太行山南麓不同海拔梯度天然林优势树种生态位特征
2021-11-04闫东锋张妍妍吕康婷周梦丽王婷赵宁
闫东锋,张妍妍,吕康婷,周梦丽,王婷,赵宁
河南农业大学林学院,河南 郑州 450002
生态位理论是物种在群落或生境中的地位和功能,其在评价种群关系、物种与环境间的关系、群落结构及功能等方面发挥着重要作用。生态位研究有助于了解群落中不同物种之间的竞争与共存关系(Soberon et al.,2005;李一凡等,2020;罗欢等,2021;赵家豪等,2021)。生态位宽度、生态位重叠度和生态位相似性是生态位量化的重要指标(杨新东等,2020),其中生态位宽度较大的物种对资源利用更充分,对环境具有较强适应能力;生态位重叠度较大的物种对具有较相似的生态学特性;物种对的生态位相似性越高表明二者对资源的利用程度和竞争能力接近(苏志尧等,2003;杨新东等,2020;张国强等,2020)。有关不同森林群落类型的种群生态位已有大量研究,如闫淑君等对万木林林隙中的主要树种的生态位进行研究,发现生物学或生态学特性相近的树种具有较大的生态位重叠(闫淑君等,2002);龚辉(2020)研究闽北正红菇宿地森林群落主要种群生态位时发现该森林群落生态位宽度大的树种更容易与其他树种产生生态位重叠;李菁等(2011)对阿尔山兴安落叶松(Larix gmelinii)下植物种群生态位的研究证实了生态位宽度较大的种群与其他种群的生态位重叠及相似性较高,但这也出现在一些生态位宽度较窄的种群,这表明生态位重叠和相似性并非仅仅取决于生态位宽度;胡喜生等(2004)在研究长苞铁杉(Tsuga longibracteata)群落优势种群生态位时,发现长苞铁杉的生态位宽度值较大,群落处于稳定状态,但黄庆丰等(2013)分析安徽铜陵叶次生阔叶混交林主要树种生态位特征认为该群落尚未达到稳定状态。上述研究有助于理解植物群落物种多样性的形成和维持机制,但是鉴于天然林物种间的生态位分化及其与环境之间的复杂关系,对天然林优势树种生态位随环境梯度变化的研究十分必要。
