不同红光与远红光比值对盐胁迫下番茄光合能力的影响
2021-10-14高登国邹志荣鲍恩财孟力力李士照
高登国,于 捷,邹志荣,曹 凯,鲍恩财,孟力力,李士照,叶 林
(1. 宁夏大学农学院,银川 750021;2. 西北农林科技大学园艺学院,农业部西北设施园艺工程重点实验室,杨凌 712100;3. 江苏省农业科学院农业设施与装备研究所,农业部长江中下游设施农业工程重点实验室,南京 210014;4. 苏州普兆农业科技有限公司,苏州 215000)
番茄(Solanum lycopersicumL.),即西红柿,广泛种植于世界各地,是菜篮子中主要的蔬菜,深受各国人民的喜爱,同时还是研究者进行遗传学及抗逆生理生化研究所用的重要模式植物[1]。近年来,我国设施番茄栽培面积迅速扩大,然而受温室半封闭环境条件和连年重茬种植、不合理灌溉、过量施肥等高度集约化经营方式的影响,土壤次生盐渍化严重,土壤中积累了大量盐分,使得设施番茄受到盐胁迫,最终导致减产及品质的降低[2]。目前,制约我国设施番茄高效、优质生产的重要原因是盐胁迫[3]。盐胁迫使番茄的光合机构受到损害[4],阻碍了光合电子的传递,严重伤害了番茄的生长发育[5]。因此,从改善设施番茄盐胁迫条件下的光合能力入手,可以有效地提高设施番茄的抗盐能力,对设施番茄抗盐高效栽培具有很重要的意义。
光是调节植物生长发育的重要环境因子,不同光质对植物生长发育也会产生不同的影响,并且与露天大田环境相比,设施温室内的光质会发生较大的变化,因此通过补光措施来改变温室内的光环境,最终达到调节植物生长的效果[6]。不同光源补光能显著改善设施番茄幼苗形态以及提高番茄幼苗全株的干鲜质量。王洪安[7]的研究表明, 温室番茄人工补光可促进番茄提前成熟期,畸形果较少,番茄植株形态健壮,品质优良。因此,根据番茄对光质的需求,提供最适宜其生长的光环境参数,可提高LED光源的利用效率。当前,对设施番茄生产中的LED光源重点在不同光质配比对番茄的光合、形态指标和生理指标等影响的研究。杨俊伟等[8]研究表明,不同红蓝光比可以促进番茄幼苗的鲜质量、叶绿素含量、根系活力等提高;张现征等[9]研究结果显示,不同红蓝光比有利于番茄幼苗干物质的积累和壮苗指数,而且有利于光合性能的提高。通常红光是指 655 ~ 665 nm 波长的光,是植物叶绿素吸收较多的光,远红光是指725~735 nm 波长的光,为植物光合提供辐射能以及作为信号调节植物整个生理过程[10]。目前已有很多研究表明,远红光和红光不仅可以促进番茄幼苗的生长[11-12],调节番茄开花时间、叶绿素含量和光合速率,还能提高番茄产量和品质[13-14]。然而,目前对红光与远红光调节设施番茄抗逆的研究却较少。鉴于此,本试验通过调节温室光环境中LED的R和Fr比例,探究在盐胁迫下,不同R:Fr比值对番茄幼苗生长的影响,以期为不同R:Fr比值的光质调控在设施蔬菜生产中的应用提供理论依据。
