干扰ELOVL6基因对奶牛乳腺上皮细胞脂代谢相关基因的表达及甘油三酯合成的影响
2021-10-14姚大为赵淑颖杨春蕾李玉鹏丁向彬
姚大为, 赵淑颖, 赵 欣, 杨春蕾, 李玉鹏, 丁向彬, 马 毅*
(1.天津市农业科学院畜牧兽医研究所,天津西青300381;2.天津农学院动物科学与动物医学学院,天津西青300384;3.南开大学生命科学学院,天津南开300071)
牛奶不仅能够提供能量,并且含有优质蛋白质、维生素和钙元素等,是国民饮食的重要营养物质来源,其丰富的脂肪酸种类及较多不饱和脂肪酸含量显现出了较高的营养价值(Demment等,2003)。研究表明,牛奶品质与脂肪酸组成和含量有着密切的关系(Chilliard等,2003)。乳脂代谢过程包括脂肪酸的从头合成、脂肪酸去饱和、脂肪酸的摄取与转运、甘油三酯的合成与降解等。因此,深入了解奶牛乳腺脂代谢调控机理,有助于为实施分子育种措施进而改善牛乳品质提供理论依据。
超长链脂肪酸延伸酶(ELOVLs)在长链脂肪酸合成过程中起关键作用,ELOVL6基因是其中的一员,主要功能是催化C12、C14、C16饱和与不饱和脂肪酸延伸为C18脂肪酸,主要参与脂肪酸延伸和脂酰辅酶A的生物合成(胡忠昌等,2018)。胡忠昌(2018)在转基因小鼠的肝脏中克隆出ELOVL6基因,全长2500bp,编码区长801 bp。ELOVL6基因广泛分布于体内的各个组织器官,在脂肪含量较高的组织或器官中高水平表达,其 余 组 织 或 器 官 表 达 量 较 低 (Li等,2019)。ELOVL6影响体外培养的小鼠肝脏细胞中C16和C18脂肪酸的比例,终产物为硬脂酸(C18:0)。硬脂酸在硬脂酰辅酶A去饱和酶1(SCD1)的作用下生成油酸(C18:1,n-9)(Matsuzaka等,2009),进而为甘油三酯的合成提供底物。在缺失ELOVL6基因的斑马鱼肝脏中SCD1表达水平显著降低,表明ELOVL6基因的缺失可能会减少斑马鱼的脂肪生成(Wang等,2020)。再娜古丽·君居列克(2020)在牛脂肪细胞中过表达ELOVL6基因显著上调了FABP3和SCD1等乳脂代谢相关基因的表达,表明ELOVL6是参与乳脂代谢的重要基因。有研究表明,处于妊娠期的鼠乳腺组织中ELOVL6基因表达量显著上调(Rodriguez-Cruz等,2011)。在人和小鼠中,雌性ELOVL6基因的表达水平高于雄性,雌二醇和孕酮可以促进ELOVL6基因的表达,调节乳腺的发育和功能维持(Schams等,1984)。综上所述,ELOVL6基因可能通过改变脂肪酸的组成在乳脂代谢中发挥重要作用。目前,有关牛乳腺上皮细胞中的ELOVL6基因在脂质代谢中的作用鲜有报道。
本研究通过克隆得到奶牛ELOVL6基因CDS区,筛选得到有效siRNA,将有效siRNA转染奶牛乳腺上皮细胞,探究其对脂质代谢相关基因表达及甘油三酯合成的影响,对进一步揭示该基因在乳腺组织脂质代谢中的调控机制,提高奶牛的生产性能及牛奶质量具有重要的指导意义。
1 试验材料
1.1 试验动物 奶牛乳腺组织cDNA由山东农业科学院馈赠;奶牛乳腺上皮细胞由扬州大学动物科学与技术学院馈赠。
1.2 主要试剂 Trizol试剂盒、LATaqDNA聚合酶、TBGreenRPremixExTaqTMII、限制性内切酶(SalⅠ和EcoRⅠ)、pMD19-T载体连接试剂盒,均购自TaKaRa公司;质粒提取试剂盒、胶回收试剂盒、DNAmarkerⅢ、核酸染料,均购自天根生化科技(北京)有限公司;DMEM/F-12培养基、胎牛血清、LipofectamineRRNAiMAX转染试剂,均购自ThermoFisher公司;甘油三酯检测试剂盒,购自北京普利莱基因技术有限公司;其他试剂均为国产。
