虾青素预处理通过调控Sirt1/miR-134信号通路改善脑缺血再灌注大鼠认知功能*
2021-10-20古春青张运克杨广华武继涛
古春青, 张运克, 杨广华, 武继涛
(河南中医药大学第一附属医院脑病科,河南郑州 450000)
缺血性中风是指由于脑血管闭塞引起的脑缺血,具有很高的发病率、致残率、致死率和复发率,近年来,其发病呈年轻化趋势,已成为医学界和社会关注的重大问题[1]。缺血再灌注(ischemia/reperfusion,I/R)损伤在缺血性中风的发生中起重要作用,据报道,血管再通后恢复到缺血区域的血流会在一定程度上加重缺血性脑损伤,从而加重病情,这种病理过程被称为脑缺血再灌注(cerebral ischemia/reperfusion,CI/R)[2]。CI/R 可引起神经元损伤,导致认知功能障碍,甚至导致“血管性痴呆”,严重影响患者的生活质量,增加中风的死亡率和复发风险[3-4。因此,有必要进一步探索其潜在的信号传导机制,并探索新颖的有希望的干预措施以减轻中风引起的认知功能障碍。
虾青素(ataxanthin,ATX)是一种类胡萝卜素,具有强大的抗氧化剂和抗炎活性。最近,ATX 因其预防或治疗包括阿尔茨海默病和帕金森病在内的神经退行性疾病的作用而受到关注[5]。ATX 对大脑具有很强的保护作用,其独特的化学结构使其可以轻松穿越血脑屏障,可减少I/R 损伤引起的神经元缺陷[6];还可通过抑制炎症反应和氧化应激,减轻阿霉素诱导的认知障碍[7]及血管性痴呆小鼠的认知障碍和海马神经元损伤[8]。然而,目前尚无研究报道ATX能否减轻CI/R所致认知功能障碍。脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)参与神经元的存活、生长和新神经元的分化[9],与神经元损伤密切相关,是治疗中风的重要靶点[10];其表达受环磷酸腺苷反应元件结合蛋白(cAMP response element binding protein,CREB)的调控,而且在多种神经退行性疾病中CREB/BDNF 信号通路的活化与认知功能的改善紧密相关[11]。沉默信息调节因子1(silent information regulator 1,Sirt1)是一种NAD+依赖的脱乙酰酶,在大脑中具有多方面的作用,与神经可塑性密切相关,而Sirt1 的缺失可通过miR-134 介导的转录后机制减弱CREB 活性,导致BDNF 的表达下调,从而削弱突触可塑性,损害大脑的认知功能[12]。以上研究提示Sirt1/miR-134 信号通路可能介导CREB/BDNF 信号通路的活化,从而参与大脑认知功能的调节,故本研究拟通过建立CI/R大鼠模型,探究ATX 对CI/R 大鼠认知功能的影响及可能的作用机制。
材 料 和 方 法
1 动物
从北京维通利华动物中心购入7~8 周龄SPF 级雄性SD 大鼠60 只,体重(300±20)g,许可证为SCXK(京)2019-0009。将大鼠饲养在可控的环境[温度(22±2)℃,湿度(55±5)%,12 h明/暗交替]中,所有大鼠可自由饮水和摄食。研究遵循《实验动物的护理和使用指南》,按实验动物使用的3R 原则给予人道的关怀,并由机构动物护理和使用委员会批准。
2 主要试剂与仪器
ATX(HPLC≥97%)购自Sigma-Aldrich;线栓购自北京西浓科技有限公司;EX527(Sirt1 抑制剂)购自ApexBi;HE 染色试剂、RIPA 裂解液和BCA 试剂盒购自碧云天生物科技公司;TUNEL 原位细胞凋亡检测试剂盒购自武汉博士德公司;Trizol RNA 分离试剂、PrimeScript™RT 试剂盒和SYBR®Premix Ex Taq™II试剂盒购自TaKaRa;兔抗大鼠Sirt1、BDNF、CREB、p-CREB 和β-actin 抗体及山羊抗兔IgG H&L 均购自Abcam;RT-qPCR 引物由GenePharma 公司合成,序列见表1。Morris水迷宫实验装置(WMT-100S)购自成都泰盟软件有限公司;荧光定量PCR 系统(Prism®7300)购自ABI;倒置荧光显微镜(IX73)购自Olympus;蛋白转膜装置购自Bio-Rad。

