不同燕麦品种抗氧化系统对秆锈病菌侵染的响应
2021-11-02韩宇婷陈晓晶徐忠山刘景辉薛国兴
韩宇婷,陈晓晶,徐忠山,刘景辉,2,薛国兴
燕麦是重要的粮饲兼用作物之一,我国种植面积约70 万hm2[1],其中裸燕麦种植面积约55 万hm2,居世界首位[2-3]。燕麦具有抗寒、抗旱、耐盐碱、耐瘠薄等特性[4]。燕麦籽粒中富含β-葡聚糖、亚油酸,相比小麦和玉米拥有更大的营养价值和药用价值,并在防治糖尿病和心脑血管系统疾病中发挥重要作用。
燕麦秆锈病(Puccinia graminis f.sp.avenae)是燕麦生产中面临的主要病害之一,近年来,主要在北美[5]、加拿大西部[6]、中国吉林白城和河北张家口[7]等地区发病,造成不同程度的减产。燕麦秆锈病病斑主要生长在叶片上,发病初期夏孢子为圆形暗红色小点,后期燕麦近枯黄时,在夏孢子堆的基础上产生黑色的冬孢子[8]。植株感病会造成千粒重降低、面粉色黑不筋、茎秆易折、饲草质量下降等问题[9]。近年来,我国加大对农业、草牧业、生态等方面的建设力度,燕麦作为粮饲兼用作物种植面积不断扩大,但是,全球气候变暖使燕麦秆锈病发病率呈上升趋势[10]。病原菌入侵后会引起植物体内的一系列防御反应,活性氧反应是其中的一种,对植物的抗病性具有重要的作用[11]。活性氧包括超氧阴离子自由基(·O2-)、羟基自由基(·OH)、单线氧(1O2)和过氧化氢(H2O2)。活性氧在植物体代谢过程中的产生和清除处于动态平衡状态[12]。活性氧的作用有两个方面,一是作为信号分子引起植物的抗病反应,二是活性氧含量过多会破坏细胞膜的结构与功能[13]。超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、苯丙氨酸解氨酶(PAL)都具有清除活性氧的作用[14]。过氧化氢酶(CAT)和过氧化物酶(POD)使植物体内过量的H2O2分解为H2O 和O2;超氧化物歧化酶(SOD)能清除多余的·O2-,维持植物体内活性氧平衡,提高植物抗性[15];苯丙氨酸解氨酶(PAL)是苯丙氨酸途径中重要的酶,是参与植物抗病过程中酚类物质和黄酮类物质合成的关键酶[16]。但是不同的病原菌与寄主,活性氧的产生机制不同,清除活性氧的酶活性高低也大不相同[17]。