1 材料与方法
1.1 试验地点及材料
本试验开展时间为2016年4月―2019年11月,开展地点为江苏省农业科学院农业农村部长江中下游设施农业工程重点实验室以及西北农林科技大学农业农村部西北设施园艺工程重点实验室。供试温室番茄品种为‘MoneyMaker’。
1.2 试验处理
利用去离子水浸泡番茄种子3~5 h后,将番茄种子放在4层纱布的培养皿中并至于25 ℃恒温培养箱中催芽48~72 h,将露白后的番茄种子播于72孔塑料穴盘中,移入人工气候室进行培养,白/昼两个阶段培养时间各为12 h,其中白天光照强度为80 ~120 μmol·m-2·s-1,温度为25 ℃,夜间温度为20 ℃,湿度为65%~70%。番茄长到2叶1心时,将番茄根部基质冲洗干净后,移栽至水培槽中,进行下一步的深液流水培养。每20 min循环1次水槽中的营养液,以15 cm×10 cm作为番茄幼苗的水培行株距。白天光照为12 h,温度为26~28 ℃,光照强度为(200 ± 20)μmol·m-2·s-1,湿度为65%~70%,白天打开风扇使空气实现环流。夜间12 h,温度为18~20 ℃,湿度为65%。由图1可知,白色LED光源有2个波峰,分别为455 nm和570 nm,白色LED的R:Fr为 7.4,LED远红光的波峰在730 nm处。本实验对照组为白色LED光源,处理组为在对照组的基础上添加不同数量的LED远红光光源,形成了番茄幼苗生长发育的光环境中R:Fr值分别为1.2和0.8,本实验中其他光质的光谱分布和光照强度都一致。
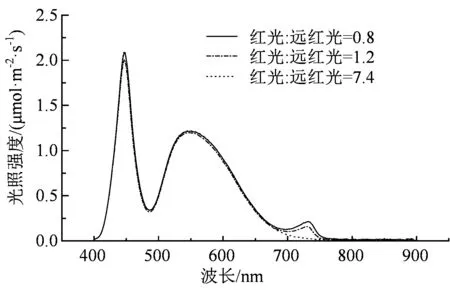
图1 LED光谱分布及光照强度Figure 1 The spectrum distribution and light intensity of LED
缓苗6 d后开始进行如下处理:1)CK,R:Fr =7.4 +正常水培;2)T1,R:Fr = 7.4 +盐胁迫;3)T2,R:Fr = 1.2 +盐胁迫;4)T3,R:Fr = 0.8 +盐胁迫。其中,正常水培:1/2山崎营养液(pH值6.2 ~ 6.5),盐胁迫处理:在1/2倍的山崎营养液中加入100 mmol·L-1NaCl的。在盐处理的同时进行不同R:Fr处理,每个处理36株番茄幼苗。
