小麦赤霉病新抗源的发掘与抗性位点的检测分析
2021-10-18翟文玲刘彩云刘颖付必胜蔡瑾郭炜张巧凤吴纪中
翟文玲,刘彩云,刘颖,付必胜,蔡瑾,郭炜,张巧凤,吴纪中
江苏省农业科学院种质资源与生物技术研究所,江苏省农业生物学重点实验室,南京210014
小麦赤霉病(Fusarium head blight,FHB)是由禾谷镰刀菌(Fusarium graminearum)引起的世界性小麦重要穗部真菌病害,其发生不仅严重影响小麦产量和面粉品质,而且能产生脱氧雪腐镰刀菌烯醇(deoxynivalenol,DON)等毒素,严重危害人畜健康[1]。在我国,小麦赤霉病正从长江中下游冬麦区和东北春麦区等常发、重发区域向黄淮麦区、北方麦区等扩展[2-3]。除长江中下游麦区外,我国大部分麦区主栽品种赤霉病抗性较差。与化学防治相比,培育和推广抗病品种是防治小麦赤霉病最经济有效且环境友好的途径。
目前,全世界约有7万份普通小麦及其近缘种属种质在人工接种或自然发病条件下被鉴定,约7 000份种质呈现不同程度的赤霉病抗性[4]。我国历时10余年,筛选出抗性强而稳定的著名抗性资源苏麦3号、望水白及稳定中抗种质温州红和尚、翻山小麦等[5]。美国鉴定出Haynes Bluestem、Preston、Glyndon Fife等中抗品种[6-9]。此外,韩国品种Chokwang[10]、日本品种Nyu Bai[11]、CIMMYT人工合成小麦[12]、阿根廷品种Barletta、瑞士品种Arina[13]、智利品种Praag 8[14]及巴西品种Polyssu、Encruzilhada、Toropi和Frontana[15]也被报道具有不同程度的赤霉病抗性。除普通小麦外,二倍体和四倍体小麦、偃麦草、大赖草、披碱草、纤毛鹅观草、鹅观草、新麦草、黑麦等小麦近缘种属中均存在一些抗赤霉病资源[16-21]。
小麦赤霉病抗性是多基因控制的数量性状,目前已经从普通小麦及其近缘种属中定位了约432个赤霉病抗性位点,覆盖小麦21条染色体上44个染色体区段[4]。已命名的抗赤霉病基因或位点有Fhb1~Fhb7[[22-29]。其中,Fhb1[30-31]和Fhb7[32]已经被克隆。由于Fhb1在不同环境和遗传背景能够稳定发挥作用,当前世界范围内均以Fhb1为主要抗源开展小麦赤霉病抗性改良工作,抗源相对单一[3]。因此,发掘新的赤霉病抗源仍然是小麦抗赤霉病研究和育种的迫切任务。本研究于2017—2021年连续四年采用单花滴注法,对642份小麦种质资源的赤霉病抗扩展性(typeⅡ)进行了鉴定,以扬麦158(中抗赤霉病)为对照,筛选赤霉病抗性种质;同时利用Fhb1~Fhb7基因分子标记或连锁分子标记对抗性种质进行检测,分析抗性种质携带的抗赤霉病基因,为小麦抗赤霉病研究及抗病育种提供新的抗源及种质。