表1 RT-qPCR引物序列Table 1. Sequences of the primers for RT-qPCR
3 主要方法
3.1 实验分组 SD 大鼠60 只随机分为假手术组(sham 组)、模型组(CI/R 组)、ATX 低剂量(50 mg/kg[13])组(ATX-L 组)、ATX 高 剂 量(100 mg/kg[13])组(ATX-H 组)和ATX(100 mg/kg)+Sirt1 抑制剂EX527(10 mg/kg[14])组(ATX+EX527 组),每组12 只。分组完成后各ATX 组给予相应浓度的ATX 溶液(ATX 溶解于5%羧甲基纤维素钠中)灌胃;ATX+EX527 组在灌胃100 mg/kg ATX 的同时,腹腔注射10 mg/kg 的EX527(DMSO 溶解稀释);sham 组和CI/R 组给予等体积的5%羧甲基纤维素钠灌胃和DMSO 腹腔注射,每天1次,连续给药3 d。
3.2 模型制备 末次给药1 h 后,采用线栓法构建CI/R 大鼠模型[6]:用戊巴比妥钠(50 mg/kg)腹腔注射麻醉大鼠,沿中线切开颈部皮肤;然后,仔细解剖右颈总动脉(common carotid artery,CCA)、颈外动脉(external carotid artery,ECA)和颈内动脉(internal carotid artery,ICA);在CCA 和ECA 的近端结扎后,将一根线栓从CCA 插入右ICA,直到感觉到轻度的阻力,表明大脑中动脉(middle cerebral artery,MCA)闭塞(MCA occlusion,MCAO),导致MCA 提供的区域血流暂时停止;MCAO 2 h 后,取出线栓以使血液通过右ICA 回流(实现再灌注);随后,用4-0 丝线缝合皮肤,在整个过程中,保持大鼠的体温在(37±0.5)℃。sham 组大鼠进行相同的手术程序,不同之处在于不将线栓插入ICA阻塞血管。
3.3 神经功能缺损评分 再灌注24 h 后,由对实验组不知情的研究人员进行神经学评估。参照Longa评分标准:0 分,无缺损;1 分,提尾时不能充分伸展对侧前肢;2 分,行走时向对侧轻度旋转;3 分,行走时向对侧倾斜;4 分,不能自发行走或伴意识障碍。分数越高表示神经功能缺损越严重。
3.4 Morris水迷宫实验 包括定位航行实验和空间探索实验两个部分。将一水池(直径120 cm,高50 cm)平均分为Ⅰ、Ⅱ、Ⅲ和Ⅳ象限,在第Ⅳ象限中央距水平1 cm 处放置一圆形隐藏平台。水中倒入墨水,保持水温21~23℃。通过计算机视频跟踪系统记录实验中的逃逸潜伏期和观察并记录在目标象限(象限Ⅳ)中停留的时间(时间百分比)及平台穿越的次数。(1)定位航行实验:每次从不同象限将大鼠放入池中,逃避潜伏期记录为大鼠从起点到目标平台的游泳时间;若大鼠120 s 内未找到平台,则潜伏期记为120 s,并将其引入平台停留10 s;每天训练4 次,每次间隔30 min,连续4 d,记录每天逃避潜伏期的平均值。(2)空间探索实验:定位航行实验第5 天,撤掉平台,将大鼠从第Ⅰ象限放入水中,记录大鼠60 s内在目标象限中停留时间的百分比和平台穿越次数。逃避潜伏期越长,在目标象限中停留的时间越短,穿越平台的次数越少,表示实验大鼠的认知功能越差。
