喜旱莲子草入侵群落主要物种生态位和种间联结研究
2021-11-04郭佳琦陈俊辰黄旬黄佳乐赵丽娅李兆华
郭佳琦,陈俊辰,黄旬,黄佳乐,赵丽娅,李兆华
湖北大学资源环境学院,湖北 武汉 430062
生态位是物种适应某一空间环境资源的综合结果,展示的是物种在群落中利用自然资源的能力、占据生态空间的大小以及相对地位(潘高等,2015),定量地反映了种群间及与环境之间的耦合关系。种间联结通常以物种的存在与否为依据(Legendre et al.,1983),用来描述由群落生境差异而造成不同物种分布的相互关系(房飞等,2012;丁文慧等,2016),定性地表述了群落演替的现状或种间竞争的结果(徐满厚等,2016),两者均会影响群落的稳定性。因此,结合生态位特征、种间关联和群落稳定性,有助于共同揭示群落结构、演替趋势以及物种的空间分布以及对环境资源的需求性等(江焕等,2019)。
喜旱莲子草(Alternanthera philoxeroides)是苋科莲子草属多年生草本植物,原产于南美洲,已入侵中国近 90年,具有极强的繁殖能力和表型可塑性,目前已侵害我国大部分地区,入侵生境包括水生(湖泊、河流、沟渠)、陆生(农田、撂荒地、房前屋后、路边)和水陆交错带(湖滨带、沼泽湿地)等。以往的研究多围绕喜旱莲子草的生物学特性(王颖等,2015)、地理分布与风险预测(Hu et al.,2020)、防控手段与应用(宋振等,2018)等方面展开,而关于喜旱莲子草入侵群落伴生物种间的联结关系的研究尚少(郭连金等,2009;吴昊等,2019)。采用生态位特征、种间联结性等数量学方法探讨被入侵群落中各物种的相互作用和种间关系逐渐成为入侵生态学的研究热点,已有研究表明外来植物紫茎泽兰(Eupatorium adenophorum)、反枝苋(Amaranthus retroflexus)、苏门白酒草(Coryza sumatrensis)的入侵使原有植物群落处于不稳定状态,降低了群落多样性并且与本地物种之间存在一定的竞争(郭连金,2011;赵彩莉等,2013;刘海等,2017;江焕等,2019)。因此,探究入侵种的群落特征以及本地物种对入侵的响应有利于理解其入侵机理,可为预测和评估入侵物种的危害程度以及入侵物种的生物防治提供参考依据(徐满厚等,2016)。
湖北省位于长江中游,由于年均气温和土壤水分等自然条件适宜,喜旱莲子草于20世纪50年代入侵湖北后迅速扩散蔓延,尤以湖泊、沟渠、沼泽等湿润生境危害严重,目前已在沙湖等湖泊的湖滨带形成入侵态势,造成植物群落物种多样性降低,严重威胁湖滨带生态系统稳定性。本研究以沙湖喜旱莲子草群落为研究对象,通过探讨喜旱莲子草入侵群落主要物种的生态位特征和种间联结,明确其与伴生物种的种间关系和相互影响,对了解入侵群落结构和演替方向具有重要意义,以期为喜旱莲子草入侵后植物群落的管理提供方向。
1 材料与方法
1.1 研究区概况
