变得缓慢,至反应10 h后基本不变,维持在约3 g/L,由此可知,羰基还原酶的酶活受到高浓度产物的抑制。R-HPBE累积质量浓度在10 h时达到最大(11.2 g/L),产物得率为74.7%,时空产率为26.88 g/(L·d),产物e.e.至始终维持在99%以上。

图6 E.coli-pET28-kncr全细胞催化OPBE不对称还原反应时间进程
表1为不同羰基还原酶重组菌不对称催化合成R-HPBE的结果。由图表1可知:应用KnCR重组菌全细胞催化合成R-HPBE效率与文献[9]报道的最大值700.0 g/(L·d)还有较大的差距。本实验还存在着一些问题,一是存在产物抑制现象,二是缺少烟酰胺辅酶再生系统。Liu等[12]以来源于圆红冬孢酵母(Rhodosporidiumtoruloides)的羰基还原酶重组菌为催化剂,以水-正辛醇(4∶1,体积比)两相体系为反应介质,催化(S)-6-氯-5-羟基-3-羰基己酸叔丁酯不对称还原,反应在9 h内达到平衡,每克湿细胞生成(3R,5S)-6-氯-3,5-二羟基己酸叔丁酯的时空产率为4.08 mmol/(L·h),是单水相介质催化时空产率的1.12倍。Shah等[13]在体积比为1∶1.5的水-甲苯介质中应用来源于醋酸杆菌(Acetobacteraceti)的醇脱氢酶,催化合成(S)-4-氯-3-羟基-丁酸乙酯的时空产率达到650.4 g/(L·d)。筛选合适的有机溶剂,构建水-有机两相反应介质能减轻甚至消除底物和产物抑制,因此能有效增强催化反应效率。羰基还原酶需要还原型烟酰胺辅酶(NAD(P)H)传递氢和电子,所以还原型烟酰胺辅酶的充分供应,是不对称还原反应顺利进行的重要保障。然而,还原型烟酰胺辅酶价格昂贵,批量使用会大幅抬高生产成本。Chen等[14]用葡萄糖脱氢酶还原再生NADPH,并与来自开菲尔乳杆菌(Lactobacilluskefir)的醇脱氢酶耦联不对称催化合成(S)-2-氯-1-(2,4-二氯苯基)乙醇,仅添加0.05 g/L NADP+,60 g/L底物在6 h内完全转化,生成产物e.e.为99.5%。Li等[15]通过加入甲酸-甲酸脱氢酶辅酶原位再生系统,使二乙酰还原酶KpDAR不对称还原合成(S)-3-hydroxy-2-butanone累积量由39.4 g/L提升至52.9 g/L,时空产率2.0 g/(L·h)增加到6.2 g/(L·h)。由此可见,添加辅酶来再生酶,构建双酶耦联辅酶原位再生体系,是解除辅酶限制、降低生产成本的常用方法。
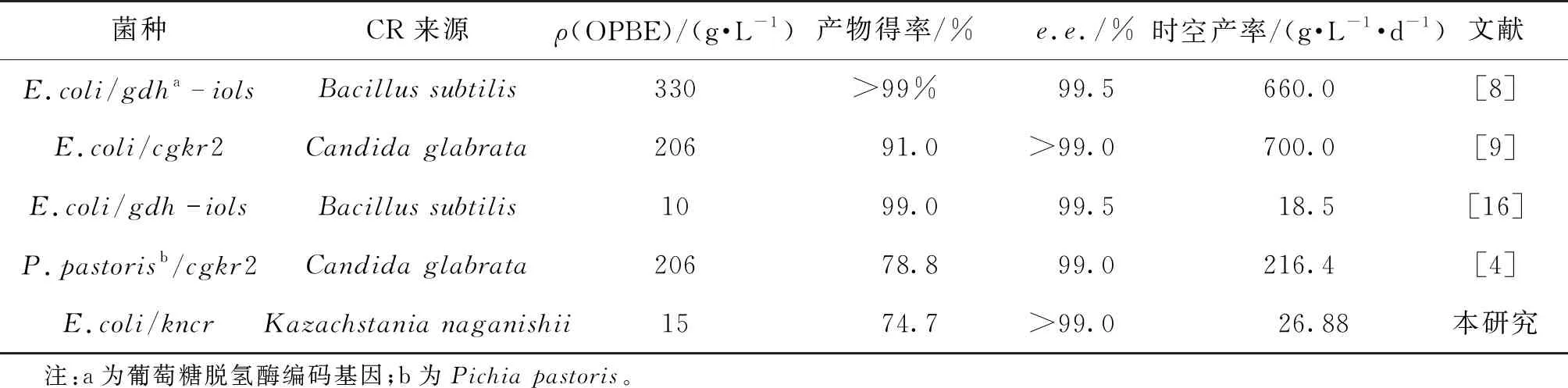
表1 不同羰基还原酶重组菌不对称催化合成R-HPBE的结果
3 结论
筛选获得了一株具有催化OPBE不对称还原生成血管紧张素转换酶抑制剂类降压药关键手性中间体R-HPBE的野生型菌种,鉴定为Kazachstanianaganishii。通过基因挖掘,获得了羰基还原酶基因序列,克隆获得了酶基因并构建了重组大肠杆菌E.coli-pET28-kncr。以重组羰基还原酶为催化剂,以潜手性化合物OPBE为底物,不对称合成R-HPBE。优化获得了重组菌诱导培养条件:当细胞培养物OD600达到0.8时,以6 g/L乳糖为诱导剂,在28 ℃、150 r/min条件下培养12 h,在30 ℃、pH7.5磷酸缓冲液中,底物、葡萄糖、细胞生物量分别为15、15和20 g/L(以干质量计)时,重组菌全细胞催化反应效率最高,为最适反应条件;在上述最优条件下,50 mL恒pH反应获得光学纯R-HPBE,时空产率达到26.88 g/(L·h),具备了工业化应用前景。但在反应过程中底物和产物均对反应产生明显抑制,使应用该羰基还原酶生产R-HPBE受到限制。