3.5 HE 染色观察海马神经元损伤 行为学检测完成后,大鼠麻醉、经心灌注后,通过断头迅速处死大鼠并取出脑组织。随机选取6 只大鼠的脑组织,在室温下用4%多聚甲醛固定7 d 后,梯度乙醇溶液脱水,石蜡包埋,将组织切成4 μm 切片。切片用苏木精染色2 min,并用伊红染色30 s。光学显微镜下观察海马组织的病理学变化。
3.6 TUNEL 染色检测海马神经元凋亡 取方法3.5 制备的脑组织切片,去石蜡并重新水化。用0.1 mol/L PBS 洗涤2 次后,将切片与10 mg/L 蛋白酶K 工作溶液(pH 7.5~8.0)在37 ℃孵育15 min。然后,再次用PBS 冲洗,在室温下用绿色荧光素标记的dUTP溶液染色10 min。DAPI染核。使用荧光显微镜检测呈绿色荧光颗粒的TUNEL 阳性细胞。选择3个随机视野,计算TUNEL 阳性凋亡神经元占DAPI 染色神经元总数的百分比,表示凋亡率。
3.7 免疫组化法检测海马组织BDNF 的表达 取方法3.5 制备的切片,二甲苯脱蜡,PBS 清洗3 次,微波修复15 min,加入3% H2O2孵育10 min,再次清洗后,用10%山羊血清封闭30 min,加Ⅰ抗(兔抗大鼠BDNF 抗体,1∶500),4 ℃孵育过夜,然后在室温下加Ⅱ抗(1∶2 000)孵育1 h,DAB显色,苏木素复染,常规脱水、透明、中性树脂封片,显微镜下观察到胞质内呈棕黄色为BDNF 阳性细胞。每张切片随机选择不重叠的3 个视野,Image-Pro Plus 6.0 对图像进行分析,计算BDNF 阳性表达的平均吸光度(mean absorbance)。
3.8 RT-qPCR 检测海马组织miR-134 表达及Sirt1、CREB 和BDNF 的mRNA 表达 每组剩余6 只大鼠,处死后分离海马组织,使用Trizol 试剂提取总RNA。分光光度计测量总RNA 的浓度。使用PrimeScript™RT试剂盒将RNA反转录为互补DNA(cDNA)。根据扩增试剂盒的说明,进行RT-qPCR。反应体系(20 μL):SYBR®Premix Ex TaqTM(2×)10 μL,上、下游引物各0.8 μL,cDNA(200 μg/L)2 μL,ddH2O 6.4 μL。反应条件:95 ℃预变性1 min;95 ℃变性18 s,60℃持续55 s,最后72 ℃延伸2 min,进行40 个循环。所有反应均重复3次。将RNA 的表达量采用2-ΔΔCt方法计算,并相对于内参照U6或β-actin进行标准化。
3.9 Western blot 检 测 海 马 组 织Sirt1、CREB 和BDNF的蛋白表达 取海马组织,在含有蛋白酶抑制剂(PMSF)和磷酸酶抑制剂的RIPA 裂解液中匀浆,12 000 r/min离心15 min,取上清,BCA 法进行蛋白定量,RIPA 裂解液平衡蛋白浓度,变性后取等量蛋白质样品上样(30 μg),SDS-PAGE 分离,湿转法转膜,5%脱脂奶粉封闭,加入Ⅰ抗(Sirt1、CREB、p-CREB和BDNF 抗体,1∶1 000;β-actin 抗体,1∶2 000),4 ℃下孵育过夜,加Ⅱ抗(1∶5 000)室温孵育1 h,ECL 显色,以β-actin 为内参照,通过与内参的灰度比,得出目的条带的相对表达水平。
4 统计学处理
所得数据采用SPSS Statistics 22.0 和Image-Pro Plus 6.0 软件进行分析。数据分布的正态性和方差齐性分别通过Kolmogorov-Smirnov 检验和Levene 检验进行分析。数据以均数±标准差(mean±SD)表示。多组间均数比较采用单因素方差分析,并使用Bonferroni 进行两两比较。以P<0.05 为差异有统计学意义。
结 果
1 各组大鼠神经功能缺损评分