1.3 测定方法
测定LED光源:使用Model PS-100光谱辐射计(Apogee Instruments Inc, Logan, Utah, US),采用五点法测定白色和远红色LED光源组合下的不同处理番茄幼苗冠层光谱分布及光照强度。
测定番茄幼苗生长指标:待处理完成后,每个处理随机选择10个植物样本。分别用游标卡尺和直尺测量番茄幼苗的株高(测量番茄幼苗茎基部至植株生长点的长度)与茎粗(测量距离水培槽面 1 cm处的茎粗)。用去离子水小心将番茄样品冲洗干净,并擦干植株上的水分,将番茄植株根据叶、茎、根分开,分别测定其鲜重(精度为0.01 g)。测量地上、地下部的干质量时,将番茄样品装入纸袋内,放至于烘箱中75 ℃完全烘干至恒质量(约48 h),分别测定干质量(精度为0.001 g)。
测定番茄幼苗的叶面积:完成各光照处理后,随机挑选10个番茄幼苗,用直尺测量叶片的长度(Lc)和最大叶片宽度(Lk),再以经验公式分别计算单叶叶面积S[15]。公式为:S=R×(Lc×Lk),式中R为叶面积系数,当0 <Lc< 20 cm时,R取0.378 2;当Lc>20 cm时,R取0.318 4。
测定番茄幼苗的叶绿素含量及叶绿素荧光参数:分别经过0、2、4、6和8 d处理后,从每个处理中随机选择5个植物样本测定幼苗顶部向下的第3片功能叶片,用45% 酒精 + 45% 丙酮 + 10% 蒸馏水提取叶绿素,用UV-1800分光光度计测定叶绿素含量[16]。同时用Li-6800光合仪(LI-COR Inc.,Lincoln, USA)测定番茄植株叶片净光合速率设定CO2浓度为400 μmol·mol-1,光照强度为200 μmol·m-2·s-1,叶温为25 ℃,相对湿度65%~70%。在检测不同光照处理下的叶绿素荧光参数时,先将番茄幼苗置于黑暗条件中1 h,使植株充分适应光照环境的变化。随后在每个处理下,选择番茄幼苗顶部向下第3片功能叶片,利用 Li-6800 便携式光合仪检测在暗适应下每个处理叶片的最大荧光(Fm)和初始荧光(Fo)。随后将各处理组的幼苗置于光下,活化叶片1 h后检测叶片在200 μmol·m-2·s-1光强下的最小荧光(Fo')、最大荧光(Fm')与稳态荧光(Fs),每个处理5次重复。最终计算出不同处理下番茄叶片的各项荧光参数,包括:PSⅡ反应中心实际光化学效率(ФPSⅡ)、表观光合电子传递效率(ETR)、实际光化学量子产量(Fv'/Fm')、PSⅡ 最大光化学效率(Fv/Fm)、光化学淬灭系数(qP)和非光化学淬灭系数(NPQ)[17]。