研究区位于武汉市中心城区(图 1),介于30°33′—30°34′N、114°18′—114°20′E,是内环线之内的最大湖泊,西临长江2.1 km,东距东湖2.9 km,属于亚热带季风性湿润气候,湖区年均气温15.8—17.5 ℃,雨热同季,雨量充沛且集中在6—8月;冬季和夏季分别长达110 d和135 d;年无霜期一般为211—272 d,年日照总时数为1810—2100 h;土壤类型以红壤、黄棕壤和水稻土为主;植被类型主要为常绿阔叶林和落叶阔叶林组成的混交林,主要乔木有樟(Cinnamomum camphora)、夹竹桃(Nerium oleander)、水杉(Metasequoia glyptostroboides)、乌桕(Triadica sebifera)等;湖滨带灌草层植物主要有双穗雀稗(Paspalum paspaloides)、马唐(Digitaria sanguinalis)、麦冬(Ophiopogon japonicus)、狗牙根(Cynodon dactylon)、结缕草(Zoysia japonica)、沿阶草(Ophiopogon bodinieri)、红花酢浆草(Oxalis corymbosa)、白车轴草(Trifolium repens)、鸢尾(Iris tectorum)、箬竹(Indocalamus tessellatus)等。由于自然环境的适宜性,沙湖环湖近岸范围内(距离湖岸线 (5±2) m)普遍存在喜旱莲子草。
1.2 样方选取及调查
2020年8—9月,在对沙湖进行预调查的基础上,平行于湖滨带进行植物样方调查,为使植被和水位条件一致,以环湖陆向带的灌草层为研究对象,每隔 (200±5) m设置1个1 m×1 m样方,共50个。现场记录每种植物的物种名称、多度、高度、盖度、频度、生境条件以及经纬度、海拔、坡度、坡向等位置信息,不能进行现场鉴定的植物拍照后采集,带回实验室参考《中国植物志》、《中国高等植物图鉴》等进行鉴定。
1.3 研究方法
1.3.1 重要值
物种重要值(IV)是研究物种间关联性的主要数量指标,反映了植物在群落的地位和重要程度。本研究选取重要值>2的物种进行生态位和联结性分析。计算公式如下:

式中:
Rc——相对盖度;
Rh——相对高度;
Rc——相对频度。
1.3.2 生态位测定
本研究把野外群落调查的各样方视为多种资源状态的综合,以各物种在各个样方的重要值(IV)作为指标测定各种群的生态位宽度及生态位重叠,能够避免因物种大小而产生的误差,体现各物种对群落资源的利用效率。计算公式如下(柳剑丽等,2013):
Levins生态位宽度(BL):

Shannon生态位宽度(BS):

式中:
Pij——第i个物种在第 j个资源等级下的重要值占该种在所有资源水平上重要值总和的比例;
r——资源等级数(样方总数)。
Schoener生态位相似性指数:
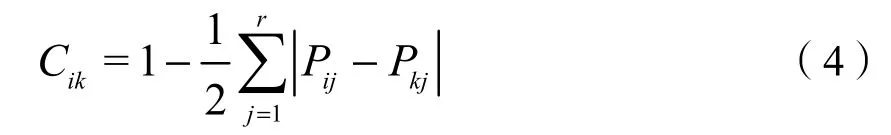
式中:
Cik——物种i和k的生态位相似性指数;
Pij——物种i在资源位j上的重要值。
Pkj——物种k在资源位j上的重要值。
Pianka生态位重叠指数:
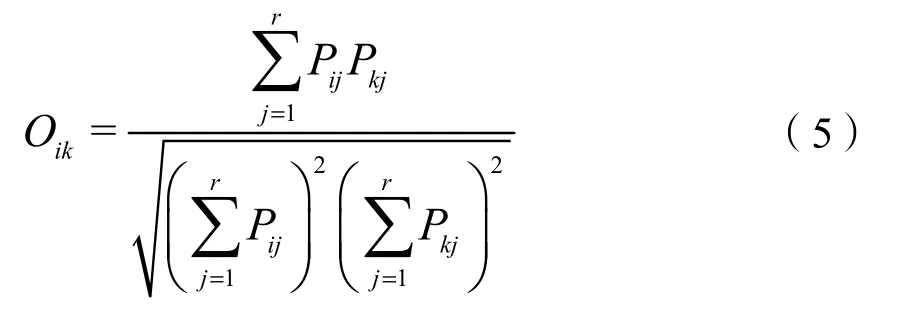