太行山南麓地区天然林分布广泛,在调节区域碳平衡、维持生物多样性等方面发挥着重要作用(赵辉,2018),该区较大的海拔差造就了垂直分布较为明显的物种分布格局,如在低于 1200 m的相对低海拔区,乔木树种以栓皮栎(Quercus variabilis)为主,海拔 1200—1500 m的地区则多见槲栎(Quercus aliena)和五角枫(Acer mono)等,1500 m以上的高海拔区以华山松(Tilia japonica)分布最为广泛。利用生态位理论解释太行山地区森林群落物种的种间关系和资源利用程度虽已有研究,但大多集中于灌草层或某一海拔梯度的乔木层树种(李军玲等,2006;闫美芳等,2006;万猛等,2009;王进等,2016)。本研究通过对比分析不同海拔区优势树种重要值及其变异系数、生态位宽度、生态位重叠指数、生态位相似性指数等指标的变化规律,以期揭示不同海拔高程梯度下群落植物物种作用的生态学过程,为该区天然林保护和物种多样性维持等提供参考依据。
1 研究区概况
研究区位于河南省济源市愚公林场的王屋山林区和黄楝树林区,北依太行山,与山西省阳城县、晋城市接壤。该地区平均海拔1000 m,气候属半湿润、半干旱大陆季风性气候,年平均气温 14 ℃,无霜期平均213 d,年平均降水量600 mm,降水多集中在 7—9月,全年>10 ℃年积温为 2753—3671 ℃。土壤类型主要为棕壤土和褐土,植被类型为暖温带落叶阔叶林,区域内植物以华北地理区系为主,多为栓皮栎林、槲栎林、黄栌(Cotinus coggygria)灌丛、荆条(Vitex negundo var.heterophylla)灌丛。
2 研究方法
2.1 样地设置
2020年8月,在研究区海拔850—1850 m天然林分布区选择林分保存完整、人为干扰较少的地段设置29块规格为20 m×20 m的固定样地,涵盖落叶阔叶林和针阔混交林2种典型天然植被类型,其中低海拔区(850—1150 m)13块,中海拔区(1150—1450 m)6块,高海拔区(1450—1850 m)10块。对样地内所有胸径 (DBH)≥1 cm、高度(H)≥1.3 m 的木本植物进行每木检尺,记录树木的名称、胸径、树高、生长状况,同时记录样地中心点海拔高度、经纬度坐标和坡度等环境因子,样地基本情况见表1。
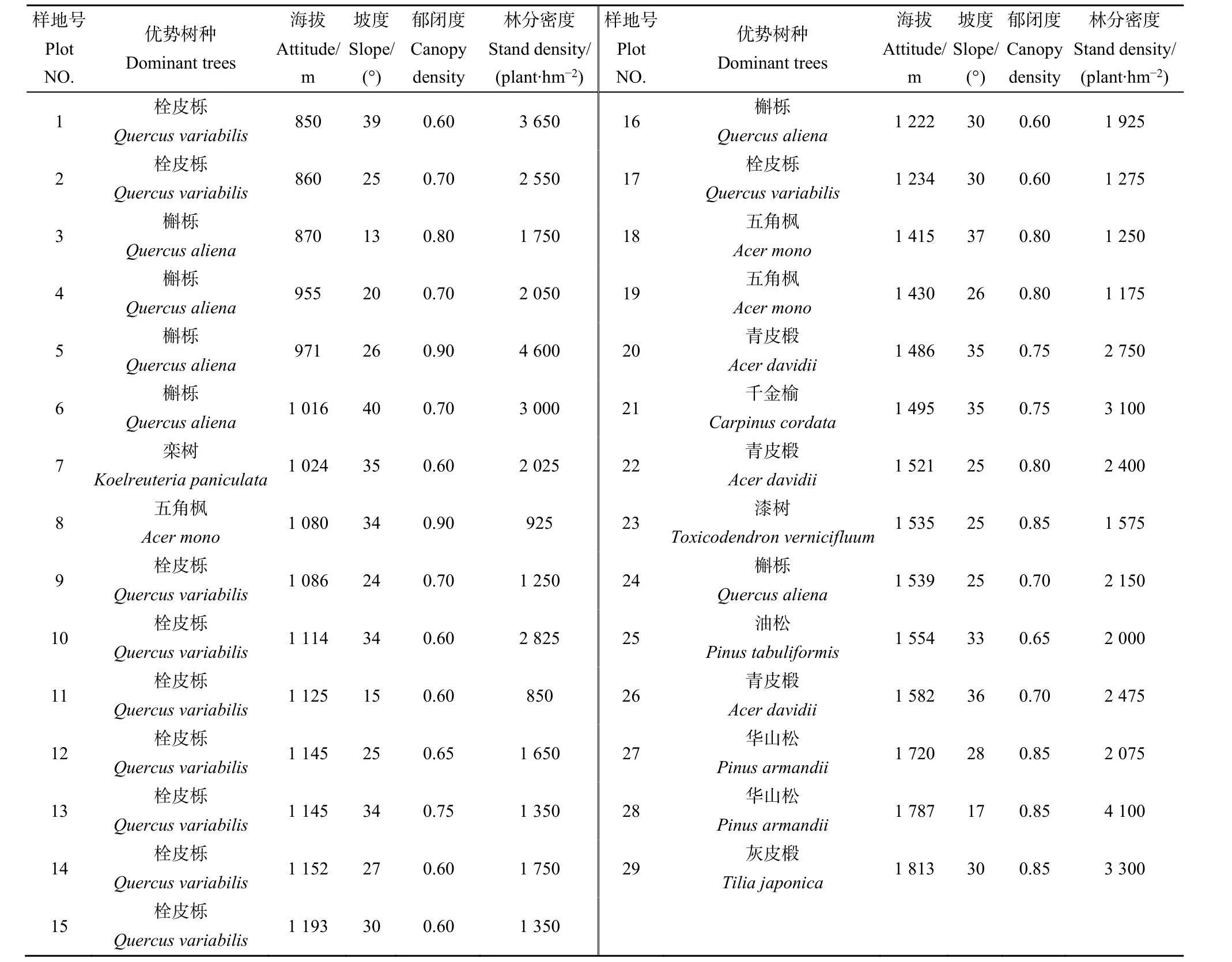
表1 样地基本信息Table 1 Basic information of sample plots
2.2 物种重要值及变异系数计算
物种重要值计算公式:

式中:
IV——物种重要值;