1.3 主要仪器 Taco核酸自动提取仪(型号为T0124),购自北京华安瑞基生物科技有限公司;荧光定量PCR仪(CFX-96),购自伯乐(中国)有限公司;PCR扩增仪(型号为VeritiV255249),购自赛默飞世尔科技(中国)有限公司;多功能酶标仪(SuPerMax3100),购自上海闪谱生物科技有限公司;凝胶成像分析仪(型号为GelDocXR),购自北京众力挽生物科技有限公司;双人单面超净工作台(型号为SW-CJ-2FD),购自上海昕仪仪器表有限公司。
2 试验方法
2.1 奶牛ELOVL6基因CDS区克隆 根据GenBank中牛的ELOVL6基因(NM_001102155)序列,利用Premier5.0软件设计特异性扩增引物(F:5’-ATATGTCAGTGTTGACTTTACAAGAAT-3’;R:5’-CAGTTCCGACACTAGTCAGCTTT-3’)。引物由生工生物工程(上海)股份有限公司合成。
以奶牛乳腺组织中cDNA为模板进行PCR扩增,PCR反应体系为:PrimeSTARMaX12.5μL、上下游引物各1.0μL、cDNA模板1.0μL、无菌水9.5μL,共25.0μL体系。延伸时间为1min,退火温度为55℃,PCR产物用1%的琼脂糖凝胶电泳检测,胶回收约800bp的目的条带并与pMD19-T载体连接,构建克隆载体pMD19-T-ELOVL6,转化E.coliTop10大肠杆菌感受态细胞,挑菌,37℃摇床12h,提质粒,SalⅠ/EcoRⅠ双酶切鉴定正确后送生工生物工程(上海)股份有限公司进行测序。
2.2 ELOVL6基因生物信息学分析 利用NCBI中 的Blastn(http://www.ncbi.nlm.nih.gov/BLAST)进行奶牛与其他物种ELOVL6基因核苷酸序列的同源性比对。利用ProtParam(http://www.expasy.org/tools/protparam.html)、ExPASyProteomicsServer(http://us.expasy.org/cgi-bin/protscale.pl)和TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)对奶牛ELOVL6蛋白的相对分子质量、理论等电点、蛋白疏水性预测、跨膜结构域进行预测。
2.3 ELOVL6基因siRNA的设计与合成 利用Invitrogen公司BLOCK-iTTMRNAiExpress软件(https://rnaidesigner.invitro-gen.com/rnaiexpress/)在线设计靶向本试验克隆的奶牛ELOVL6基因的CDS区的siRNA和对照siRNA(表1),并由广州锐博生物技术有限公司进行合成。

表1 针对奶牛ELOVL6基因的siRNA序列及对照组siRNA序列
2.4 细胞培养及siRNA的转染 将奶牛乳腺上皮细胞(BMEC)置于37℃、CO2浓度为5%的培养箱中培养,培养基成分为D-MEM/F12、10%胎牛血清、5μg/mL胰岛素、10μg/mL表皮生长因子、100μ/mL青霉素、100μ/mL链霉素、1μg/mL氢化可的松。每24h换一次液,当细胞密度达到80%以上时进行转染。siRNA转染时,根据LipofectamineRRNAiMAX转染说明书配制转染复合物,避光放置20min,同时转染至对照组和试验组乳腺上皮细胞中,每组3个重复。
