貉细小病毒RDPV-JL3株的分离鉴定及VP2基因序列分析
2021-10-14郭晓芹于小亚李滋睿李卓宸吕甜甜曹海旭宋海岩赵传芳
郭晓芹,于小亚,李滋睿,李卓宸,吕甜甜,曹海旭,宋海岩,赵传芳※
(1.中国农业科学院特产研究所,吉林 长春130112;2.吉林省科技创新平台管理中心,吉林 长春130012;3.中国农业科学院后勤服务中心,北京100081;4.吉林冠界生物技术有限公司,吉林 梅河口135000)
貉子,又名狗狸,是起源于东亚的一种本土食肉目犬科动物[1]。通过训化后人工养殖的貉子背部毛皮质量能接近于蓝狐的毛皮质量[2],使得貉皮在国际市场中的价格不断上涨,给毛皮养殖业带来巨大的经济效益。而貉细小病毒(RDPV)能引发貉细小病毒肠炎,是危害养貉业的重要传染病之一[3]。幼貉的发病率在70%以上,死亡率高达90%,成龄貉的感染率和死亡率均在30%以下,多呈慢性和隐性感染,其带毒的排泄物(粪便和尿液等)成为最危险的传染源[4]。
RDPV属于细小病毒科(Parvoviridae)细小病毒属(Parvovirinae)成员,与犬细小病毒(CPV)、猫泛白细胞减少症病毒(FPV)、水貂肠炎病毒(MEV)、浣熊细小病毒(RPV)以及蓝狐细小病毒(BFPV)的基因组同源性高达98%以上[5],肉食动物细小病毒被科学家认为是由同一始祖毒株在生态变化或自然选择等因素的影响下,突破了种间屏障,获得跨种属传播的能力后逐渐进化成的一种具有种属特异性的病毒[6-9]。1978年CPV以CPV-2型首次在犬体中被发现,CPV-2作为一种新病原其最被研究学者认可的产生途径是FPLV借助狐或貉等中间宿主变异进而感染犬出现的[10]。RDPV也被认为是CPV跨宿主感染貉产生[11]。RDPV自1984年(首次在我国黑龙江省发生)至今所分离出的毒株虽然大都属于CPV-2亚型,但从2016开始,RDPV主要衣壳蛋白VP2上的多个氨基酸残基发生了非同义性突变,如S27T、S297A和I418T,这种新型CPV-2可能是细小病毒进化的过渡阶段[12,13]。
本实验从吉林某养殖场疑似患貉细小病毒的貉肠道组织中成功分离出1株貉细小病毒,并对此分离毒株的VP2基因进行克隆测序和基因序列分析,了解该地区RDPV分子流行病学的变化趋势,以期为更好的研究貉细小病毒病的诊断、疫苗防控以及揭示CPV-2进化机制提供思路。
1 材料与方法
1.1 材料
1.1.1 病料病料来自吉林省吉林市某貉养殖场,发病貉表现为呕吐及腹泻等,临床症状疑似为RDPV感染,Anigen Rapid CPV Ag Test Kit检测粪拭子显示阳性,采集病死貉的肠组织及肠道内容物。
1.1.2 细胞及试剂猫肾传代细胞F81株、RDPV阳性血清及RDPV VP2蛋白单抗由本实验室保存;细胞培养用的胎牛血清、消化液胰酶及MEM购自Gibco公司;DNA Marker(DL5 000、DL2 000、DL1 000)基因组提取试剂盒、凝胶回收试剂盒、EasyPure Plasmid MiniPrep Kit、Peasy-T1 Cloning Kit、大肠杆菌DH-5感受态细胞等购自北京全式金生物技术有限公司;LA Taq DNA聚合酶、购自Takara公司;Goat Anti-Mouse IgG(H+L)/FITC购自北京博奥森生物有限公司;引物由库美生物科技有限公司合成。
1.1.3 仪器设备细胞培养箱、低温离心机(Thermo公司产品);凝胶成像仪(GE公司产品);组织研磨仪(Thmorgen公司产品);荧光倒置显微镜(Leica公司产品)。