RA——相对多度;
RF——表示相对频度;
RD——相对显著度。



式中:
A——某物种的个体总数;
∑A——乔木层所有物种的个体总数;
F——表示某物种在样地中出现的频度;
∑F——乔木层所有物种的频度总和;
D——某物种所有个体的胸高断面之和;
∑D——乔木层所有物种胸高断面积总和(赵家豪等,2021)。
物种重要值变异系数计算公式:

式中:
CV——物种重要值的变异系数;
SD——各物种重要值的标准差;
M——各物种的平均值。
2.3 生态位指数计算
2.3.1 生态位宽度
生态位宽度分别采用Levins生态位宽度B(L)i和Shannon生态位宽度B(S)i计算公式(郭燕等,2018):




式中:
B(L)i——取值范围为[1/r, +∞];
B(S)i——取值范围为[0, lnr];
Pij——物种i在资源位j的重要值占其在所有资源位的重要值总和的比例;
Nij——物种i在所有资源位的重要值总和;
nij——物种i在资源位j的重要值;
r——样方数。
2.3.2 生态位重叠指数
生态位重叠度指数采用PianKa计算公式(郭燕等,2018):
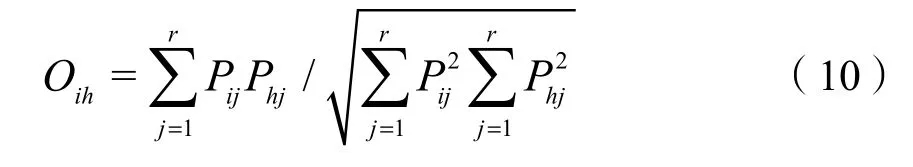
式中:
Oih——物种i和物种h的生态位重叠度指数,取值范围为[0, 1];
Pij——为物种 i在资源位 j的重要值占其在所有资源位的重要值总和的比例;
Phj——物种h在资源位j的重要值占其在所有资源位的重要值总和的比例。
2.3.3 生态位相似性指数
生态位相似性指数 Cih计算公式(苏志尧等,2003):

式中:
Cih——物种i和物种h的生态位相似性指数,取值范围为[0, 1]。
2.4 数据处理
物种重要值的计算在Excel 2016中完成,生态位特征值计算和PCoA主坐标分析及其群落相似性ANOSIM检验分别利用R 4.0.5中的spaa包和vegan包进行。在对原始数据进行正态性检验(Kolmogorov-Smirnov检验)的基础上,对符合t检验条件的数据,利用t检验进行显著性检验。
3 结果与分析
3.1 不同海拔梯度天然林树种结构PCoA主坐标分析