1 材料与方法
1.1 供试材料
在弥雾保湿条件下对642(田间自发赤霉病抗性表现为中抗或中感以上)份种质的赤霉病抗扩展性进行鉴定,包括育成品种211份、高代品系361份、异染色体系16份、地方品种24份和引进品种30份。其中,育成品种分别来自北部冬麦区(61份)、黄淮冬麦区(36份)、长江中下游冬麦区(60份)、西南冬麦区(54份);高代品系分别来自北部冬麦区(93份)、黄淮冬麦区(33份)、长江中下游冬麦区(111份)、西南冬麦区(124份)。以苏麦3号为抗病对照,扬麦158为中抗对照,安农8455为感病对照。其中,扬麦158曾是我国长江中下游麦区的主栽品种和骨干亲本。
1.2 赤霉病抗扩展性的鉴定方法
试验于2017—2018年、2018—2019年、2019—2020年和2020—2021年连续四年将642份小麦种质种植于江苏省农业科学院六合动物科学基地弥雾保温大棚内,行长0.6 m,行距0.25 m。每隔100份种质设置一组对照,依据“NY-T 1443.4-2007小麦抗病虫性评价技术规范”,采用单花滴注法,即用连续注射器于小麦开花期将10 μL混合菌株(F0609、F0301、F0980和F1126)孢子液注入刚开花穗中部的小花中,接种浓度为10倍显微镜视野下100个孢子。每份材料接种15~20个穗子。接种后弥雾保湿,21 d后调查总小穗数和病小穗数,按照以下公式计算病小穗率。并以病小穗率与对照相比,评价各参试材料的抗扩展性。
病小穗率(%)=(病小穗数/总小穗数)×100。
1.3 小麦基因组DNA提取与分子标记分析
取连续3年抗性鉴定为中抗以上(优于扬麦158)种质的幼嫩叶片置于2.0 mL离心管中,用CTAB法提取小麦基因组DNA。用Fhb1~Fhb7的分子标记进行鉴定,明确不同抗性种质携带的抗病基因。分子标记信息详见表1。

表1 小麦抗赤霉病基因/QTL连锁分子标记名称及染色体位置Table 1 Molecular markers of gene/QTL for wheat Fusarium head blight used in the present study
1.4 数据统计与分析
采用Excel 2010对数据进行统计分析,用SPSS 19.0进行相关分析。
2 结果与分析
2.1 相关分析