1.2 方法
1.2.1 病料处理将采集的貉肠道组织及其内容物研磨充分后用PBS按1:5的比例稀释,于﹣79℃反复冻融3次,5000r/min,4℃离心45min,取上清用0.22m滤膜过滤除菌后加入双抗(Penicillin-Streptomycin,100),使其终浓度为100U/mL,4℃过夜后放入﹣79℃保存。
1.2.2 病毒分离培养将生长良好的F81细胞消化成细胞悬液,按2%同步接种病料上清,用含10%血清的MEM于37℃、5% CO2培养箱中培养,同时设立正常细胞对照,每天观察细胞病变的情况。当细胞病变达80%左右收毒,反复冻融3次后置于﹣79℃保存备用。
1.2.3 DNA提取及PCR鉴定合成检测引物后[14]于第三代细胞培养物中提取DNA进行PCR鉴定。反应总体积20L,模板1L,2 LA Taq酶10L,上、下游引物各1L,加dd H2O至总体积20L。PCR扩增程序94℃预变性4 min;98℃变性10 s,55℃退火30 s,72℃延伸1min,32个循环;最后72℃延伸6min。扩增后的产物用0.9%琼脂糖凝胶电泳进行鉴定。
1.2.4 血凝试验(HA)使用96孔微量血凝板对RDPV阳性第3代细胞培养物上清进行HA测定,每孔先加入25L PBS缓冲液(pH 7.2),每排第1孔中加入25L的抗原,吹打混匀后吸25L至第2孔,依次倍比稀释至第11孔,弃掉25L。第12孔作为红细胞对照组,实验设置2个重复和阳性对照组。每孔补加25L PBS,最后每孔加入50L 1%猪红细胞悬液,震荡均匀后于4℃静置45 min后进行判定。将100%的红细胞凝集判为阳性标准,对应的稀释倍数即为该病毒的血凝效价。
1.2.5 病毒TCID50测定 调整F81细胞浓度约为2 105个/mL,每孔100L均匀铺于96孔培养板中,同时将收取的有典型病变的第3代细胞培养液依次按10倍做倍比稀释,分别接种于F81细胞96孔培养板中,每个滴度做8个重复,100L/孔,并设正常细胞对照组,观察并记录4 d内的病变孔数,按Reed-Muench法计算病毒TCID50。
1.2.6 间接免疫荧光(IFA)首先将F81细胞均匀铺在96孔板并按体积的5%同步接种分离株病毒,于37℃,5% CO2培养箱中培养;根据细胞状态,弃上清液,用PBST洗3次;依次进行固定(加入4%多聚甲醛4℃感作45 min,用PBST清洗3次)、穿透(用0.1% Triton-X100室温感作15 min,用PBST清洗3次)、封闭(加入4% FBS,37℃封闭1 h)等步骤,弃掉封闭液,加入一抗并放入湿盒,4℃孵育过夜。用PBST清洗3次加入1:200稀释的二抗100L。室温避光孵育1 h,清洗3次后,于荧光倒置显微镜中观察。同时设未接种病毒的细胞作为阴性对照。
1.2.7 VP2基因克隆及遗传进化分析 参照Genbank上登录号为(GQ857614)的RDPV株的VP2基因序列,利用Primer 5.0软件设计引物,扩增目的片段大小为1 755 bp,引物序列为R:ATGAGTGATGGAGCAGTTCAAC(5′~3′),F:-TTAATATAATTTTCTAGGTGC TAGTTGAG(-5′~3′),PCR反应体系为50L:模板2L,2 LA Taq 25L,上、下游引物各1L,加ddH2O至总体积50L。PCR扩增中关键程序:退火30 (s55℃);最后72℃延伸10 min。反应结束后用1%琼脂糖凝胶电泳进行鉴定。