式中:
Oik——物种i和k的生态位重叠指数;
1.3.3 种间总体关联性检验
方差比率(Rv)用于检验多物种间的总体联结性,并结合统计量W检验关联程度。Rv值在独立性假设条件下期望值为1,若Rv=1,则原假设成立,所有物种间表现为无关联;若Rv>1,则所有物种种间表现为净的正联结;若Rv<1,则种间表现为净的负联结。统计量W用于检验Rv值偏离1的显著程度,W 值服从χ2分布(df=N−1),若,则所有物种间关联性不显著,否则,总体呈显著关联。计算公式如下(Schluter,1984):


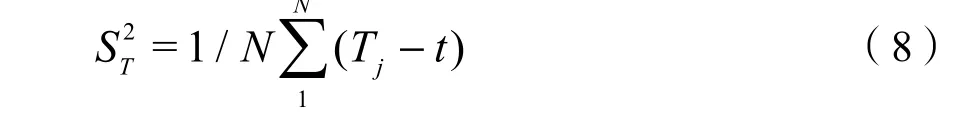
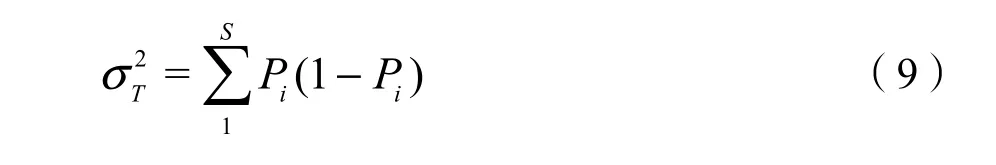

式中:
S——所有物种数;
N——所有样方数;
Tj——样方j内出现的物种数;
T——样方中物种的平均数,t=(T1+T2+…+Tn)/N;
ni——物种i出现的样方。
1.3.4 种对间关联性检验
根据群落中待测定的成对物种在样方中的存在与不存在的观测数据构建2×2联列表。本研究采用Yates的连续校正公式计算χ2统计量判定种间联结性,计算公式如下(郭志华,1997):
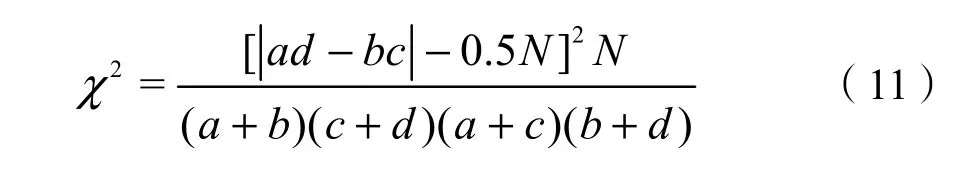

式中:
N——总样方数;
a——物种A、B都存在的样方数;
b——物种A存在而B不存在的样方数;
c——物种B存在而A不存在的样方数;
d——物种A、B都不存在的样方数。
若3.841<χ2<6.635 (0.01
匹配系数Ochai用于进一步验证χ2统计量的结果及说明种对间关联程度。计算公式如下(徐满厚等,2016):
匹配系数 Ochai(IO):
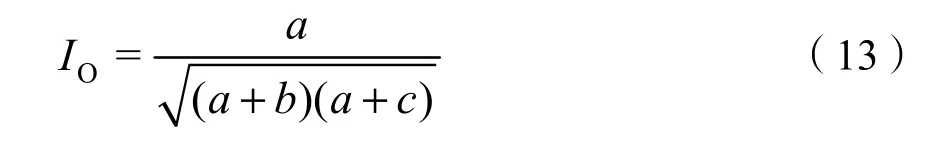
1.3.5 群落稳定性分析
参考改良过的贡献定律法,17个主要物种按照相对频度大小排列后逐步累积,再将植物种数取倒数并按顺序与之对应逐步累积。横坐标(x)为物种倒数累积百分比,纵坐标(y)为累积相对频度,建立模糊散点曲线模型。该比值越接近(20,80),群落就越稳定。计算公式如下(郑元润,2000):
平滑曲线模拟方程:

直线方程:

将直线方程代入平滑曲线模拟方程,解得交点横坐标x:

所有数据的整理分析在SPSS 20.0、Excel 2016和 R 3.5.2中进行处理。种间联结和生态位测定使用 spaa 程序包中 sp.assoc()、sp.pair()、niche.width()、niche.overlap()计算。
2 结果与分析
2.1 重要值和生态位分析
在喜旱莲子草入侵群落共发现草本植物 18科38属 41种,其中禾本科(Poaceae)、菊科(Asteraceae)、旋花科(Convolvulaceae)和苋科植物(Amaranthaceae)较多,分别有10、8、4、3种。重要值>2的主要物种中禾本科植物最多(表 1),有5种(占主要物种的29.41%),其次为菊科和蓼科(Polygonaceae),分别有3种(17.65%)和2种(11.76%)。