基于29块样地所有68个乔木树种的个体数量计算样地间的Bray-Curtis距离矩阵,在此基础上开展PCoA主坐标分析并绘制样地排序图(图1)。由图1可知,第1排序轴和第2排序轴分别解释了总方差的49.37%和7.56%,不同海拔梯度的样地沿第1和第2排序轴有序排列,但各海拔梯度内的样地均聚集在一起,ANOSIM群落相似性检验结果也表明,ANOSIM统计量r值为0.199(P=0.006),说明不同海拔区乔木树种群落结构存在显著差异。从图1还可看出,相对于高海拔区和中海拔区样地,低海拔区样地之间的离散程度较大,说明低海拔区域各样地之间乔木树种结构的差异性较大。

图1 基于乔木树种群落结构的样地主坐标分析Fig. 1 Principal Coordinate Analysis of plots based on community structure of tree species
3.2 不同海拔梯度天然林优势树种重要值及其变异系数
研究区29块样地中共记录到木本植物68种,隶属于29科52属,本文选取重要值大小前14的树种作为优势树种进行重要值及其变异系数分析,结果见表 2。低海拔区物种重要值较大的树种有栓皮栎(0.389)、槲栎(0.281)、五角枫(0.056)、栾树(Koelreuteria paniculata)(0.048)和君迁子(Diospyros lotus)(0.047);中海拔区物种重要值较大的树种有栓皮栎(0.332)、槲栎(0.211)、五角枫(0.103)、千金榆(Carpinus cordata)(0.086)和青皮椴(Acer davidii)(0.081);高海拔区重要值较大的树种有槲栎(0.227)、青皮椴(0.174)、油松(Pinus tabuliformis)(0.127)、华山松(0.093)和千金榆(0.088)。在所有样地中,重要值较高的树种有槲栎、栓皮栎、青皮椴、千金榆、五角枫,其值分别为0.227、0.213、0.084、0.063、0.060。相同树种在不同海拔的重要值排序位置差别较大,其中栾树的重要值变异系数(5.231)最大,其次是白桦(Betula platyphylla)(3.721)、油松(3.043)和华山松(2.814),这表明海拔因子影响着物种的空间分布及其优势度。
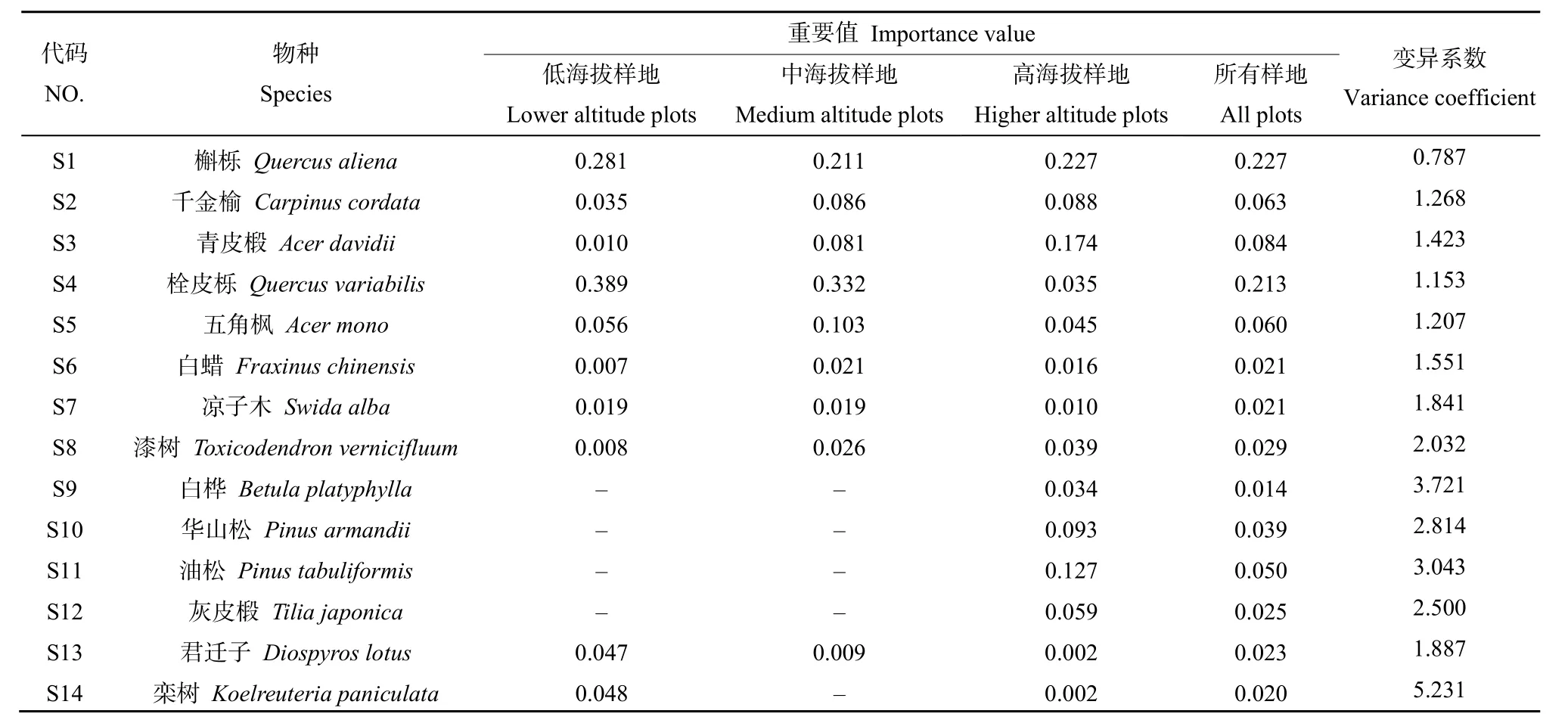