MCAO 术后,造模大鼠表现出明显的神经功能缺损,与sham 组相比,CI/R 组大鼠神经功能缺损评分显著升高(P<0.05);与CI/R 组相比,ATX-L 和ATX-H 组大鼠神经功能缺损评分显著降低(P<0.05),且ATX-H 组的效果优于ATX-L 组;与ATX-H组相比,ATX+EX527 组大鼠神经功能缺损评分显著升高(P<0.05),见表2。

表2 各组大鼠神经功能缺损评分Table 2. Neurological deficit scores of the rats in each group(Mean±SD. n=12)
2 各组大鼠学习及认知能力
与sham 组相比,CI/R 组大鼠在MCAO 术后第2~4 天,在定位航行实验中的逃避潜伏期显著延长(P<0.05),并且在空间探索实验中目标象限停留时间和穿越平台次数显著减少(P<0.05);与CI/R 组相比,ATX-L 和ATX-H 组大鼠的逃避潜伏期显著缩短(P<0.05),目标象限停留时间和穿越平台次数显著增加(P<0.05),且ATX-H 组的效果优于ATX-L 组;与ATX-H 组相比,ATX+EX527 组大鼠的逃避潜伏期显著延长(P<0.05),目标象限停留时间和穿越平台次数显著减少(P<0.05),见表3、4。

表3 定位航行实验中逃避潜伏期Table 3. Escape latency in positioning navigation experiment(s. Mean±SD. n=12)
3 各组大鼠海马组织病理学变化
HE 染色结果显示,sham 组海马组织学正常,神经元淡染,细胞核圆形;与sham 组相比,CI/R 组呈现核固缩、神经元数量减少和神经元萎缩等组织病理学变化;与CI/R组相比,ATX-L和ATX-H组大鼠海马神经元数量增加,神经元萎缩得到缓解,且ATX-H组海马组织病理损伤较轻;而ATX+EX527 组海马神经元数量较ATX-H 组明显减少,病理损伤加重,见图1。

Figure 1. HE staining results of hippocampal CA1 region of the rats in each group. A:sham group;B:CI/R group;C:ATX-L group;D:ATX-H group;E:ATX+EX527 group. The scale bar=20 μm.图1 各组大鼠海马CA1区HE染色结果
4 各组大鼠海马神经元凋亡情况
与sham 组相比,CI/R 组海马神经元凋亡率显著升高(P<0.05);与CI/R 组相比,ATX-L 和ATX-H 组海马神经元凋亡率显著降低(P<0.05);与ATX-H 组相比,ATX+EX527 组海马神经元凋亡率现在升高(P<0.05),见图2、表5。

Figure 2. TUNEL(green)staining results of hippocampal tissues of the rats in each group. A:sham group;B:CI/R group;C:ATXL group;D:ATX-H group;E:ATX+EX527 group. The scale bar=20 μm.图2 各组大鼠海马组织TUNEL染色结果
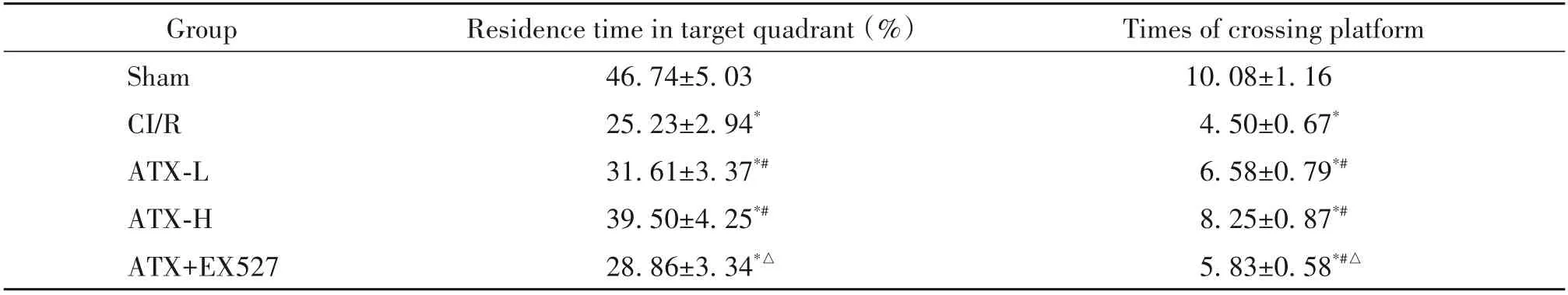
表4 空间探索实验中目标象限停留时间和穿越平台次数Table 4. Residence time in target quadrant and times of crossing platform in space exploration experiment(Mean±SD. n=12)
5 各组大鼠海马组织BDNF的表达