重要值体现了物种在群落的地位,生态位宽度能够反映物种对环境的适应状态和资源的利用竞争状况。主要物种的重要值和生态位宽度的变化较大(表 1),其中喜旱莲子草占据最大的重要值(IV=18.06)和生态位宽度(BL=0.80,BS=3.78),麦冬的重要值(IV=2.00)和生态位宽度(BL=0.11,BS=1.91)最小。生态位宽度的排序和重要值的排序和整体趋势相似,但并非完全一致。除喜旱莲子草外,禾本科植物狗牙根、双穗雀稗、马唐的重要值居群落前10%,且生态位宽度较广,表明这些物种在群落中占据优势竞争地位。
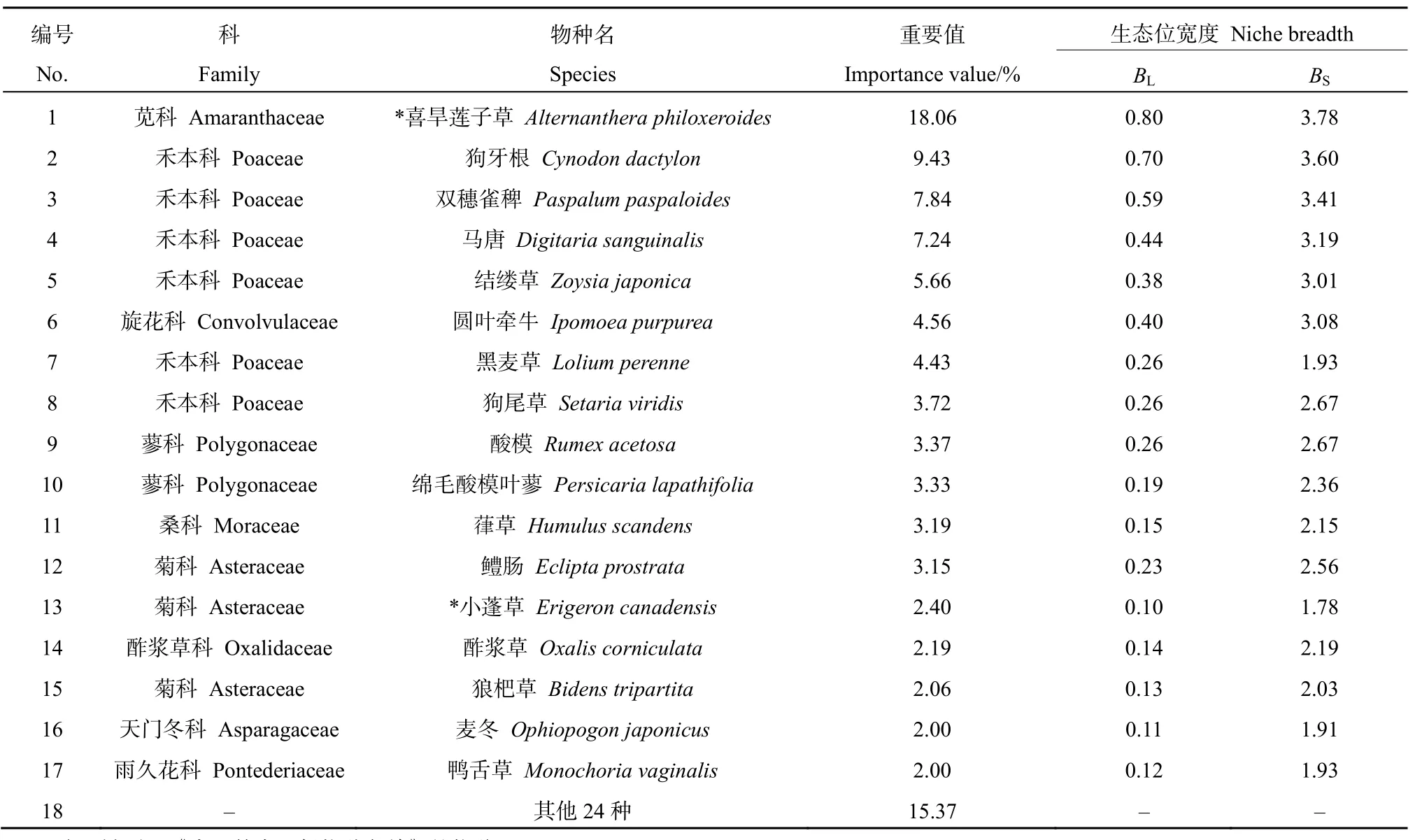
表1 喜旱莲子草群落植物的重要值和生态位宽度Table 1 Importance value and niche breadth of plants in Alternanthera philoxeroides community
生态位相似性指数能反映群落中不同物种利用环境资源的相似程度。由表2可知,物种间生态位相似性指数最大的是双穗雀稗-狗牙根(Cik=0.67),0.5以上的种对有6个(4.41%),介于0—0.5的种对共123个(90.44%),为0的种对有7个(5.15%),表明大部分物种对环境资源的需求有较大差异。喜旱莲子草-狗牙根、喜旱莲子草-双穗雀稗的生态位相似性指数在0.5以上,表明这些种对具有相似的资源需求。

表2 喜旱莲子草群落主要物种生态位相似性指数(对角线左)和生态位重叠指数(对角线右)Table 2 Niche similarity (diagonal to the left) and niche overlap (diagonal to the right) of main plant species in the Alternanthera philoxeroides community
生态位重叠可在一定程度上解释群落种间的竞争和共生关系。物种间生态位重叠最高的是双穗雀稗-狗牙根(Oik=0.74),0.5以上的种对有 14个(10.29%),介于0—0.5种对共115个(84.56%),未发生重叠的种对共7个(5.15%),表明大部分物种间的竞争较小,在资源充裕时尚能共存。喜旱莲子草与狗牙根、双穗雀稗、黑麦草(Lolium perenne)、圆叶牵牛(Ipomoea purpurea)、马唐存在较高的生态位重叠(Oik>0.55),随着环境资源的消耗,喜旱莲子草可能会与这些物种产生激烈的竞争。
2.2 种间关联性
种间关联性检验对于探究群落种对间的相互关系及作用机理具有重要意义。喜旱莲子草入侵群落的种间总体联结性结果见表3,方差比率Rv=0.92,统计量W=45.82落在内,表明群落整体呈现不显著的负关联。χ2检验结果见表4,所有种对中,86个种对呈正关联,47个种对呈负关联,3个种对无关联,正负关联比为1.83。