环境因素对盾形陀螺珊瑚摄食及共生藻光合生理的影响
2021-10-12郑子豪张艳苹
沈 城,郑子豪,张艳苹,刘 丽
(1.徐闻国家级珊瑚礁保护区管理局,广东 湛江 524143;2.广东海洋大学水产学院,广东 湛江 524088)
盾形陀螺珊瑚 (Turbinaria peltata) 隶属于刺胞动物门珊瑚虫纲石珊瑚目陀螺珊瑚属,是最原始的后生动物类群之一[1],一般生活在赤道及其附近的热带、亚热带地区,多生长于水体平静而清澈的岩礁、平台、斜坡、崖面、凹缝。珊瑚有自养和异养2 种营养方式。自养时珊瑚通过共生虫黄藻的光合作用供给营养,异养时捕食有机颗粒[2]。黄晖等[2]认为,在环境适宜条件下,光合自养是健康造礁石珊瑚的主要营养方式,当珊瑚自养营养供给受限时,异养营养可在一定程度上弥补营养缺失,甚至成为主要营养方式。关于环境因子对珊瑚摄食的影响已有报道,滨珊瑚(Porites compressa)摄食率随温度的升高而降低,团块滨珊瑚(Porites lobata)摄食率随温度的升高而升高[3-4],萼形柱珊瑚(Stylophora pistillata)摄食率随海水pH 下降而降低[5]。
在全球珊瑚礁逐渐退化的背景下,研究珊瑚在不同胁迫条件下的行为,了解珊瑚的环境可塑性对珊瑚资源保护意义重大。目前,关于环境因子对珊瑚摄食行为影响研究主要集中在温度、pH 方面[6-9],而未涉及其他环境因子。多数研究只关注不同环境条件下珊瑚摄食率变化,有关环境变化影响珊瑚摄食行为的具体机制,仅见Ferrier-Pagès 等[10]的温度升高情况下珊瑚摄食率及虫黄藻光合速率的变化,但未深入探究。研究珊瑚自养与异养行为之间的联系有助于更全面解读环境变化对珊瑚生存的影响,进而了解珊瑚抵御环境变化的生理机制。
轮虫有繁殖速度快、活动缓慢、易培养、营养价值高、无残饵污染等优势,已广泛应用于海水养殖业[11-12]。研究表明,高温条件下珊瑚对轮虫(Brachionus plicatilis) 的摄食率显著提高[13];本课题组对盾形陀螺珊瑚饵料筛选研究表明,盾形陀螺珊瑚对轮虫有较好的适应性[14],轮虫有发展为珊瑚人工养殖饵料的潜力。本研究分析不同环境条件对珊瑚摄食轮虫速率的影响,为盾形陀螺珊瑚优化养殖提供基础数据。
1 材料与方法
1.1 材料
盾形陀螺珊瑚母体1 株,于2020 年5 月采自徐闻珊瑚礁国家级自然保护区(109°55'18.28",20°16'30.55"),采样点深度约4 m。运回实验室暂养在水温25 ℃、pH 8.1、盐度30、钙400 mg/L、镁1 250 mg/L 的50 L 珊瑚养殖箱中,光源为全光谱LED 珊瑚灯(Maxspect),光照时间为7:00–19:00,缸底有效辐射约为100 μmol/(m2·s)。暂养7 d,待珊瑚触手正常伸展后,于21:00 将其切割成大小相近的珊瑚小穗,用阿隆发胶水将小穗固定在塑料底座上,培养至珊瑚触手正常伸展,开始投喂实验。珊瑚恢复期间不喂食。