测定番茄叶片光响应曲线:利用Li-6800 便携式光合仪测定番茄幼苗叶片光响应曲线,选择番茄幼苗顶部向下第3片功能叶片,将各个处理的待测叶片放至光下活化1 h,设定番茄叶片的净光合速所需CO2浓度为400 μmol·mol-1。光强度梯度设置为0、20、50、100、150、200、500、800、1 100、1 400、1 700和2 000 μmol·m-2·s-1。
1.4 数据处理与分析
试验所有数据采用 Excel 2019 进行统计分析,应用SPSS20.0进行试验数据方差及显著性分析,采用Origin 8.5进行图表的绘制。
2 结果与分析
2.1 不同R:Fr值对盐胁迫下番茄幼苗生长的影响
盐胁迫导致植物细胞渗透压增大,植物体内缺水而抑制其生长。由表1可知,与CK相比,T1(R:Fr = 7.4 +盐胁迫)处理使得番茄幼苗的叶面积、茎粗、株高与地上部及地下部的干鲜重分别降低了34.34%、28.40%、53.68%、43.28%和52.02%(P<0.05),说明番茄幼苗的生长受到显著抑制(P<0.05)。T2(R:Fr = 1.2 + 盐胁迫)处理条件下与T1处理相比番茄幼苗植株形态指标均显著升高,分别升高了30.49%、18.56%、58.17%、40.99%和48.19%,表明适当降低光环境中的R:Fr值时,盐胁迫对番茄幼苗的伤害被显著缓解(P<0.05)。T3(R:Fr = 0.8+盐胁迫)处理条件下与T1相比番茄幼苗植株形态指标均显著升高(P<0.05),分别升高了44.69%、35.73%、99.34%、71.60%和95.18%。但是,与CK相比,T3处理对番茄在盐胁迫下的生长指标差异不显著(P<0.05)。综上所述,在适宜范围内,降低番茄幼苗生长的光环境中的R:Fr值能够显著提高番茄抵抗盐胁迫的能力,促进番茄幼苗正常生长发育。
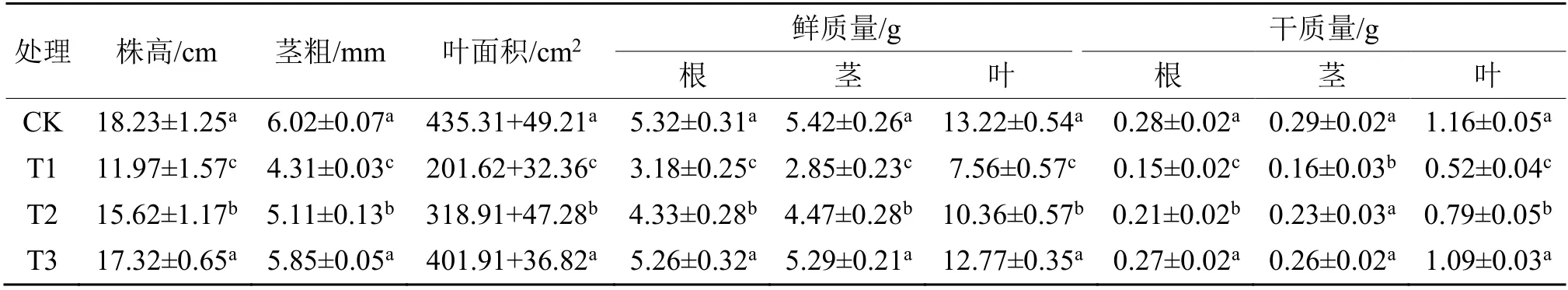
表1 不同红光远红光处理对盐胁迫下番茄植株株高、茎粗、叶面积和干鲜质量的影响Table 1 Effects of different R: Fr values on the height, stem diameter, leaf area, fresh and dry mass of tomato plants under salt stress
2.2 不同R:Fr值对盐胁迫下番茄幼苗叶片叶绿素含量和光合速率的影响