表2 不同海拔优势树种重要值及其变异系数Table 2 Important values and coefficients of variation of dominant tree species at different altitudinal gradients
3.3 不同海拔梯度天然林优势树种生态位宽度
由不同海拔梯度优势树种生态位宽度(表 3)可知,在所有29块样地中,Levins和Shannon生态位宽度最大的树种均为槲栎,其值分别为18.316和3.098;栓皮栎和五角枫的生态位宽度仅次于槲栎,其Levins生态位宽度值分别为12.700和12.054,Shannon生态位宽度值分别为2.752和2.634,说明上述3个树种在不同海拔区分布广泛,对该地区的环境适应能力较强。

表3 不同海拔梯度优势树种生态位宽度Table 3 Niche breadth of dominant species at different altitudinal gradients
不同海拔梯度范围生态位宽度较大的优势树种并不相同,如栓皮栎的Levins生态位宽度值和槲栎的Shannon生态位宽度值在低海拔区最大,分别为8.013和2.153,表明这两种乔木在低海拔区分布较广,资源利用能力较强,在天然林群落中处于优势地位;而除槲栎、栓皮栎和五角枫外,白蜡(Fraxinus chinensis)、漆树(Toxicodendron vernicifluum)和君迁子在中海拔区的生态位宽度亦较大;在高海拔区,槲栎、栓皮栎、五角枫和千金榆的生态位宽度较大,Levins宽度值分别达到18.316、12.700、12.054 和 11.365。
3.4 不同海拔梯度天然林优势树种生态位重叠
对所有海拔及不同海拔区内优势树种生态位重叠度分别进行计算后发现(表4),在全部29块样地中,生态位重叠度最大的物种对是千金榆-青皮椴,其次是五角枫-凉子木,其PianKa生态位重叠度Oih分别为0.73和0.71,生态位重叠值较小的物种对多为栾树与其他物种的重叠。所有优势树种生态位重叠指数均值在不同海拔梯度的大小关系为:中海拔 (0.49±0.35)>高海拔 (0.31±0.27)>低海拔(0.30±0.30),这表明中海拔区优势树种对资源环境需求较为相似,生态位分化不明显,种间竞争较为激烈;而低海拔区和高海拔区优势注重对资源环境利用能力的相似程度均较低,生态位分化明显。