有研究表明,条锈病菌侵染小麦感病品种后在侵染早期快速产生H2O2和·O2-,但在抗病品种早期侵染点未检测到·O2-和H2O2的积累[18]。但苜蓿茎点霉侵染苜蓿后抗性品种能快速、大量产生H2O2[19]。樊秦[20]研究表明,在病原菌侵染初期H2O2少量增加,随着侵染程度不断加深,H2O2含量过度积累,对植物造成伤害,病原菌易入侵。枯萎病菌侵染番茄后,抗性品种能快速大量产生活性氧并提高POD 活性[21]。棉花接种黄萎病菌后,SOD、POD 活性均呈上升趋势,且抗病品种增幅大于感病品种[22]。鉴于不同的病原菌与宿主体系变化规律不同,本试验通过研究不同抗性品种燕麦接种秆锈病菌后各器官活性氧变化及防御酶活性变化,探讨燕麦响应秆锈病侵染的生理机制,为燕麦抗秆锈病研究提供理论依据。
1 材料和方法
1.1 试验材料
供试燕麦品种为高感秆锈病品种坝莜1 号(张家口市农业科学院提供)、蒙燕1 号(张家口市农业科学院提供)和抗病品种坝莜18 号(张家口市农业科学院提供)、白燕5 号(吉林省白城市农业科学院提供)[23]。秆锈菌孢子样本于2019年9月在张家口坝上燕麦生产田采集获得,将取回的燕麦完全阴干后用塑封袋包好,放入冰箱4 ℃保存。参照黄振涛等[24]方法分离扩繁。
1.2 试验设计
试验于2020年6—9月在内蒙古农业大学燕麦产业研究中心温室进行。挑选大小一致的燕麦种子,3%的次氯酸钠消毒后摆放至发芽盒内,萌发后移至装有1/2 Hoagland 营养液的聚乙烯盆中培养[25]。当幼苗生长一叶一心时,将新鲜燕麦秆锈病夏孢子6×1010个悬浮液用毛笔均匀接种在燕麦叶片上,对照涂抹蒸馏水(CK)。接菌后的幼苗和CK 置18~22 ℃黑暗保湿箱内保湿24 h,之后取出在18~25 ℃、光照16 h、湿度60%~80%的温室中隔离培养,分别在接菌后1、3、5、7、9 d 取样。每个处理每个时间点选择5株不同燕麦植株。
1.3 测定指标及方法
1.3.1 活性氧含量 过氧化氢(H2O2)含量测定:参照SERGIEV 等[26]的方法,接种秆锈病菌后在1、3、5、7、9 d 分别取样,称取新鲜叶片0.5 g,5 mL 0.1%的三氯乙酸冰浴研磨,在12 000 r/min,4 ℃下离心20 min,取1 mL 上清液分别加入1 mL 0.1 mol/L pH值7.0 的磷酸缓冲液和2 mL 1 mol/L 的KI 溶液,摇匀,放置20 min 后,用分光光度计在390 nm 处测OD值。超氧阴离子自由基(·O2-)采用羟胺氧化法测定[27]。
1.3.2 防御酶系统 超氧化物歧化酶(SOD)活性:采用氮蓝四唑(NBT)光还原法测定[28]。过氧化物酶(POD)活性:采用愈创木酚法测定[29]。过氧化氢酶(CAT)活性:采用过氧化氢法测定[30]。苯丙氨酸解氨酶(PAL)活性:在接种病原菌后1、3、5、7、9 d 分别取样,称取新鲜叶片0.2 g,用pH 值8.8 的硼酸缓冲液加PVP 冰浴中研磨成匀浆,然后移入10 mL 容量瓶中定容,4 ℃条件下,4 000 r/min 离心30 min,取上清液即为粗提酶液。