盐胁迫使得番茄幼苗叶片中叶绿素含量显著地降低,并且随着盐胁迫时间的增加,Chla和Chlb的含量逐渐下降(图2)。在盐处理并进行低R:Fr值处理的初期,会导致番茄幼苗Chla/b升高,但随着时间的推移,Chla/b逐渐降低。在盐胁迫处理8 d后,与CK相比,T1处理中的番茄幼苗叶片Chla、Chlb以及Chla/b的值都分别下降了28.89%、19.70%和12.81%。比较T3处理与T1处理时发现,T3处理的番茄幼苗叶片中,Chla、Chlb以及Chla/b的含量显著升高,分别升高了25.17%、16.51%和7.50%。综上所述,在适宜范围内,降低番茄生长环境中R:Fr值,能够显著缓解盐胁迫导致的番茄叶片叶绿素含量的下降。
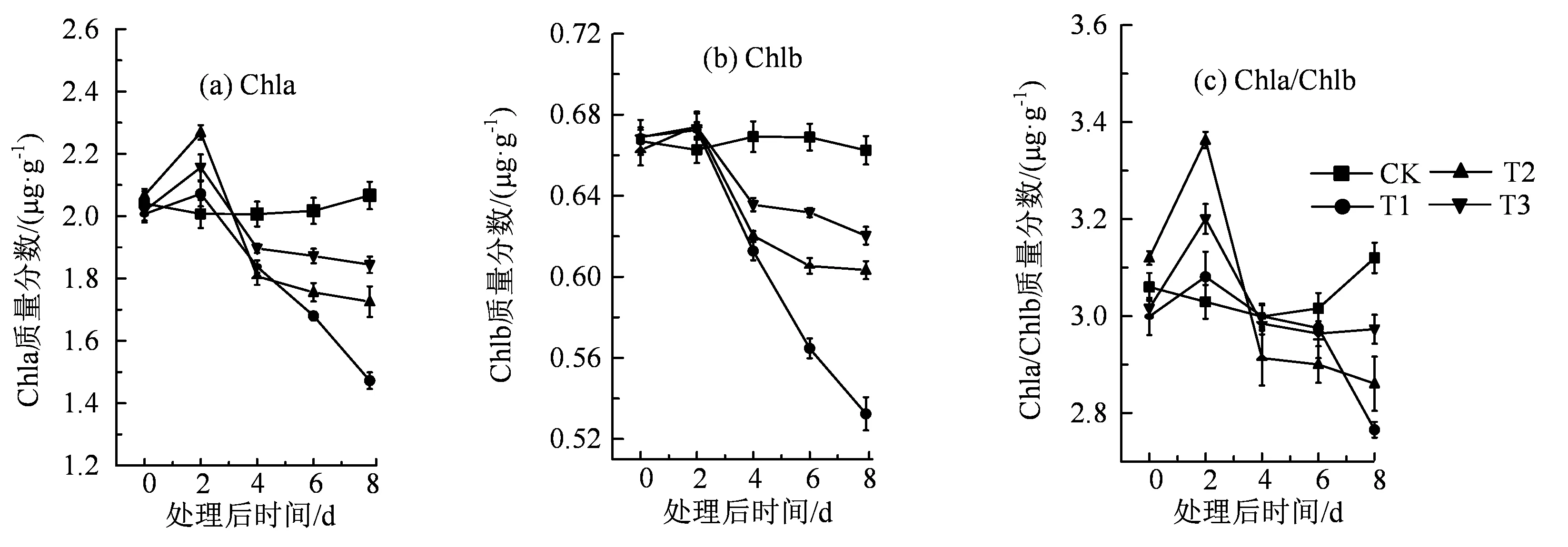
图2 不同红光远红光处理对盐胁迫下番茄植株的叶绿素a、b的影响Figure 2 Effects of different R:Fr values on the content of chlorophyll a and b in tomato plants under NaCl stress
各光照处理下番茄叶片的光合速率都被盐胁迫处理显著地影响。如图3(a)所示,在盐胁迫下,光照强度在0~200 μmol·m-2·s-1范围内时,各光照处理组中的番茄叶片净光合速率显著升高,CK、T1、T2和T3的平均净光合速率分别为8.24、3.87、6.14和7.41 μmol·m-2·s-1,差异显著(P<0.05)。但是,当光照强度达到800 μmol·m-2·s-1时,各个处理番茄叶片的净光合速率基本保持不变,CK、T1、T2和T3的平均净光合速率分别为 17.08、5.07、10.15和13.09 μmol·m-2·s-1,差异显著(P<0.05)。图3(b)和(c)中的光响应曲线表明:不同处理的光饱和点由大到小排列顺序为:CK、T3、T2和T1,即降低盐胁迫下光环境中的R:Fr比值能够显著提高番茄幼苗的光饱和点,降低番茄幼苗的盐胁迫。同时,随处理后时间的变化和光合日变化,不同光质处理下番茄幼苗的光合速率由大到小的排列顺序也为:CK、T3、T2和T1。综上所述,在盐胁迫下,适宜范围内在R:Fr值较低的光环境能够使得番茄叶片的净光合速率显著降低,提高番茄幼苗的抗盐性。
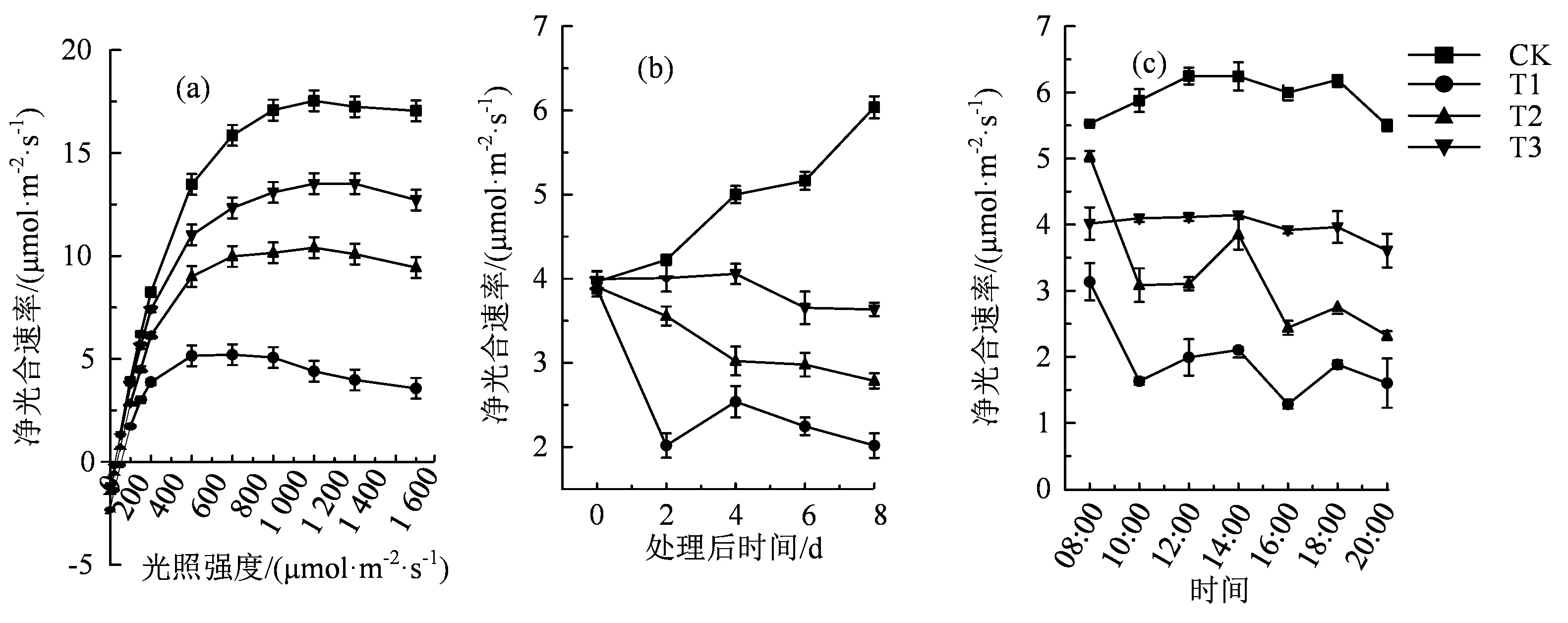
图3 不同红光远红光处理对盐胁迫下番茄植株光合作用的影响Figure 3 Effects of different R:Fr values on photosynthesis in tomato plants under NaCl stress