不同年份间赤霉病病小穗率的相关性均达到极显著水平(表2)。其中,2017—2018年与2019—2020年病小穗率的相关性最高,为0.41;其次为2018—2019年 与2019—2020年,相 关 系 数 为0.31;2020—2021年与其他年份病小穗率的相关性最低。由此表明,赤霉病抗性在不同年份间相关系数较低,需经过多年多点鉴定才能对种质的赤霉病抗性做出准确评价。

表2 不同年份642份小麦种质赤霉病病小穗率的相关性Table 2 Correlation coefficients among environments for percentage of symptomatic spikelets per spike(PSS)of Fusarium head blight of 642 wheat lines
2.2 病小穗率的变异
如表3所示,2017—2018年赤霉病病小穗率变幅最大,平均值也最高(18.41%);2019—2020年病小穗率变幅最小,平均值也最低(10.53%)。2019—2020年病小穗率变异系数最高,达63.90%;2020—2021年病小穗率变异系数最小,为41.99%。结果表明不同种质之间病小穗率差异数大,从高抗种质到高感种质之间病小穗率差异在50%以上即半个麦穗以上。
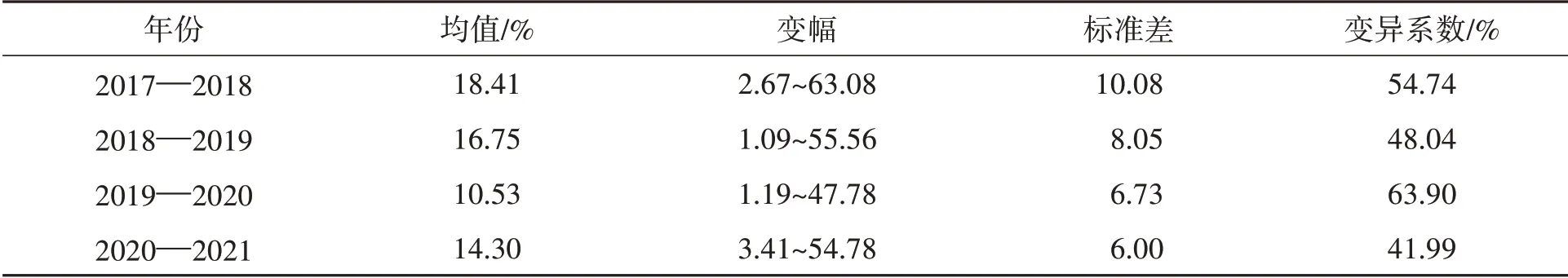
表3 不同年份642份小麦种质的赤霉病病小穗率变异Table 3 Variation in percentage of symptomatic spikelets per spike(PSS)of Fusarium head blight in 642 wheat lines among environments
2.3 抗赤霉病种质的筛选
经过2017—2021年连续4年的赤霉病抗扩展性鉴定,结果(表4)表明,3年及以上赤霉病抗性优于中抗对照扬麦158的种质有81份,包括育成品种33份,高代品系38份,地方品种5份,引进种质5份,大部分抗性种质来自于长江中下游麦区;连续4年抗性优于扬麦158的种质有33份,分别为CD0151268-1、Cp06-57-5-1-2-1F9、Grandin、光明麦1319、浩麦1号、冀师7225-28、剑子麦、克旱10号、南农13Y110、宁麦9号、农林20、农林26、软秆洋麦、瑞华14405、石优17、苏麦2号、苏州8921、望水白、武农6号、湘1022、新中长、扬11-125、扬11G4、扬12-126、扬12-149、扬12-24、扬麦1、扬麦20、扬麦4号、长江8863、镇7495、郑州5(37)和资0821。三年及以上抗性与苏麦3号相当的种质有9份,分别为望水白、Grandin、浩麦1号、剑子麦、魁小麦、农林26、软秆洋麦、苏麦2号和武农6号(表4;图1);连续4年抗性与苏麦3号相当的种质有4份,分别为剑子麦、软秆洋麦、苏麦2号和Grandin。