取1 mL 0.02 mol/L L-苯丙氨酸,2 mL pH 值8.8 的硼酸缓冲液,0.2 mL 酶液,以缓冲液代替酶液为对照,摇匀后立即在752B 型分光光度计290 nm 处测定起始OD 值,将第1 次测定后的各支试管在30 ℃恒温水浴锅中保温,30 min 后分别测定OD 值,两次值的差值即为反应液的酶活性[31]。
1.4 数据处理与方法
采用SPSS 19.0 软件进行数据分析,Microsoft Excel 2016 软件进行整理绘图。
2 结果与分析
2.1 秆锈病菌侵染对燕麦叶片H2O2 含量的影响
接种秆锈病菌后,感、抗病燕麦品种叶片H2O2含量呈现不同的变化趋势(图1)。感病品种坝莜1 号、蒙燕1 号在接菌后叶片H2O2含量整体呈现先上升后下降的趋势。接菌后5 d 达到最大值,较CK分别显著升高92.8%、155.5%(P<0.05)。抗病品种坝莜18 号、白燕5 号在接菌后1、5 d 叶片H2O2含量出现峰值。抗病品种坝莜18 号在接菌后1、5 d 叶片H2O2含量较CK 显著升高32.3%、99.1%(P<0.05),抗病品种白燕5 号在接菌后1、5 d 叶片H2O2含量较CK 显著升高34.6%、113.6%(P<0.05)。接菌后抗病品种比感病品种积累更多的H2O2。
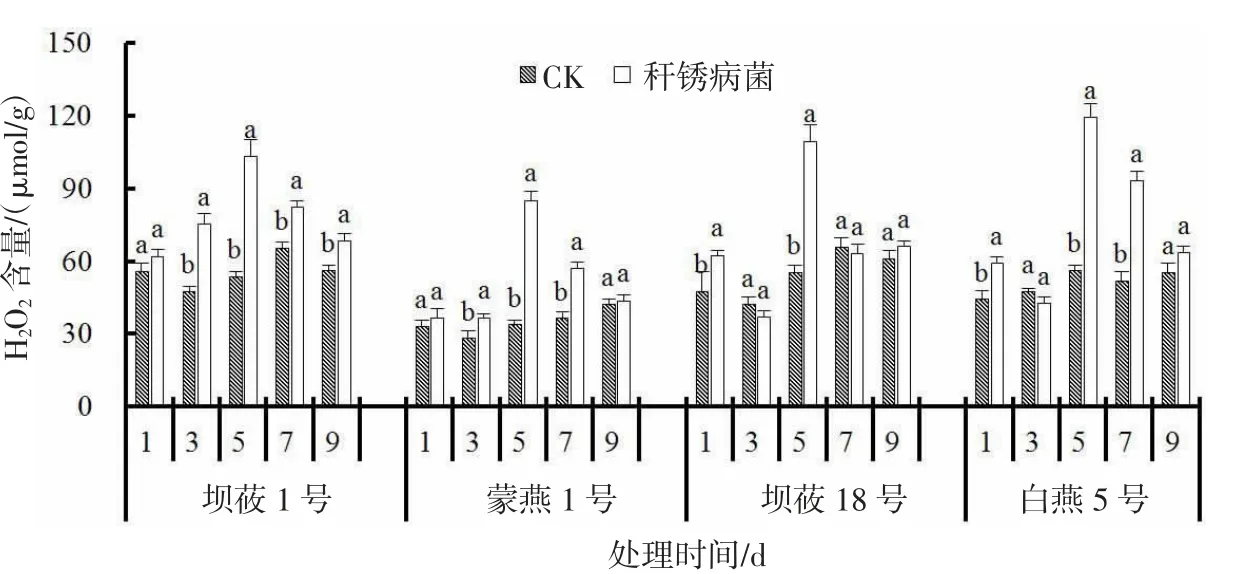
图1 秆锈病菌侵染对燕麦叶片H2O2 含量的影响
2.2 秆锈病菌侵染对燕麦叶片·O2-含量的影响
接种秆锈病菌后,感、抗病燕麦品种叶片·O2-含量均呈现上升的趋势(图2)。与CK 相比,感病品种坝莜1 号、蒙燕1 号叶片中·O2-含量较CK 平均增幅75.3%、42.7%,抗病品种坝莜18 号、白燕5 号叶片中·O2-含量较CK 平均增幅43.9%、40.2%。感病品种坝莜1 号、蒙燕1 号上升幅度大于抗病品种坝莜18 号、白燕5 号。
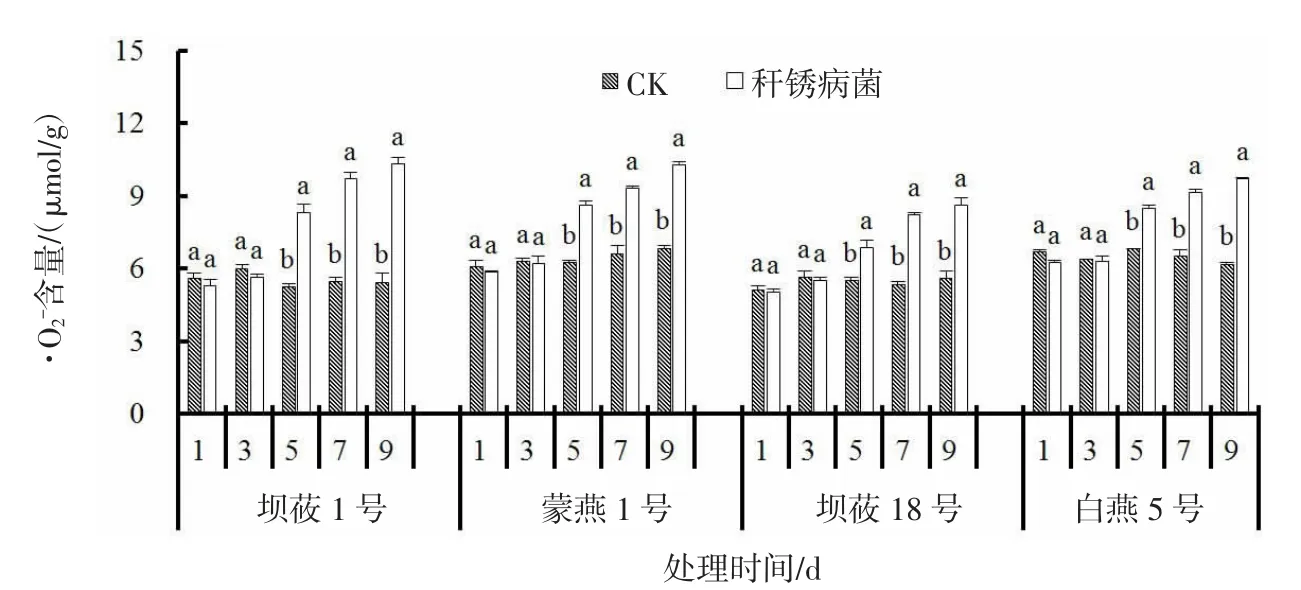
图2 秆锈病菌侵染对燕麦叶片·O2-含量的影响
2.3 秆锈病菌侵染对燕麦叶片SOD 活性的影响
秆锈病菌侵染燕麦后,感、抗病品种SOD 活性在3、9 d 出现两个峰值(图3)。感病品种坝莜1 号、蒙燕1 号在接菌后3 d 分别较CK 显著升高22.2%、63.2%(P<0.05),抗病品种坝莜18 号、白燕5 号在接菌后3 d 分别较CK 显著升高50.0%、48.0%(P<0.05)。接菌后9 d 感病品种坝莜1 号、蒙燕1 号SOD 活性相较CK 增加53.1%、47.5%,抗病品种坝莜18 号、白燕5 号SOD 活性相较CK 增加73.8%、79.2%。接菌后抗病燕麦品种SOD 活性增幅大于感病燕麦品种。