2.3 不同R: Fr值对盐胁迫下番茄幼苗叶片中叶绿素荧光参数的影响
盐胁迫能够导致植物细胞渗透压升高而损伤植物体内的光合机构,最终严重影响植物的正常生长。由图4可知,NaCl胁迫下,番茄幼苗叶片的Fv/Fm、ФPSⅡ、Fv'/Fm'、ETR和qP都显著降低,NPQ显著升高(P<0.05)。盐胁迫下适当降低光环境中的R:Fr值,使得能量能够快速进入PSⅡ反应中心。与T1相比,T3处理中Fv/Fm、Fv' /Fm'、ФPSⅡ、ETR和qP分别升高了12.86%、17.54%、24.00%、10.26%和 11.25%,NPQ降低了20.00%。结果表明,在适宜范围内,降低R:Fr比值的光环境能够逐渐抑制盐胁迫对番茄幼苗PSⅡ反应中心造成的损伤,从而缓解番茄幼苗的盐胁迫。
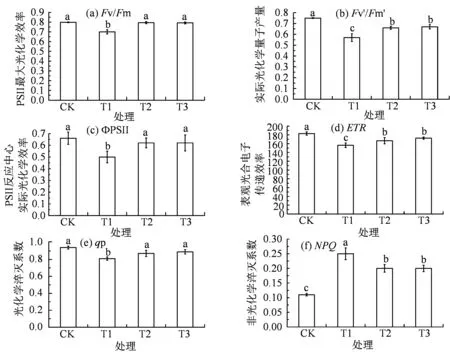
图4 不同红光远红光处理对盐胁迫下番茄幼苗叶片叶绿素荧光参数的影响Figure 4 Effects of different R: Fr values on the chlorophyll fluorescence parameters in leaves of tomato seedlings under NaCl stress
3 讨论与结论
在适宜范围内,降低植物生长光环境中的R:Fr值时,会促使植物体内感受外界光源变化的受体发生变化,即光敏色素从Pfr型转化成为Pr型[18]。当升高R:Fr值,植物矮化,茎节变小,光合产物降低;反之,当R:Fr比值降低时,则植物的茎节间距变大、徒长,光合产物降低[19-20]。此外,降低光环境中R:Fr的比值能够促进萝卜组培苗根系的生长[21]。本试验中,在120 mmol·L-1NaCl胁迫下,适当降低R:Fr比值,则番茄植株的形态指标均显著升高。当120 mmol·L-1NaCl胁迫下光环境中R:Fr比值为0.8 时,番茄植株的各生长指标与非盐胁迫下光环境中R:Fr比值为7.4时基本一致,差异不显著。研究结果表明,在适宜范围内,降低R:Fr比值能明显促进番茄地上部和地下部的生长,提高番茄植株的耐盐性。
NaCl 胁迫对番茄幼苗有抑制作用,包括对净光合速率、叶绿素含量、气孔导度、蒸腾速率和胞间 CO2浓度均有着明显的抑制作用[22]。