与sham 组相比,CI/R 组大鼠海马组织BDNF 阳性表达显著降低(P<0.05);与CI/R组相比,ATX-L和ATX-H 组大鼠海马BDNF 阳性表达显著升高(P<0.05);与ATX-H 组相比,ATX+EX527 组大鼠海马BDNF阳性表达显著降低(P<0.05),见图3、表5。

表5 各组大鼠海马神经元凋亡和BDNF的表达Table 5. Neuronal apoptosis and BDNF expression in the hippocampus of rats in each group(Mean±SD. n=6)

Figure 3. Immunohistochemical staining results of BDNF in hippocampal CA1 region of the rats in each group. A:sham group;B:CI/R group;C:ATX-L group;D:ATX-H group;E:ATX+EX527 group. The scale bar=50 μm.图3 各组大鼠海马CA1区BDNF免疫组化染色结果
6 各组大鼠海马组织miR-134 表达及Sirt1、CREB和BDNF的mRNA表达水平
与sham 组相比,CI/R 组大鼠海马组织miR-134表达水平显著升高(P<0.05),Sirt1、CREB 和BDNF的mRNA 表达水平显著降低(P<0.05);与CI/R 组相比,ATX-L 和ATX-H 组大鼠海马组织Sirt1、CREB 和BDNF 的mRNA 表达水平显著升高(P<0.05),miR-134 表达水平显著降低(P<0.05);与ATX-H 组相比,ATX+EX527组大鼠海马组织miR-134表达水平显著升高(P<0.05),Sirt1、CREB 和BDNF 的mRNA 表达水平显著降低(P<0.05),见表6。

表6 各组大鼠海马组织miR-134表达及Sirt1、CREB和BDNF的mRNA表达Table 6. The expression of miR-134,and the mRNA expression of Sirt1,CREB and BDNF in hippocampal tissues of the rats in each group(Mean±SD. n=6)
7 各组大鼠海马组织Sirt1、CREB、p-CREB 和BDNF的蛋白水平
与sham 组相比,CI/R 组大鼠海马组织Sirt1 和BDNF 蛋白表达及p-CREB/CREB 比值显著降低(P<0.05);与CI/R 组相比,ATX-L 和ATX-H 组大鼠海马Sirt1和BDNF 蛋白表达及p-CREB/CREB 比值显著升高(P<0.05);与ATX-H 组相比,ATX+EX527 组大鼠海马Sirt1 和BDNF 蛋白表达及p-CREB/CREB 比值显著降低(P<0.05),见图4。

Figure 4. Protein levels of Sirt1,CREB,p-CREB and BDNF in hippocampal tissues of the rats in each group. A:sham group;B:CI/R group;C:ATX-L group;D:ATX-H group;E:ATX+EX527 group. Mean±SD. n=6.*P<0.05 vs sham group;#P<0.05 vs CI/R group;△P<0.05 vs ATX-H group.图4 各组大鼠海马组织Sirt1、CREB、p-CREB和BDNF蛋白水平
讨 论