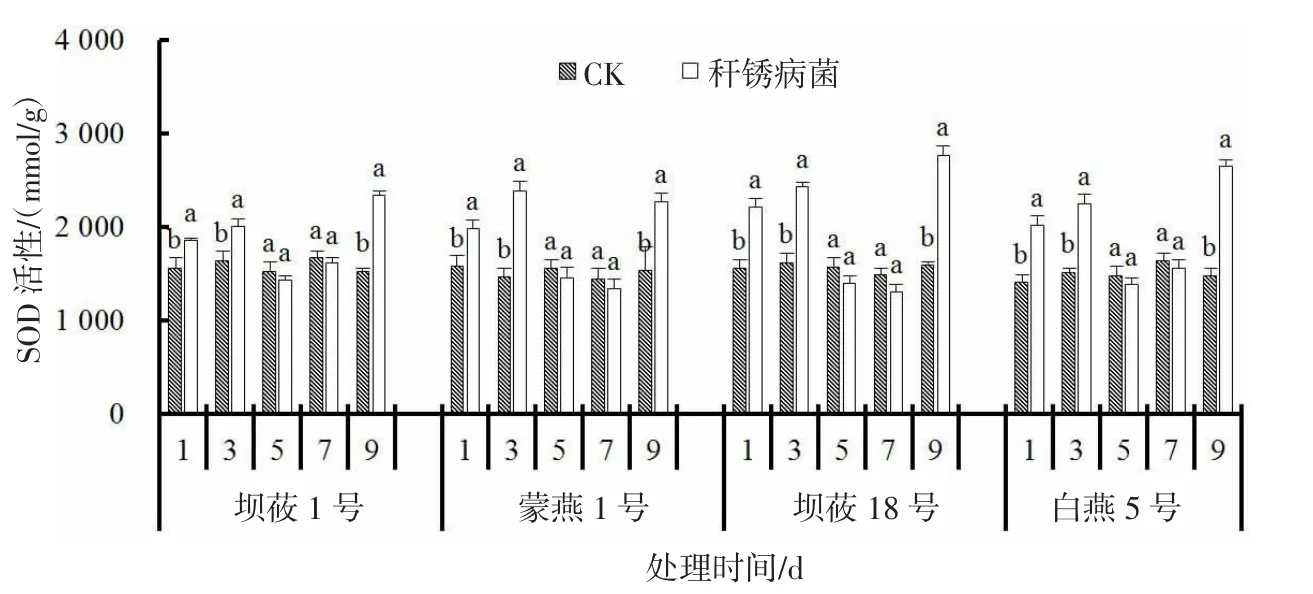
图3 秆锈病菌侵染对燕麦叶片SOD 活性的影响
2.4 秆锈病菌侵染对燕麦叶片POD 活性的影响
秆锈病菌侵染燕麦后,叶片POD 活性均呈现逐渐上升的趋势(图4)。接菌后1、3、5、7、9 d,感病品种坝莜1 号叶片POD 活性较CK 增加10.8%、14.5%、34.3%、52.4%、39.4%,蒙燕1 号叶片POD 活性较CK 增加17.4%、28.4%、35.8%、32.1%、34.3%;接菌后1、3、5、7、9 d,抗病品种坝莜18 号叶片POD活性较CK 增加14.7%、31.0%、50.7%、51.4%、92.1%,白燕5 号叶片POD 活性较CK 增加4.1%、0.9%、19.1%、29.7%、69.0%。
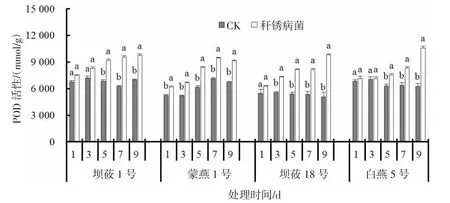
图4 秆锈病菌侵染对燕麦叶片POD 活性的影响
2.5 秆锈病菌侵染对燕麦叶片CAT 活性的影响
秆锈病菌侵染燕麦后,4 个燕麦品种叶片CAT活性呈现不同的变化趋势(图5)。接菌后1、3 d,感病品种坝莜1 号、蒙燕1 号叶片CAT 活性较CK 分别显著上升19.0%~39.2%、14.7%~46.5%(P<0.05);接菌后5 d,蒙燕1 号叶片CAT 活性与CK 相比显著下降19.9%(P<0.05);接菌后7、9 d,坝莜1 号、蒙燕1 号叶片CAT 活性与CK 相比分别上升24.8%~30.7%、20.4%~30.0%,差异达到显著水平(P<0.05)。接菌后抗病品种坝莜18 号、白燕5 号叶片CAT 活性先降低再升高。接菌后1、3 d,坝莜18 号、白燕5号较CK 叶片中的CAT 活性分别降低11.0%~12.2%、28.0%~31.9%;接菌后5、7、9 d,坝莜18 号、白燕5 号叶片CAT 活性较CK 上升27.6%~55.8%,37.2%~48.3%。
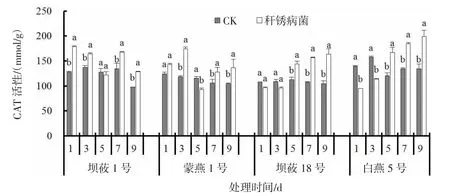
图5 秆锈病菌侵染对燕麦叶片CAT 活性的影响
2.6 秆锈病菌侵染对燕麦叶片PAL 活性的影响