1.2 温度对盾形陀螺珊瑚摄食及共生藻光合生理的影响
将9 支珊瑚小穗集中置50 L 养殖箱中驯养,待珊瑚恢复正常生长状态后,将珊瑚小穗随机放入3个10 L 实验缸。温度设置24(对照)、28、32 ℃。使用鱼缸加热棒(SUNSUN)调节水体温度,各实验组温度以每天1 ℃的速度升至设置温度。测量摄食速率前停饲24 h。实验持续29 d,前8 d 为温度调整期,各实验组到达指定温度后,驯化7 d,开始摄食实验。摄食试验共进行14 d,实验过程中每日测定珊瑚的摄食率。养殖实验结束后测定珊瑚净光合速率、呼吸速率及虫黄藻密度。
1.3 pH 变化对盾形陀螺珊瑚摄食及共生藻光合生理的影响
用CO2发生器或石灰水 [Ca(OH)2] 调节海水pH。每日检测海水pH 值,使海水pH 维持在设计范围。随机取驯化的珊瑚小穗移入10 L 实验缸,每缸3 支小穗。各实验缸pH 条件见表1。以珊瑚采集海域实际pH 值8.0 为对照组。其他参数测定同1.2。实验共进行14 d。养殖实验结束后测定珊瑚摄食率、净光合速率、呼吸速率及虫黄藻密度。

表1 各处理组pH 条件Table 1 pH condition of each treatment group
1.4 不同氮源及氮磷比对盾形陀螺珊瑚摄食及共生藻光合生理的影响
将15 支大小相近的珊瑚小穗随机分配至5 个10 L 实验缸中,每缸3 支,除无机盐浓度外,各实验缸条件与珊瑚养殖箱的条件相同,每2 d 全换水1 次,更换后实验缸中无机盐浓度不变。各处理组无机盐条件见表2。实验为期14 d。养殖实验结束后测定珊瑚摄食率、净光合速率、呼吸速率及虫黄藻密度。

表2 各组无机盐条件Table 2 Inorganic salt conditions for groups
1.5 相关参数测量
1.5.1摄食率测定 在有1 L 海水的烧杯中添加轮虫至密度为12 000 只∙L-1,于9:00 将所饲喂珊瑚移入,1.5 h 后计数剩余轮虫量,另设相同轮虫密度烧杯作对照。
1.5.2光合速率及呼吸速率的测定 珊瑚小穗分别移至500 mL 盛满过滤海水(过孔径0.45 µm 滤膜)的烧杯中,烧杯中过滤海水条件与实验缸一致。珊瑚表面积通过锡纸法获得。用(YSI PROODO)溶氧仪测定珊瑚净光合速率(Pn)、呼吸速率(RD):
1)在100 µmol·m−2·s−1光照条件下测定Pn;
2)于黑暗条件下1 h 后测定RD,测量前按照仪器操作说明,在氮气饱和海水和空气饱和海水(分别为0%和100%氧饱和度水平)中对溶氧仪进行校准。