ATX 是一种具有高抗炎和抗氧化活性的天然类胡萝卜素,广泛分布于藻类、蟹、虾、鲑鱼和甲壳类动物中,被认为是一种潜在的神经保护药物,可减轻I/R 所致的脑损伤[6];其预处理可呈剂量依赖性地抑制急性脑梗死后的氧化应激,并上调神经营养因子(如BDNF、NGF)的表达,减轻神经损伤[13];可促进创伤性脑损伤小鼠恢复认知功能[15]。研究显示,单次或多次摄入ATX后会积聚在大鼠大脑的海马和大脑皮层中,有利于认知功能的维持和改善[16-17]。在本研究中,ATX 表现出类似的神经保护作用,可抑制海马神经元凋亡,极大地促进了I/R 大鼠的认知功能恢复。说明ATX可改善CI/R后认知功能障碍。
BDNF在海马组织中广泛表达,在突触的正常生长、发育和可塑性中起重要作用[9]。CREB 家族是BDNF 转录的主要调节剂,敲减CREB将降低BDNF的转录[18];而CREB/BDNF 通路的激活可减轻海马神经元损伤,改善认知功能[11]。在本研究中,I/R 大鼠海马组织中p-CREB/CREB 比值和BDNF 表达显著低于假手术大鼠,即CREB 磷酸化被抑制。因此,我们猜想ATX 对认知功能的改善作用可能与CREB/BDNF通路的激活有关。研究结果也证实了此猜想,经ATX 干预的大鼠海马组织中p-CREB/CREB 比值和BDNF 表达明显升高,说明ATX 可激活CREB/BDNF 通路。此结果提示ATX 可能通过激活CREB/BDNF 通路减轻CI/R 后认知功能障碍,但是尚不清楚介导CREB/BDNF表达的上游信号传导通路。
随着分子技术的发展,研究者发现miRNA 参与海马突触可塑性,在学习和记忆形成中具有显著潜力。许多miRNA 已经从神经系统中分离,据报道,miR-134 在缺血性中风中其表达明显增加,对CREB的翻译有限制作用,可通过参与树突棘大小变化、神经元可塑性等过程来影响脑发育[19];抑制其表达可保护缺血性中风小鼠的大脑和神经元免受缺氧损伤[20];并可通过靶向促进CREB mRNA 翻译,从而上调BDNF 的表达[21]。因此,miR-134 可能是CREB/BDNF 通路的上游调节因子。此外,有研究发现miR-134 在海马的过度表达,与Sirt1缺失的效果非常相似,Sirt1缺失可通过miR-134 减弱CREB 活性,下调BDNF 的表达,损害大脑的认知功能[12]。白藜芦醇可通过Sirt1/miR-134 信号通路在体内和体外调节海马中的CREB/BDNF 表达,预防慢性不可预测的轻度应激引起的认知功能障碍[22]。本研究结果显示,在I/R 大鼠海马组织中Sirt1 的表达减少,而miR-134 呈高表达,与以上研究结果一致;由此可见,Sirt1/miR-134 通路可能是CREB/BDNF 通路的上游信号传导通路。最近的研究数据表明,ATX 可以调控Sirt1蛋白表达[23],它通过激活Sirt1,减轻小鼠颅脑损伤后的氧化应激和神经元凋亡[24];还可上调BDNF和海马突触蛋白的表达,从而改善衰老模型小鼠的学习、认知和记忆能力[25]。在本研究中,可观察到经ATX 干预的大鼠海马中Sirt1 mRNA 和蛋白表达明显高于I/R模型大鼠,miR-134表达也降低了,故我们推测ATX 对CI/R 后CREB/BDNF 通路的调控作用可能与Sirt1/miR-134 信号通路有关。为了验证此推测,我们使用Sirt1 特异性抑制剂EX527 与ATX 共同干预大鼠,结果显示ATX 对CI/R 后认知功能的改善作用和对CREB/BDNF 通路的激活作用明显被减弱,提示ATX 可能通过激活Sirt1 下调miR-134 的表达,进而激活CREB/BDNF 通路,然后对大鼠CI/R后认知功能发挥保护作用。
综上所述,ATX 预处理可能通过调控Sirt1/miR-134 通路而激活CREB/BDNF 通路,进而减轻大鼠CI/R 后的认知功能障碍。本研究仅从动物水平上进行了初步探究,下一步将采用体外细胞实验对此结论进行深入验证。