不同海拔梯度优势树种生态位重叠明显的物种对有一定的差别,如低海拔区生态位重叠指数最大的物种对为青皮椴与漆树,其次为五角枫-漆树和五角枫-青皮椴,这说明这些种对对生境要求最为相似。中海拔区生态位重叠度最大的物种对是白蜡-凉子木(Oih=0.95);高海拔区五角枫与青皮椴的生态位重叠度最大(0.90),与其在低海拔区生态位重叠度0.89较为接近,表明这2个树种在低海拔区和高海拔区内生境需求相似,竞争最为激烈。
3.5 不同海拔梯度天然林优势树种生态位相似性
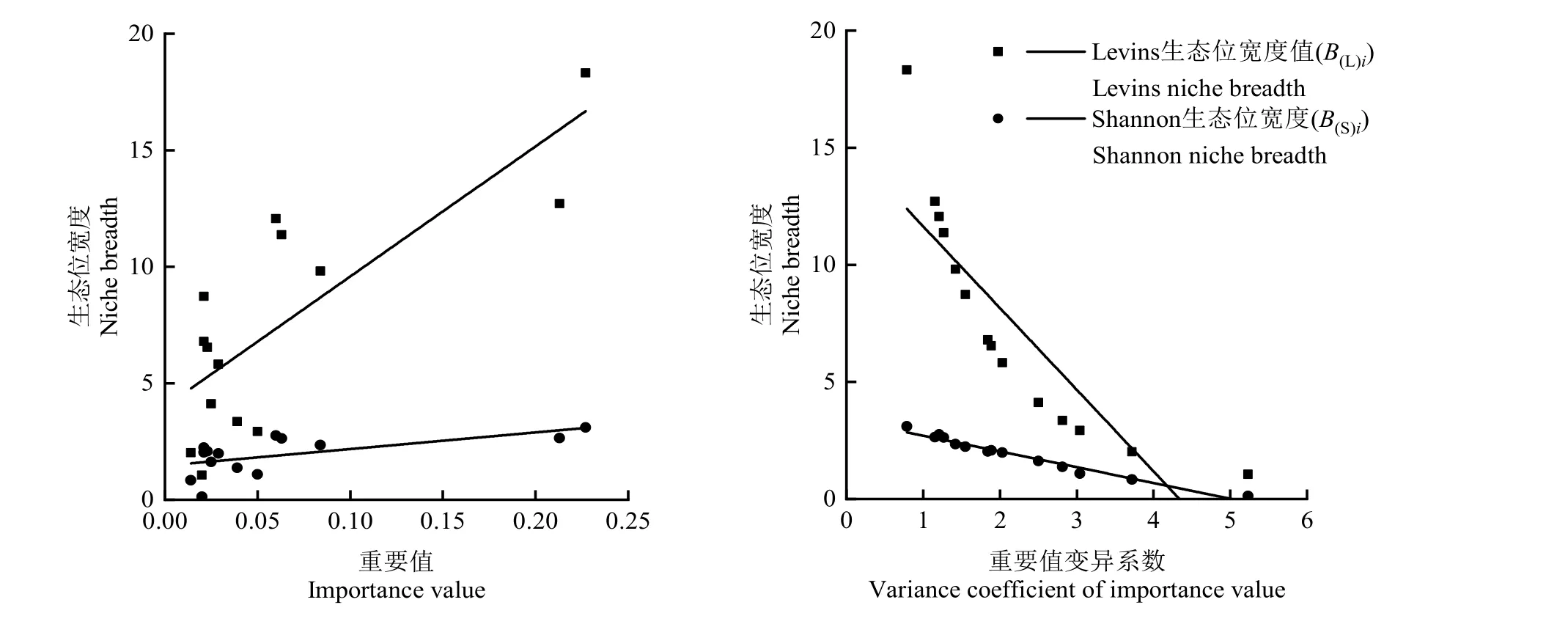
由不同海拔优势树种生态位相似性指数 Cih(表4)可知,3个海拔所有样地优势树种生态位相似性指数Cih均值为0.18,Cih≥0.5的仅有5对(占6.41%),0.3≤Cih<0.5的有 19对(占 24.36%),0 表4 不同海拔梯度优势树种生态位相似性指数及其生态位重叠指数Table 4 Niche similarity index and niche overlap index of dominant tree species at different altitudinal gradients 对所有样地的优势树种重要值分别与其Levins和Shannon生态位宽度值进行相关分析后发现,Levins生态位宽度值(B(L)i)、Shannon生态位宽度值(B(S)i)与重要值之间均呈显著正相关关系,其Pearson 相关系数分别为 0.791(t= −5.777,P=0.010,n=14)和 0.599(t= −8.813,P=0.024,n=14)。回归分析结果表明(图2),Levins生态位宽度值(B(L)i)、Shannon生态位宽度值(B(S)i)与重要值之间均可以采用线性回归方程进行拟合,拟合方程分别为y=55.869x+3.994、y=7.131x+1.462,其决定系数分别为0.626和0.359。重要值变异系数与Levins生态位宽度值(B(L)i)、Shannon生态位宽度值(B(S)i)呈显著负相关关系,其Pearson相关系数分别为−0.858(t=−3.366,P<0.001,n=14)和−0.983(t=0.483,P<0.001,n=14),它们之间的关系亦可用线性回归方程进行拟合,拟合方程分别是 y= −3.486x+15.126、y=−0.673x+3.379,其决定系数分别是0.736和0.967。 