其中,呈极显著正关联的为狼杷草 (Bidens tripartita)-鸭舌草 (Monochoria vaginalis);呈显著正关联的有2对:麦冬-小蓬草(Erigeron canadensis)、圆叶牵牛-小蓬草;呈不显著正关联的有83对;呈极显著负相关的种对为黑麦草-马唐;无种对呈显著负相关;呈不显著负相关的种对有46个;3个种对无关联。以上种对所占比例依次为0.74%、1.47%、61.03%、0.74%、0、33.82%、2.20%。132个种对没有达到显著水平或呈无关联,占总数的97.06%,表明主要物种间联结性较弱,种间关系松散甚至趋于独立,群落极易受到扰动。

表3 种间总体关联性Table 3 Overall interspecific associations among main plant species in the Alternanthera philoxeroides community

表4 主要物种种间关联性的χ 2统计量检验Table 4 χ 2 correlation test of main plant species in the Alternanthera philoxeroides community
匹配系数 IO表示物种间相伴出现的几率和联结性程度,可以克服列联表中d值过高影响而造成的误差问题,作为对χ2检验结果的补充能较准确地反映物种间联结性的强弱(徐满厚等,2016)。由图2可知,喜旱莲子草-双穗雀稗、喜旱莲子草-狗牙根、喜旱莲子草-马唐、双穗雀稗-狗牙根表现出非常紧密的关联性(0.7 图2 主要物种种间匹配系数Ochiai(IO)半矩阵Fig. 2 Semi-matrix diagram of Ochiai index (IO) of main species in the Alternanthera philoxeroides community 由图3可知,平滑曲线模拟方程与直线方程的交点为(37.62,62.38),与稳定点坐标(20,80)有一定距离,结合种间总体呈不显著负相关关联性结果,推断群落目前处于非稳定状态,可能处于演替初期,喜旱莲子草的入侵使得多个物种间存在由资源制约的竞争作用。随着演替的进行,种间会发生激烈竞争,为了降低竞争态势物种间会出现一定的分离,从而使群落达到稳态。 图3 喜旱莲子草群落稳定性分析Fig. 3 Stability graph for Alternanthera philoxeroides community 重要值和生态位宽度都反映了种群在群落中的地位和作用,但两者的生态学意义略有不同,植物的重要值表明了植物在群落中的优势程度,生态位宽度则反映了其对资源的适应能力以及生态适应性等特征(邓贤兰等,2016)。在本研究中,主要物种的生态位宽度与重要值排序基本一致。喜旱莲子草占据最大的重要值和生态位宽度,说明其在群落中占主导地位,具有较强的竞争力以及适应环境、利用资源的能力,这也是喜旱莲子草快速占领新生境、入侵成功的主要原因(王颖等,2015),喜旱莲子草具有水陆两栖的生物学特性,更容易在湖滨带等湿润生境中繁殖扩散。