2.5 有效siRNA的筛选及RT-qPCR检测干扰ELOVL6基因对乳脂代谢关键基因表达量的影响 转染24h后进行换液,48h后用PBS润洗两遍,采用Trizol法提取总RNA检测浓度和纯度后反转录为cDNA,并以此为模板通过RTqPCR检测ELOVL6基因的表达量,ELOVL6基因的定量引物见表2,采用2-ΔΔct法进行相对定量计算各组siRNA的干扰效率,筛选出干扰效率最佳的siRNA。
将干扰效率最佳的siRNA转染至乳腺上皮细胞,48h后采用Trizol一步法提取总RNA并反转录成cDNA用于RT-qPCR。利用PrimerPremier5.0软件,根据NCBI已经公布的基因序列设计ELOVL6基因的定量引物,其他脂质代谢相关基因的定量引物参照表2,反应体系:TB10μL,上下游引物各0.8μL,cDNA模板2μL,RNase水6.4μL。反映步骤:95℃预变性30s,95℃5s,60℃30s,39个循环,95℃30s,65℃5s,95℃5s。每个样品设置3个重复。采用UXT、MRPL39为内参基因。通过SPSS统计软件对数据进行显著性分析,结果用“平均值±标准差”表示,P<0.05表示差异显著,P<0.01表示差异极显著。

表2 实时荧光定量PCR产物
2.6 检测干扰ELOVL6基因对奶山羊乳腺上皮细胞甘油三酯含量的影响 在乳腺上皮细胞中分别将对照组和干扰组(n=3)siRNA转染后,弃去培养基,将细胞用PBS清洗3次,加入甘油三酯检测试剂盒中的裂解液室温轻柔摇晃20min后,用细胞刮铲将细胞碎片和裂解液全部转移至离心管中进行超声破碎,根据试剂盒说明书利用酶比色法检测甘油三酯含量。细胞中的总蛋白含量使用BCA蛋白检测试剂盒进行检测。甘油三酯的总含量使用每孔对应的蛋白含量进行标定校准。
3 结果
3.1 奶牛ELOVL6基因CDS区的克隆 以奶牛乳腺组织提取的cDNA为模板,通过PCR技术进行扩增。经1%的琼脂糖凝胶电泳检测扩增产物为约800bp的片段,与预期结果基本相符(图1A)。与pMD19-T载体连接后进行双酶切鉴定(图1B),得到两条清晰的条带,其中一条与目的片段大小一致,确认连接成功并进行测序。结果通过Blast比对,确认克隆得到牛ELOVL6基因的CDS区,全长795bp,编码264个氨基酸,提交至GenBank,登录号为MW960011。
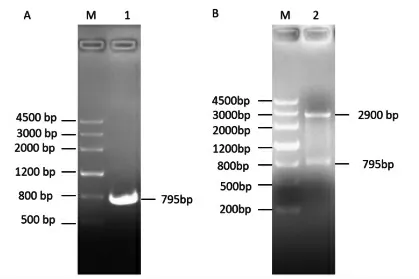
图1 ELOVL6基因的克隆与鉴定
3.2 奶牛ELOVL6基因生物信息学分析 经NCBI中Blast进行同源性分析后发现,奶牛ELOVL6基因CDS区核苷酸序列与牛(NM_001102155.1)、山 羊(NM_001314257.1)、大羚羊(XM_040237469.1)的相似度分别为100%、96.73%、96.35%。ELOVL6蛋白相对分子质量为31.28ku,理论等电点为5.94。经蛋白疏水性预测发现,该蛋白疏水最大值为2.467,最小值为-2.411,具有较强的疏水性(图2)。主要定位在内质网,具有6个跨膜区域(图3)。

图2 ELOVL6蛋白疏水性分析
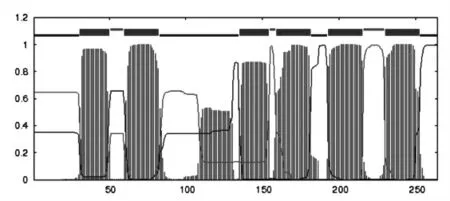
图3 ELOVL6蛋白跨膜区分析