1.2.8 克隆测序及遗传进化分析VP2基因扩增后产物经胶回收后连接到Peasy-T1克隆载体上,转化至DH-5感受态细胞并于含有氨苄青霉素的LB培养皿上37℃,180 r/min过夜培养。次日挑取单克隆菌落进行鉴定、扩大培养并提取阳性质粒,用限制性内切酶HindⅢ和XhoⅠ进行双酶切鉴定。将酶切鉴定正确的重组质粒送往吉林省库美生物科技有限公司测序并进行拼接。登录NCBI用BLAST选取与VP2基因序列相似性高的核苷酸序列进行同源性分析,用DNASTAR中的MegAlign程序对分离株VP2基因序列与GenBank发布的其他19株肉食兽细小病毒VP2基因序列进行分析并利用Mega7.0软件用最大似然法(Maximum Likelihood)构建系统发育树。
2 结果
2.1 病毒分离
正常细胞轮廓清楚,形态大小较均一;接种RDPVJL3毒株后的F81细胞,细胞变得肿大,呈拉网样典型的病变,见图1。

图1 RDPV-JL3感染F81细胞的形态学观察Fig.1 The morphology of F81 cells infected by the RDPVJL3
2.2 PCR鉴定
PCR扩增产物经1%琼脂糖凝胶电泳检测,在573 bp处出现特异性条带,与预期目的条带片段大小一致,见图2。
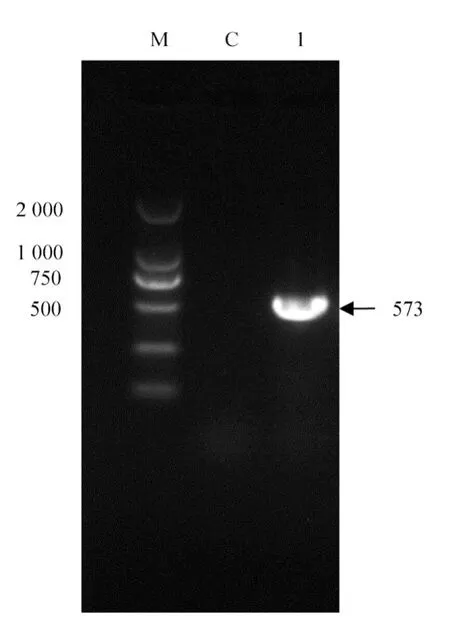
图2 RDPV-JL3 PCR鉴定结果Fig.2 The PCR-amplified form the RDPV-JL3
2.3 HA结果
RDPV-JL3第3代细胞病毒培养物的血凝效价保持在28以上。
2.4 TCID50结果
利用Reed-Muench法算得RDPV-JL3分离株的病毒含量为105.67 TCID 50/mL。
2.5 间接免疫荧光结果
荧光显微镜下分离毒株RDPV-JL3可见特异性绿色荧光;正常细胞对照未见特异性荧光,证实了该分离毒株为RDPV,见图3。

图3 RDPV-JL3的IFA结果Fig.3 The IFA result for the RDPV-JL3 strains of the parvovirus infected F81 cells
2.6 酶切鉴定
酶切鉴定与预期的目的条带片段大小一致,见图4。
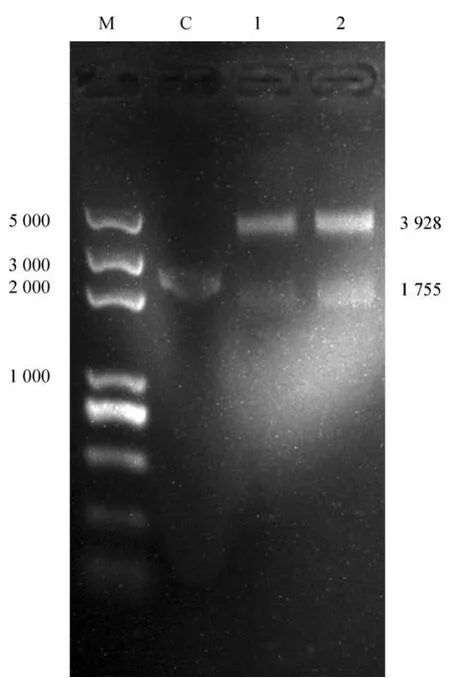
图4 RDPV-JL3基因重组质粒酶切鉴定结果Fig.4 The identification of the pEASY-T1-VP2 vector by restriction endonuclease digestion
2.7 VP2基因进化分析结果
将分离毒株RDPV-JL3 VP2序列与GenBank中公布的FPV、MEV、CPV和RDPV等19株肉食兽细小病毒VP2基因序列进行比对,通过对该株病毒与19个参考毒株VP2蛋白10个关键氨基酸位点进行比较分析,结果表明其80R、87M与FPV、MEV的氨基酸位点相同,根据其93N、103A、297S和426N位置氨基酸特性确定该株细小病毒属于CPV-2株,未发现有S27T、S297A和I418T等氨基酸位点的同义替换,见表1。核苷酸进化树分析结果表明,RDPV-JL3株与RDPV HeB10-3(登录号KJ194463)及RDPV-17-JL-9(MH581183)株处于同一进化分支,与CPV-Vac1疫苗株亲缘关系比较近,见图5。