伴生物种中,狗牙根、双穗雀稗、马唐的重要值和生态位宽度均较大,表明这些禾本科植物在群落中占有一定优势地位,这与李安定等(2013)、从春蕾等(2016)、吴昊等(2020)和对喜旱莲子草群落组成的研究结果一致。上述物种在群落中具有较强的竞争力,这与禾本科植物具有较强的生长和繁殖能力、较强的资源竞争能力以及较广的生境适应性有关,故在资源充裕时能与占据主要优势地位的喜旱莲子草共存一定时期;麦冬等植物的重要值和生态位宽度较小,说明这些物种对环境的适应能力、争夺及利用资源的竞争力相对较弱,可能有一定的特化倾向,在资源紧张时处于不利地位。 生态位相似和生态位重叠是是判断物种间利用资源的异同性和反映种间竞争关系的重要指标。生态位宽度越广的物种易形成较高的生态位重叠(郑超超等,2015;刘润红等,2020),本研究证实了这一观点,生态位宽度最大的喜旱莲子草与多数物种产生了较高的生态位重叠和生态位相似,揭示了其作为入侵物种,能与大部分植物甚至生态位较窄的物种具有相似的环境适应性和资源争夺。生态位宽度较高的种对间,例如双穗雀稗-狗牙根、喜旱莲子草-狗牙根、喜旱莲子草-双穗雀稗等种对的生态位重叠和生态位相似性指数均较高,表明这些生态适应性强的物种有相似的生活习性或资源需求,随群落演替的进行将发生激烈竞争。本研究中生态位宽度均较窄的狼杷草-鸭舌草产生了较大的生态位重叠,这可能由于调查区域所在的湖岸湿润生境适于两种植物生存繁殖,故二者同时出现的频率较高,在资源受限时趋向于这种适宜的生境斑块(刘建康等,2014;吴倩楠等,2017),因而产生了较高的生态位重叠。 多数种对的生态位相似性指数和生态位重叠指数在0—0.5之间,表明目前群落中多数物种间竞争作用较小,资源充裕时将存在一定的共存期。有研究表明,外来物种的入侵会明显降低本地物种间的生态位重叠(郭连金,2011;江焕等,2019),并且增强了本地物种与入侵物种的竞争态势(刘海等,2017;吴昊等,2019)。而喜旱莲子草与多数主要物种产生了较高的生态位重叠,从宏观角度分析必然会导致种间竞争的发生。 种间联结理论认为,正联结有2种情况:(1)一个物种对另一物种产生依赖;(2)在异质环境中,几个物种对环境有相似的适应和反映,但往往由于演替后期资源的限制产生激烈的种间竞争。负联结是:(1)物种在竞争资源中相互排斥;(2)环境需求的不相似性(徐满厚等,2016)。一般认为,随着群落演替的推进,群落的组成和结构会不断地完善和稳定,种间关系会趋向于正关联(Greig-Smith,1983;张金屯,2011)。本研究显示喜旱莲子草群落总体呈不显著负关联,种对间关联的显著性仅2.94%,这与苏门白酒草(郭连金,2011)和紫茎泽兰(刘海等,2017)入侵群落的联结特征相似。目前喜旱莲子草入侵群落的结构和功能处于动态调节中,一部分种对存在对光热、水分和养分等资源的争夺互斥,而大部分物种属于正联结的第二种情况,即目前具有彼此适应、共生的能力,但随群落逐渐成熟,由于生态位的分离以及生长习性的不同,物种间会产生竞争排斥作用,并随资源的耗竭将愈发强烈。 