3.3 奶牛ELOVL6基因有效siRNA序列的筛选及干扰效率检测 将3对干扰组的siRNA及对照组siRNA分别转染至乳腺上皮细胞,培养48h后,采用Trizol法提取总RNA,反转录为cDNA进行荧光定量PCR。由图4可知,3对siRNA均有显著的干扰效率,其中siELOVL6-003的干扰效率最高,达到90%以上(P<0.01)。
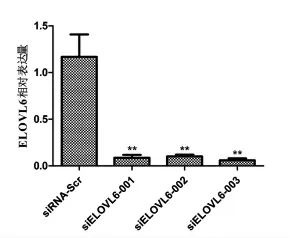
图4 奶牛ELOVL6基因siRNAs序列的干扰效率检测
3.4 干扰ELOVL6基因对脂代谢相关基因表达的影响 将筛选得到的有效siRNA转染至乳腺上皮细胞48h后,进行总RNA提取并反转录成cDNA,以此为模板通过荧光定量PCR检测脂质合成相关基因的表达情况。结果表明,脂肪酸从头合成及去饱和相关基因(ACACA、SREBF1、PPARG、SCD1)、脂肪酸转运相关基因(FABP3)、甘油三酯合成相关基因(DGAT1和AGPAT6)、甘油三酯水解相关基因(HSL)的表达量显著降低(图5)。

图5 干扰ELOVL6基因对脂代谢相关基因表达的影响
3.5 干扰ELOVL6基因对细胞内甘油三酯含量的影响 在乳腺上皮细胞中进行对照组与干扰组siRNA转染,培养48h后,弃去培养基,将细胞用PBS清洗3次,采用试剂盒进行细胞内甘油三酯含量的测定并以每孔对应的蛋白含量进行标定校准,结果表明,干扰ELOVL6基因后细胞内甘油三酯含量降低(图6)。

图6 干扰ELOVL6基因对奶牛乳腺上皮细胞中甘油三酯含量的影响
4 讨论
在哺乳动物中,脂肪酸合酶(FASN)只能催化C16以下脂肪酸的生成,而不能催化C16以上脂肪酸的延伸。ELOVL6可以将FASN合成的棕榈酸(C16:0)催化生成硬脂酸(C18:0)。ELOVL6在羊、啮齿动物中通过改变脂肪酸的合成来影响甘油三酯的合成,对ACACA、FABP3等乳脂代谢相关基因的表达有影响(吴皓,2014;吴敏,2014;Beigneux等,2006;)。然而,目前该酶在奶牛乳腺上皮细胞中是否发挥同样的作用还鲜有报道。本试验通过干扰ELOVL6基因的表达,验证了ELOVL6在奶牛乳腺上皮细胞对甘油三酯合成及相关基因表达的影响。
ACACA、SREBF1、PPARG等酶参与了脂肪酸的从头合成(Wu等,2019)。乙酰CoA羧化酶是一种生物素依赖性酶,催化乙酰CoA转化为丙二酰辅酶A(Bionaz等,2008),ACACA是其中的一个亚型,在脂肪生成组织中产生,用于从头合成脂肪酸,被认为是控制脂肪酸合成的主要亚型(Witters等,1994)。本研究结果表明,当ELOVL6基因被干扰后,ACACA表达量的下调降低了脂肪重新生成的能力,这与Harris(2011)的试验结果一致。包梅(2021)研究表明,SREBP1是人和动物的脂肪生成调控因子,通过调控参与乳脂代谢相关基因(FASN、SCD1)的表达以及转运蛋白的活性,进而影响甘油三酯的合成。吴敏(2014)研究表明,在奶山羊乳腺上皮细胞中超表达ELOVL6基因导致SREBP-1基因表达量显著下降。本试验数据显示,干扰ELOVL6基因的表达显著下调SREBP1表达量,导致其下游靶基因SCD、ACACA等的表达量也随之下调,甘油三酯合成减少,与上述试验结果一致。过氧化物酶体增殖物激活受体γ(PPARG)是核受体家族的成员,在多种组织和器官中表达,主要调控脂肪的合成与代谢(Marion-Letellier等,2016)。在奶山羊乳腺上皮细胞中的过表达ELOVL6基因对PPARG的表达量没有影响,而本研究结果中干扰ELOVL6基因表达显著下调PPARG的表达量,与上述结果不一致,原因可能是在不同的物种中脂肪酸对转录因子PPARG的调节机制不同。