表1 RDPV-JL3株与GenBank中已知病毒的关键氨基酸比较Table 1 The comparison of key amino acids of known viruses between the RDPV-JL3 strain and GenBank

图5 RDPV-JL3 VP2基因核苷酸序列的遗传进化树Fig.5 Phylogenetic relationships among 19 parvoviruses of carnivores isolates based on the nucleotide sequences of the VP2 gene
3 讨论
细小病毒科是一种小的DNA病毒,具有线形单链DNA基因组,全长约5 kb,两端有发夹结构。该病毒有两个主要的开放阅读框,一个编码两个非结构蛋白(NS1和NS2),另一个编码衣壳蛋白VP1和VP2[15]。其中,VP2蛋白与病毒的致病性、毒力以及血凝特性密切相关。VP2蛋白上几个关键氨基酸位点的改变就可以影响病毒的抗原性、宿主范围和血凝特性,如VP2蛋白的aa77、aa88、aa370、aa375和aa377等位点发生突变后,细小病毒便失去血凝特性;aa426和aa297位的氨基酸特征是鉴定CPV-2变异株(CPV-2a、CPV-2b、New CPV-2a、New CPV-2b和CPV-2c)的关键因素[16,17]。1979 1980年,在美国,从狗身上发现了一种抗原性变异株,命名为CPV-2a。与原CPV-2不同的是VP2有5个氨基酸突变(Met87Leu、Ile101Thr、Ala300Gly、Asp 305 Tyr和Val 555 Ile)[18],与此同时,另外两个变异株CPV-2b(Asp 426)和CPV-2c(Glu 426)分别于1984年和1996年出现[19]。在过去的几十年中,VP2蛋白Ser 297 Ala突变的CPV-2a和CPV-2b被命名为NewCPV-2a和NewCPV-2b[20]。
本试验从貉肠道中成功分离出一株RDPV,经NCBI对其VP2基因序列进行BLAST分析表明,分离株RDPV-JL3与RDPV HeB10-3(KJ194463)相似性为100%,与RDPV-17-JL-9株相似性为99.94%。将其与Genbank上已公布的CPV分离株、CPV-Vac1疫苗株、RDPV分离株、FPV分离株和MEV分离株等19株已知毒株进行氨基酸同源性比较以及遗传进化分析,结果发现所分离毒株与RDPV-17-JL-9株和CPVHeB10-3株的关键氨基酸位点一致,且根据103A、297S、300A和426N几个关键氨基酸位点判定为CPV-2型。RDPV-JL3分离株与RDPV-17-JL-9分离株相比仅在S192F不一致,说明本次分离毒株与RDPV HeB10-3(KJ194463)和RDPV-17-JL-9株具有共同的起源;与同型的美国野生毒株CPV-b、CPV-d对比分析,在D375N位有突变,可能是地理因素造成的;与CPV-Vac1疫苗株相比,存在D375N、Q386K的突变,这是否对疫苗防疫失败有影响还需要进一步深入研究。从系统进化树上来看,RDPV-JL3与RDPV-17-JL-9株、CPVHeB10-3株处于同一进化分支并与CPV-Vac1疫苗株亲缘关系比较近,推测可能是由于养殖场对犬使用犬细小病毒弱毒疫苗(CPV-2型)进行免疫,免疫后的犬持续性排毒,被CPV-2易感的貉子接触感染发病[21]。