许多学者研究表明,叶绿素在盐胁迫下其含量逐渐降低,但也有叶绿素含量升高,且叶绿素含量与光合作用效率呈正相关关系[23]。盐胁迫下叶绿素类囊体膜受损使得合成叶绿素的含量降低,从而导致植株光合速率下降[24]。在本试验中,适当地降低光环中的R:Fr 值,盐胁迫下番茄叶片捕获光能的配比调整为对低R:Fr值最适应的状态,从而缓解番茄叶片叶绿素含量的下降,以实现对光能最大限度的捕获。Emerson等[25]研究发现,当光源红光与远红光同时照射植物,光合效率大于这两种单独照射的效率之和。这是由于光系统Ⅰ(PSⅠ)主要吸收大于680 nm的远红光,而光系统Ⅱ(PSⅡ)主要吸收680 nm的红光[26]。此外,远红光可以提高叶片的扩展度从而提高整株植物的光合能力[27]。Zhen等[28]的研究发现,增加Fr可以有效提高生菜PSⅡ反应中心光化学效率,从而提高生菜产量。秦红艳等[29]研究表明,盐胁迫下葡萄叶片的实际光化学效率、光化学淬灭系数以及表观光合电子传递速率显著降低,非光化学猝灭系数升高,而非光化学猝灭系数的升高保护了植株叶片正常进行光合,减少盐胁迫对植株的伤害。魏霞等[30]研究发现,造成植物叶绿素含量降低的原因主要是盐胁迫下类囊体膜的完整性受损,低叶绿素值能提高植物对远红光的吸收从而缓解盐胁迫。唐玲等[31]研究表明,鸡爪槭叶片的Fv/Fm、ФPSⅡ、ETR和qP随盐浓度的增大而呈下降趋势,而NPQ在低盐浓度时上升,这是缓解盐胁迫的表现。本试验中,在120 mmol·L-1NaCl胁迫下番茄叶片的Fv/Fm、Fv'/Fm'、ФPSⅡ、ETR和qP下降,NPQ升高,表明番茄叶片主要是通过增加热耗散形式来消耗过多的光能,避免120 mmol·L-1NaCl 胁迫下对光合机构造成的损害,确保番茄植株的正常生长。在120 mmol·L-1NaCl胁迫下适当降低R:Fr的比值,番茄叶片的Fv/Fm、Fv'/Fm'、ФPSⅡ、ETR和qP显著升高,同时显著抑制了NPQ。由此可见,在适宜范围内,较低的R:Fr比值能够显著提高盐胁迫下番茄幼苗PSⅡ反应中心的光化学效率。
在120 mmol·L-1NaCl胁迫下适当降低R:Fr值,能显著促进番茄幼苗的生长发育,其中根茎叶干鲜质量、叶面积、茎粗、株高以及叶片叶绿素含量、净光合速率等显著升高;叶绿素荧光参数中Fv/Fm、Fv' /Fm'、ФPSⅡ、ETR和qP显著升高,NPQ显著降低。在120 mmol·L-1NaCl胁迫下,R:Fr比值为0.8时,番茄植株的各生长指标与非盐条件下R:Fr比值为7.4时基本一致;在120 mmol·L-1NaCl胁迫下,在适宜的低R:Fr比值的光环境下可以减弱PSⅡ的损伤,促进PSⅡ的电子传递。