秆锈病菌侵染燕麦后,叶片PAL 活性呈现增加的趋势(图6)。接菌后1、3、5、7、9 d,感病品种坝莜1 号叶片PAL 活性较CK 增加6.7%、17.8%、10.7%、29.3%、28.1%,而蒙燕1 号叶片PAL 活性较CK 增加11.1%、21.3%、30.5%、27.0%、27.0%。接菌后1、3、5、7、9 d 抗病品种坝莜18 号叶片PAL 活性较CK 增加31.8%、21.8%、25.2%、34.6%、28.8%,白燕5 号叶片PAL 活性较CK 增加9.0%、23.4%、24.2%、33.3%、37.9%。

图6 秆锈病菌侵染对燕麦叶片PAL 活性的影响
3 讨论
植物体内的活性氧一直处于一个动态平衡的状态,当外界病原菌侵染时,会打破细胞内的活性氧产生和清除的动态平衡[32]。抗氧化酶CAT 和SOD 一起参与H2O2的代谢,在植物抗病中发挥重要作用[33]。番茄通过增大活性氧在寄主体内的积累,导致SOD活性增加,增强番茄的抗性[34]。张丽等[35]研究指出,病原菌侵染苜蓿后抗病品种通过提高过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、苯丙氨酸解氨酶(PAL)的活性,增强植物的抗病性。
本研究表明,接种秆锈病菌后对燕麦叶片活性氧的平衡造成了很大影响。秆锈病菌侵染前期坝莜1 号、蒙燕1 号叶片中SOD 活性、CAT 活性上升,使植物体内产生的·O2-被歧化H2O2,H2O2被CAT 降解为H2O 和O2,因此,在秆锈病侵染前期坝莜1 号、蒙燕1 号叶片活性氧积累较少;之后叶片SOD 活性、CAT 活性下降,使植物体内产生的H2O2、·O2-被大量积累。坝莜18 号、白燕5 号在接种秆锈病后前期叶片SOD 活性上升、CAT 活性下降,导致H2O2在叶片中被积累,猜测H2O2可能作为植物的抗病信号,引发植物的防御反应[36-37]。这与小麦洛夫林13 接种不同致病力的条锈病菌后植物产生抗病反应[38]和苜蓿抵抗苜蓿茎点霉的早期机制[12]研究结果一致。在接菌7 d 坝莜1 号、蒙燕1 号、坝莜18 号、白燕5 号叶片中SOD、CAT 活性与CK 相比均上升。此时酶活性上升可能是由于底物ROS 浓度增加诱导了酶活性的增加[39]。
4 结论
秆锈病菌侵染后导致燕麦叶片活性氧含量均有所上升,抗病燕麦品种叶片中H2O2含量分别在接菌1、5 d 出现两个高峰,感病燕麦品种叶片在接菌5 d达到最大值;接菌后感、抗病燕麦品种叶片·O2-含量均呈上升趋势,抗病燕麦品种活性氧含量增幅整体大于感病燕麦品种。接菌后感、抗病燕麦品种叶片中SOD、POD、PAL、CAT 活性上升,且抗病燕麦品种增幅大于感病燕麦品种,说明抗病燕麦品种通过快速产生活性氧,促使燕麦较早产生防御反应,抵抗外界病原菌入侵。