图1 部分小麦种质赤霉病抗扩展性鉴定Fig.1 Identifications of Fusarium head blight severity of several wheat lines
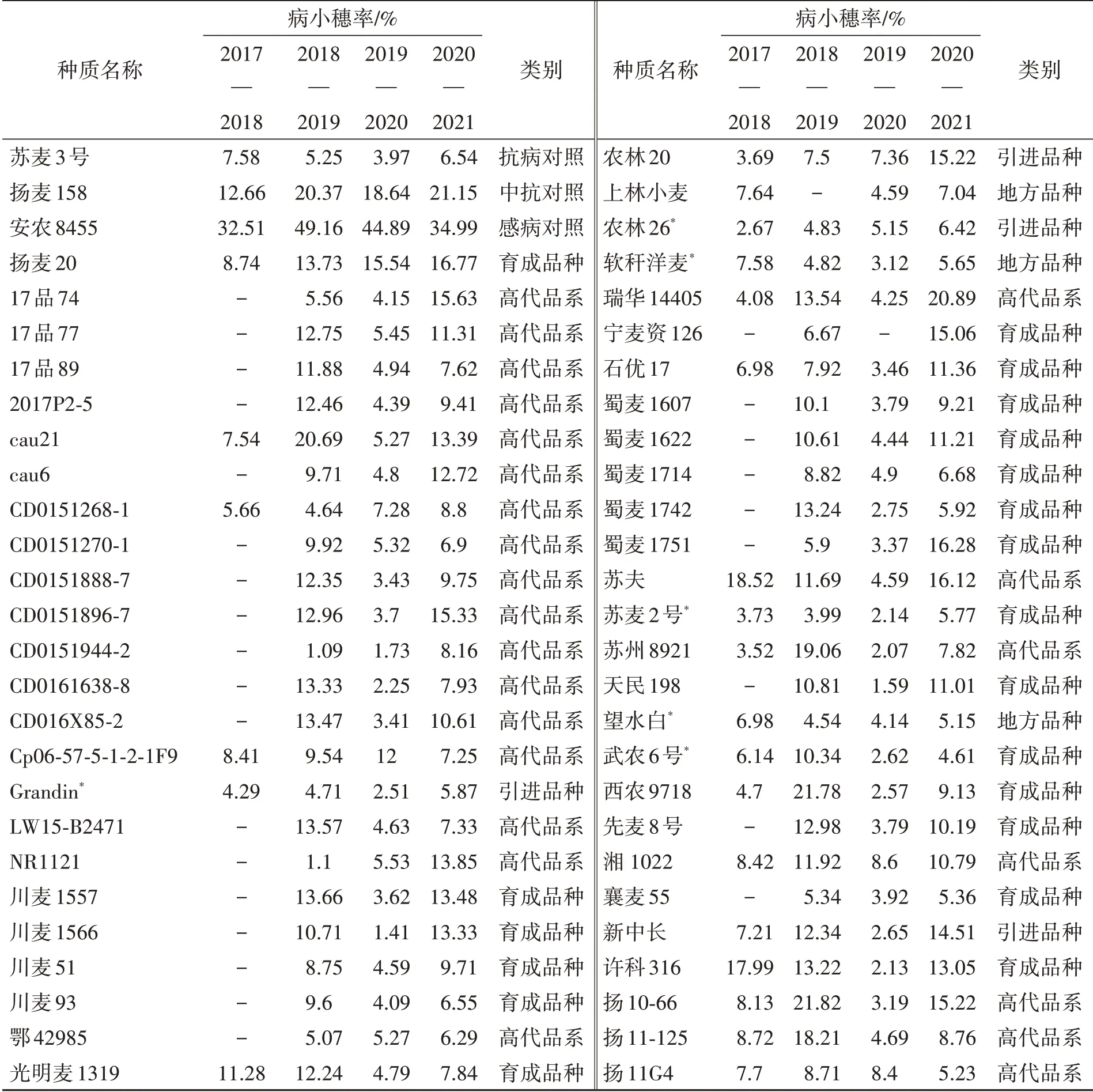
表4 三年及以上赤霉病病小穗率低于中抗对照扬麦158的种质Table 4 Wheat lined showed lower percentage of symptomatic spikelets per spike of Fusarium head blight than Yangmai 158 in at least three years

续表
2.4 抗赤霉病基因/位点的检测
为明确赤霉病中抗及以上种质所携带的抗病基因,利用已知抗赤霉病基因/位点的分子标记对这些种质进行基因型分析,结果(图2,表5)表明,对照品种中,苏麦3号携带Fhb1、Fhb2和Fhb5基因;扬麦158不携带Fhb1~Fhb7基因;抗性种质望水白携带Fhb1、Fhb2、Fhb4和Fhb5基因;苏麦2号携带Fhb1、Fhb4和Fhb5基因;农林26携带Fhb1和Fhb4基因;剑子麦和软秆洋麦携带Fhb1和Fhb5基因;Grandin携带Fhb4基因;新中长携带Fhb5基 因;而 浩 麦1号、冀 师7225-28、南 农13Y110、石优17和武农6号未检测到Fhb1~Fhb7基因,可能携带新的抗赤霉病基因。

表5 部分稳定中抗及以上(抗扩展)种质已知抗赤霉病基因型分析Table 5 Genotype of the known gene/QTL for wheat Fusarium head blight in wheat lines
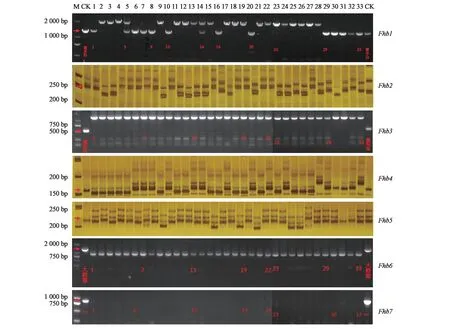
图2 部分小麦种质抗赤霉病基因/位点的基因型Fig.2 Genotypes of gene/QTL known Fusarium head blight in wheat lines
3 讨论
小麦赤霉病抗性是受环境影响较大的数量性状,其表型鉴定准确与否取决于开花期的环境温度、湿度及接种方法和赤霉菌孢子液浓度[35]。为准确评价642份小麦种质的赤霉病抗性,本研究连续4年进行鉴定。对4年的抗病性鉴定结果进行相关分析发现,2017—2018年与2019—2020年的病小穗率相关性最高;其次为2018—2019年与2019—2020年;2020—2021年与其他年份间的鉴定结果相关性最低。推测原因可能是2017—2018年小麦开花期气温较高,赤霉病发病条件充分,病害较重;而2019—2020和2020—2021年小麦开花初期(3月底4月初)气温较低,异常天气较多,赤霉病发病不充分。