图2 生态位宽度值与重要值、重要值变异系数的关系Fig. 2 Relationship between niche breadth and importance value and variance coefficient of importance value 物种重要值能够反映物种在群落中的优势度,生态位宽度能够反映物种对环境资源的利用状况、对环境的适应能力(Arellano et al.,2014;张建利等,2013;万娟等,2014;董东等,2019)。生态位变化的因素包括植物自身生物学特性,外部环境资源变化导致的物种对环境资源需求的差异等(董东等,2019;李一凡等,2020)。本研究中,槲栎和栓皮栎在研究区不同海拔梯度所有样地中生态位宽度最大,是研究区森林群落的主要建群树种,且对光照和生存空间要求较高,相比于中海拔地区的环境条件,2个优势树种在低海拔区和高海拔区适应能力最强。本研究还发现,物种重要值的大小排序与其生态位宽度值排序并不完全一致,在群落中占优势的树种其宽度值并不一定也较大,如青皮椴的重要值在所有优势树种中居第三,但其生态位宽度值却排第五。不同树种对海拔的适应性差异较大,如灰皮椴(Tilia japonica)、白桦、华山松仅在高海拔区出现且有一定的生态位,这应与其不耐热的生态学特性有关。 生态位重叠是物种对同级资源利用的相似性程度及其在空间上的配置关系,物种之间的生态位重叠值越大,其对环境资源的需求条件越相似,越容易出现竞争关系(Muller et al.,2003;曾洪等,2017;王慧敏等,2019;吴霖东等,2020)。本研究发现,物种重要值排前3的优势树种(槲栎、栓皮栎、青皮椴),其种对在中高海拔区的重叠值均表现出较大值,说明这些树种之间在中海拔地区竞争最为激烈。以往研究认为,生态位宽度大的物种其重叠机会大,重叠值较大(Walker,1995;郭平平等,2019;李一凡等,2020),本研究发现,高海拔区宽度值较大的青皮椴-五角枫,其生态位重叠值(Oih=0.90)也较高,与上述发现基本一致。但也有研究认为,种间重叠引起种间竞争不仅与资源匮乏有关,还与物种生态学特性密切相关,生态学特性相似的物种可能出现较高的生态位重叠,有限的资源条件也会引起较高的生态位重叠(李玫等,2005)。 