有研究表明,ELOVL6可为甘油三酯合成提供基质,在山羊乳腺上皮细胞过表达ELOVL6基因后,甘油三酯浓度增加,在ELOVL6被敲除后降低(Shi等,2017),本试验与上述结果一致。干扰ELOVL6基因显著下调DGAT1、AGPAT6基因的表达量,表明ELOVL6基因或其底物参与甘油三酯的合成。研究结果表明,DGAT1与乳脂含量显著相关(Pacheco-Pappenheim等,2019;Tzompa-Sosa等,2016;Tabaran等,2015),在牛肌肉细胞中过表达DGAT1显著提高甘油三酯含量(Liu等,2007)。本试验结果表明,DGAT1酶含量的降低是导致甘油三酯含量减少的原因之一,干扰ELOVL6基因的表达使得DGATI酶含量降低,进而影响甘油三酯的含量。Massimo和Loor(2008)的研究表明,AGPAT6是小鼠、牛、羊等动物甘油三酯合成的关键酶,AGPAT6在牛哺乳期的表达显著上调。本试验数据显示,干扰ELOVL6基因显著下调AGPAT6基因的表达量。油酸(C18:1n-9)是合成甘油三酯的首选底物(Green等,2010),其是在C18:0去饱和后生成的(Stone等,2004)。硬脂酰-CoA去饱和酶-1(SCD1)负责脂肪酸的去饱和,由ELOVL6基因催化生成的硬脂酸(C18:0)经SCD1去饱和为油酸(C18:1,n-9)。SCD1和ELOVL6在甘油三酯合成过程中处于连锁反应(Marion-Letellier等,2016)。本试验研究表明,干扰ELOVL6基因显著下调SCD1基因的表达量,与上述结论一致。陈俐静(2020)认为,乳脂代谢是一个合成与分解动态平衡的过程,HSL基因将甘油三酯、甘油二酯和甘油一酯分解为游离脂肪酸和甘油,是脂肪分解过程中的限速酶。本试验结果表明,干扰ELOVL6基因显著下调HSL基因的表达量,可能是为了维持细胞内甘油三酯的稳态。
从头合成脂肪酸对甘油三酯的合成具有重要作用,而从外周循环和细胞内运输中摄取长链脂肪酸在乳脂合成中也很重要(Green等,2010)。Bionaz和Loor(2008)认为,FABP3结合长链脂肪酸将其运输到内质网,为SCD提供底物。本试验结果表明,干扰ELOVL6显著下调FABP3基因表达量,可能是由于FABP3可结合的底物长链脂肪酸含量减少,导致转运到内质网SCD1的底物含量减少,甘油三酯含量降低。
虽然本研究提供了干扰ELOVL6基因表达可以降低奶牛乳腺上皮细胞中脂肪酸合成和甘油三酯合成的试验结果,但是还是要考虑本试验的一些局限性。应该测量其中一些酶的活性,以更好地反映和量化脂肪酸去饱和及延长的速率。ELOVL6基因和与乳脂代谢相关基因之间的具体调控机制还有待进一步研究。
5 结论
本试验克隆了奶牛ELOVL6基因并对其进行了生物信息学分析。成功筛选到奶牛ELOVL6基因的有效siRNA(干扰效率达到90%以上),通过RT-qPCR检测乳脂代谢相关基因的表达,通过与对照组对比发现,干扰ELOVL6基因的表达可 显 著 下 调ACACA、SREBF1、PPARG、SCD1、FABP3、DGAT1、AGPAT6和HSL基因的表达量,细胞内甘油三酯含量显著降低。该结果为研究ELOVL6基因在奶牛乳脂合成中的调控机理奠定了基础。