近年来,小麦抗赤霉病研究已经取得巨大进展,Fhb1和Fhb7的成功克隆为小麦赤霉病抗性的遗传解析奠定了基础。Fhb1贡献率介于15%~30%[2],是主效基因,在不同环境与遗传背景中稳定表现,与其他抗赤霉病基因间存在加性效应,因此,将多个效应大且稳定的抗性基因进行聚合、选育高抗赤霉病的小麦新品种是实现小麦抗赤霉病的有效途径[33-36]。本研究利用Fhb1~Fhb7的分子标记对中高抗种质进行基因型分析发现,农林20、苏麦3号、苏麦2号、农林26、软秆洋麦、剑子麦和望水白都含有1~4个抗性基因,因此,具有较好的赤霉病抗性。然而,抗病基因通常与不利的农艺性状连锁[33],这些种质的农艺性状普遍较差,在育种利用中还需进一步改良。
尽管小麦抗赤霉病育种取得了一定的进展,但利用抗源相对比较单一,只依赖一个或少数抗性基因,导致遗传基础日益狭窄。因此,挖掘新的赤霉病抗源对小麦抗赤霉病育种具有重要意义。本研究表明,石优17、武农6号、南农13Y110、冀师7225-28、瑞华14405、浩麦1号和扬麦158不携带Fhb1~Fhb7基因,推测可能含有新的抗赤霉病基因。Zhang等[36]利用扬麦158和郑麦9023构建作图群体将赤霉病抗性基因定位于小麦2DS和3AL染色体上。且石优17、武农6号、南农13Y110、冀师7225-28和扬麦158的农艺性状较好,未来在小麦抗赤霉病研究和育种中有极大的利用潜力。
经过多年抗赤霉病育种,我国长江中下游麦区的赤霉病得到有效控制,但是近年来赤霉病逐年南迁北移,黄淮麦区也开始受到赤霉病威胁。众所周知,黄淮麦区是我国小麦最重要的产区,亩产量在20世纪90年代初就已经达600 kg以上[37],但该麦区缺乏可供育种利用的赤霉病抗源。育种家曾利用长江中下游麦区的赤霉病抗源改良黄淮麦区品种,但由于两个麦区小麦品种生育期与农艺性状的差异,杂交后代分离严重,难以选育出抗赤霉病、农艺性状又好且适宜黄淮冬麦区种植的品种[38-41]。本研究在2017—2021年对642份种质的赤霉病抗性进行鉴定,筛选出5份黄淮麦区赤霉病抗性达到中抗以上材料(石优17、郑州5(37)、冀师7225-28、武农6号和西农9718),有效弥补了黄淮麦区赤霉病抗性种质极度匮乏的现状。其中,石优17和冀师7225-28农艺性状较好,不携带Fhb1~Fhb7基因,推测可能含有新的抗赤霉病基因,为黄淮麦区小麦抗赤霉病育种提供了新的可利用抗性资源。