生态位相似性是体现群落中各物种对共同资源的利用关系,物种对之间相似性比例值越小,说明其利用资源的相似程度越低,种群对环境资源分享越充分,群落稳定性越高(许诺等,2017;刘润红等,2018;董东等,2019;杨新东等,2020)。各物种对不同环境因子的适应性存在差异,海拔因子对树种的分布以及其在群落内与其他树种的组合有一定影响(白晓航等,2017;李怀珠等,2020)。大量研究发现,生态位宽度与生态位相似性有一定关联关系,其关系有正相关关系也有无明显关联关系,这主要与物种对的生态习性及立地因子有关(王正文等,2001;刘润红等,2018)。本研究发现,生态位宽度较大的物种对,其生态位相似性也较高,生态位重叠值较大的物种对,其生态位相似性值也较大。如低海拔区宽度值最大的树种槲栎和栓皮栎,其生态位相似性值为0.45;高海拔区重叠值较大的种对青皮椴-五角枫,其生态位相似性值为0.78,均远高于相同海拔区所有优势树种种对生态位相似性值的均值。 本文研究仅考虑了海拔这一立地因子对优势树种重要值及其生态位的影响,未来可以将坡向、坡度、土壤及其他立地因子加入生态位特征分析,这将会有助于对太行山南麓区天然林森林群落稳定性的研究,以及制定出物种多样性维持的最佳策略。 本研究发现,研究区不同海拔区间乔木树种群落结构存在着显著的差异,其中低海拔区乔木树种结构的离散性较大,说明低海拔区乔木树种结构变化较大,对资源的利用较为充分。研究区物种重要值较高的树种有槲栎、栓皮栎、青皮椴、千金榆、五角枫,其中栓皮栎的物种重要值最大,槲栎的生态位宽度最大,其次为栓皮栎和五角枫,说明上述3个树种在不同海拔区分布广泛,对该地区环境的环境适应能力较强,而生态位相似性指数最大的种对是青皮椴-漆树,树种之间竞争较为激烈。相对于而低海拔区和高海拔区,中海拔区优势树种对资源环境需求较为相似,生态位分化不明显,种间竞争较为激烈,因此在森林经营过程中,应注重对中海拔区域树种结构的调整,尤其是减小生态位相似较大的树种之间的竞争,进而促进健康稳定的林分形成。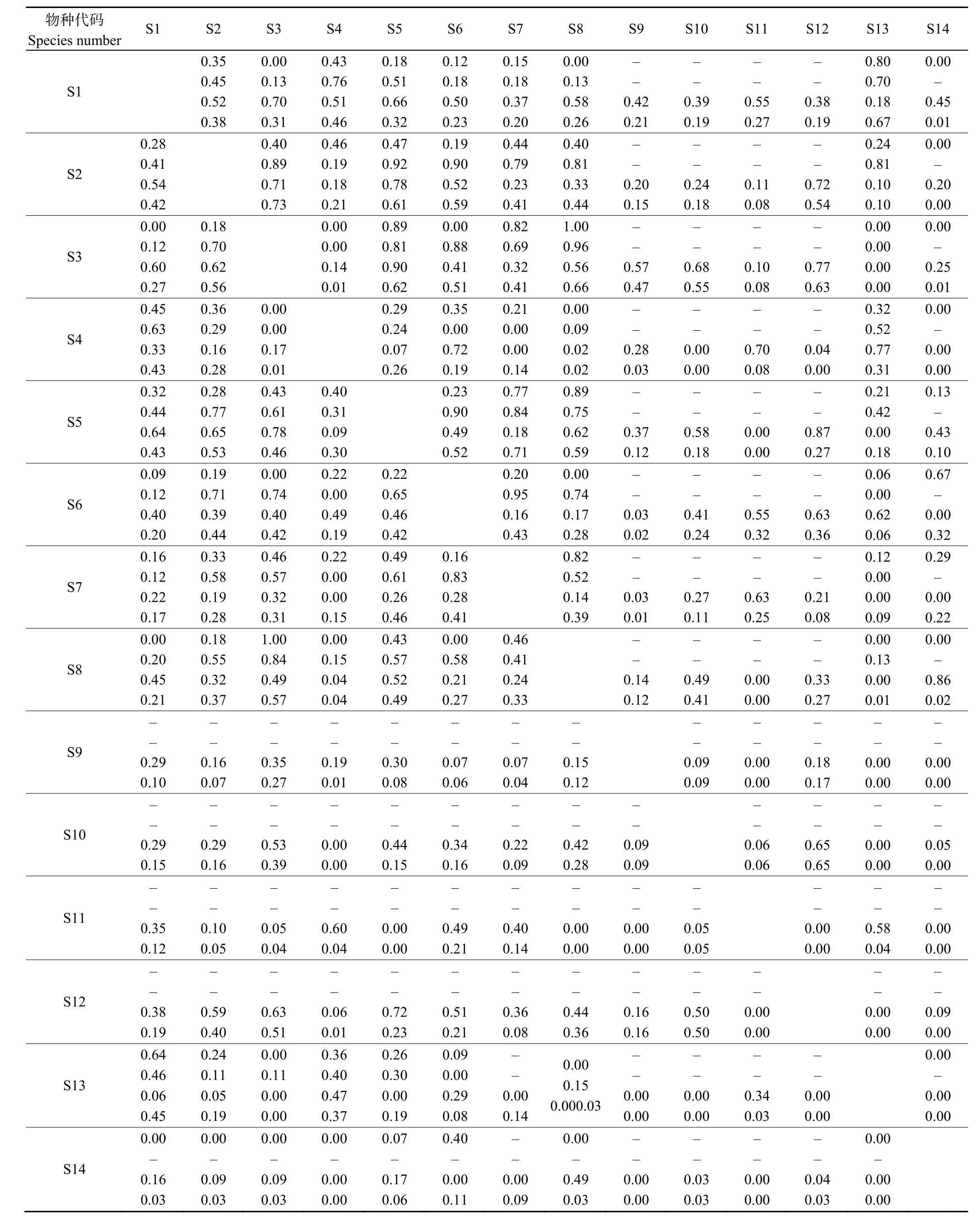
3.6 重要值及其变异系数与生态位宽度的关系
4 讨论
4.1 重要值及生态位宽度
4.2 生态位重叠及其相似性
5 结论