结合匹配系数结果,喜旱莲子草与圆叶牵牛、黑麦草、狗尾草(Setaria viridis)等其他13个物种表现出不同程度的负关联,由于不同的生态习性而导致的需求差异,以及喜旱莲子草具有强势的入侵性,与其他植物争夺生存空间和水肥资源而产生种间排斥(赵彩莉等,2013;王颖等,2015)。郭连金等(2009)认为外来植物在开始入侵群落时,由于资源丰富和相似的适应性会与部分本地物种呈现正联结,当种群达到一定量时会因资源争夺而排斥抑制本地物种,由原来的正联结逐渐独立甚至趋于负联结,最终排挤其他植物从而形成单种优势群落。 本研究中,喜旱莲子草仅与双穗雀稗、马唐、狗牙根3种禾本科植物呈现高生态位重叠和紧密正关联,推测这些物种对喜旱莲子草的入侵具有一定抵抗性。罗瑛等(2017)的试验表明,喜旱莲子草水浸提液对禾本科植物的萌发具有“低促进高抑制”的化感作用;从春蕾等(2016)的研究也表明,当喜旱莲子草种群逐步入侵群落时,会通过显著改变土壤和生境而利于同种或他种植物的生存。而张建利等(2018)通过研究喜旱莲子草群落的数量特征,认为禾本科植物对喜旱莲子草的入侵具有竞争替代性;吴昊等(2020)进一步将喜旱莲子草群落划分不同的功能群,指出群落中禾本科和菊科功能群的存在显著提高了群落多样性水平,而增强本土物种多样性被认为是抵抗生物入侵进程的重要途径。在本研究中,喜旱莲子草与3种禾本科植物可能存在某种关联,能在资源充裕时共存一段时间,另一方面也说明此阶段这些物种对喜旱莲子草的侵入存在抵抗能力,在群落演替初期,上述种对有着相似的资源偏好和利用方式,构建了较为紧密的正关联,如禾本科植物一般具有较强的分蘖能力,与喜旱莲子草同样具有快速扩张种群的能力(Wu et al.,2017)和资源竞争能力,但随着群落演替的进行,种间会因生长性的差异而产生激烈的竞争,如禾本科植物的茎秆通常高于喜旱莲子草,占据喜旱莲子草上层的生存空间便于抢夺地上资源,而喜旱莲子草通过匍匐生长,每个茎节都能萌发出不定根,从而抢夺土壤水分和养分,占据下层生存空间。 群落的稳定性结果显示,交点坐标远离(20,80),表明喜旱莲子草入侵群落处于非稳定状态,目前群落的结构组成和功能尚不稳定,多物种联系并不紧密。结合种间关联结果,物种间正负关联比大于 1,大多数物种有着相似的环境需求和适应,但整体上存在资源竞争。 群落稳定性不仅与群落的组成结构和功能相互关联,还受外界扰动的性质和强度的影响(张金屯,2011)。前人对阔叶丰花草(洪思思等,2008)、苏门白酒草(郭连金,2011)、反枝苋(赵彩莉等,2013)、紫茎泽兰(刘海等,2017)等入侵群落的研究表明,外来植物的侵入改变了原有群落的结构和物种组成,增加了群落的不稳定性。本研究区位于城市内陆湖泊,沙湖水系承担一部分调蓄雨洪的功能,加上近岸灌草层物种丰富,植物群落存在一定波动性;除自然因子外,城市湖泊还受到周围城市化扩张以及人类活动等干扰。这些干扰因素在外来物种入侵前改变了原有群落的种间关系和组成,打破了原有植物群落的平衡与稳态(Rousset et al.,2000),增加了外来植物的入侵机会。在喜旱莲子草入侵后,基于较强的表型可塑性和繁殖能力,其种群快速以斑块状扩张成单种优势群落(潘晓云等,2006;王颖等,2015),造成物种均匀度下降,从而减低原有群落多样性和稳定性(吴昊等,2019)。因此,维护群落稳定性有利于减少入侵植物的入侵机会。 物种的生态位特征与其联结性有着紧密的联系。物种间的正关联通常表明种对间有相似的资源偏好及生态位重叠;负关联则体现了物种间的排斥及生态位分离。本研究中,群落总体联结性为不显著负相关,而种对间的正负关联比却大于 1,推断群落处于发育初期,物种间的关联趋向于随机性,未能形成稳定的种间关系,这与喜旱莲子草的入侵性有关,也与湖滨带的自然环境变动有关。一般来说,群落中呈显著正关联的种对数越多,表明种间联系越紧密,群落也就越稳定(Callaway et al.,2006)。而目前喜旱莲子草群落不显著关联的种对占绝大多数,种对间关联的显著性仅2.94%,环境干扰对其稳定性具有决定性作用。因此,建议加强对沙湖湖滨带植物的维护管理来减少喜旱莲子草种群,如采用人工手段清除喜旱莲子草全株,晒干或焚烧全部茎节以防二次入侵。 本研究发现,入侵群落中喜旱莲子草与正联结的双穗雀稗、狗牙根和马唐出现了较高的生态位重叠,这反映了上述种对有着一致的生境资源要求,加之此3种植物的生态位宽度较大,对环境的适应能力强,可考虑在喜旱莲子草已入侵的群落中优先栽培,既可以抵抗喜旱莲子草的入侵,又能在资源匮乏时与其形成激烈竞争,遏制喜旱莲子草的蔓延,同时搭配种植与喜旱莲子草生态位重叠较小的物种如黑麦草、狗尾草等作为伴生种,以防止种间恶性竞争。对于已清除喜旱莲子草的群落,可采用与其生态位重叠较高且呈负关联的物种如结缕草等进行植被恢复。 (1)喜旱莲子草入侵群落的优势物种是喜旱莲子草+双穗雀稗等禾本科植物,喜旱莲子草重要值和生态位宽度均最大(IV=18.06,BL=0.80,BS=3.78),狗牙根、双穗雀稗和马唐的重要值和生态位宽度较高。入侵植物喜旱莲子草与群落中大多数物种产生较高的生态位重叠,其平均值为0.42。 (2)喜旱莲子草与圆叶牵牛、黑麦草、狗尾草等 13个主要物种呈不同程度的负关联,表明喜旱莲子草与多数物种产生了竞争排斥;喜旱莲子草与双穗雀稗、狗牙根、马唐等3种禾本科植物呈现高生态位重叠且密切正关联,在演替初期种对间可能构建某种关联而存在一定共存期,随着群落逐步发育将会产生激烈竞争。同时这些禾本科植物对喜旱莲子草的入侵有一定抵抗作用,可考虑作为替代植物用于入侵初期以及清除后植被的修复管理。 (3)入侵群落总体呈现不显著负关联、非稳定状态,喜旱莲子草的入侵以及湖滨带环境的特殊性使得现有植物群落存在一定波动性,大部分物种间联系不紧密,多数种对会因生长繁殖型的差异而产生分离。而喜旱莲子草与多数物种存在竞争,随着自然条件的变化和演替的深入,竞争将更加激烈。建议加强对湖滨带群落的维护管理,减控喜旱莲子草种群数量以及干扰因素,有利于预防外来物种的入侵。
2.3 群落稳定性

3 讨论
3.1 喜旱莲子草群落主要物种生态位
3.2 喜旱莲子草群落主要物种种间联结
3.3 喜旱莲子草群落稳定性
3.4 生态位特征、种间关联与群落演替
4 小结