1.5.3虫黄藻密度测定 用冲牙器从骨骼中剔取珊瑚组织,放入5 mL 经孔径0.45 µm 滤膜过滤的海水中,用研磨棒充分研磨,组织混匀液在4 000g条件下离心10 min,去上清,所得藻体用1.5 mL过滤海水重新悬浮、混匀,用血细胞计数板镜检获得悬浮液中虫黄藻数量,并均一化至珊瑚面积。
1.6 统计分析
用SPSS17.0 软件对摄食率、共生虫黄藻密度、净光合速率、呼吸速率等数据进行统计分析,结果以平均值±标准误表示,显著性检验采用单因素方差分析(one-way ANOVA),并采用Duncan 多重比较检验。显著水平α设为0.05 或0.01。
2 结果
2.1 温度对盾形陀螺珊瑚摄食及共生藻光合生理的影响
由图1 可知,温度对盾形陀螺珊瑚摄食率有显著影响,在9 d 内,各组珊瑚摄食节律相似,10 d后,32 ℃条件下摄食率呈持续下降趋势。喂食实验结束时,32 ℃条件下珊瑚摄食速率比峰值降低约29.6%。此外,温度的提高可增加珊瑚的摄食率与24 ℃相比,28 ℃和32 ℃时珊瑚的峰值摄食率分别增加14.81%和20.83%,28 ℃和32 ℃处理组珊瑚摄食率均显著高于24 ℃组(P< 0.05)。

图1 不同温度下单位珊瑚触手的摄食率变化Fig.1 A coral polyp feeding rate changes at different temperatures
由图2 可知,在高温条件下,摄食后共生虫黄藻的密度显著降低。在28 ℃时,珊瑚共生的虫黄藻数比对照组 (24 ℃) 降低19.89%,为 (3.63±0.14)×106cm-2,而在32 ℃时,虫黄藻数量比对照组降低49.90%,为 (2.04±1.09)×106cm-2。而净光合速率随着温度的升高而升高,在28 ℃下,珊瑚净光合速率比对照组提升 24.45%,呼吸速率提升32.24%,光合呼吸比下降5.18%。在32 ℃下,珊瑚净光合速率比对照组提升71.81%,呼吸速率提升97.61%,光合呼吸比下降20.25%。
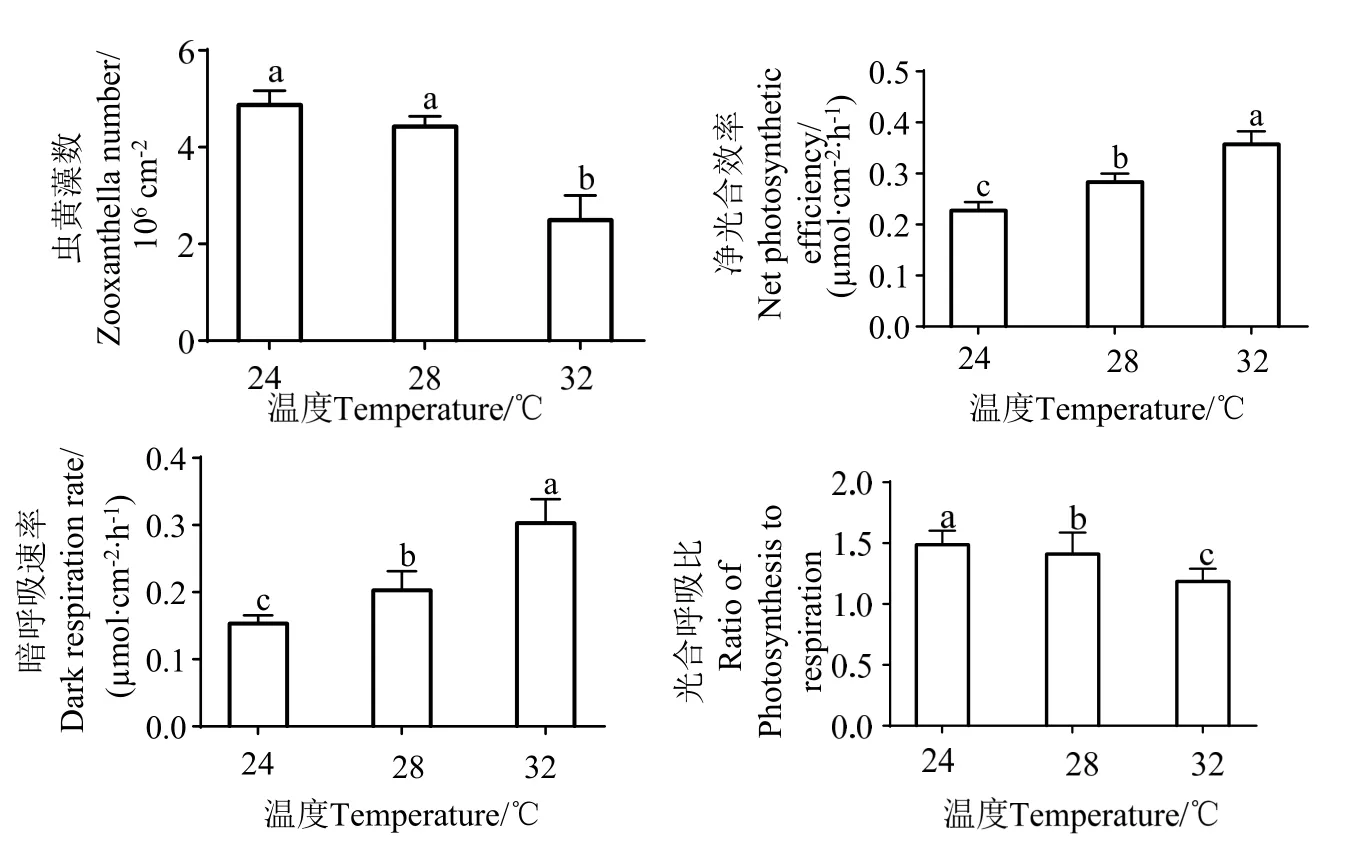
图2 不同温度下珊瑚水螅体摄食率、虫黄藻密度及光合速率变化Fig.2 Changes of coral polyp feeding rate,zooxanthellae density and photosynthetic rate at different temperatures
2.2 pH 对盾形陀螺珊瑚摄食及共生藻光合生理的影响
由图3 可知,pH 降低时盾形陀螺珊瑚的摄食率差异显著 (P< 0.05),摄食率随着pH 的下降而下降,与对照组 (pH 8.0) 相比,摄食率在pH 为7.8时下降20.00%,pH 为7.6 时下降34.41%。同时,pH 变化对珊瑚呼吸速率、净光合速率以及虫黄藻密度也有显著影响,pH 降低时珊瑚虫黄藻密度、净光合速率和呼吸速率均显著降低 (P< 0.05)。pH 7.8 时,珊瑚共生虫黄藻数量从 (4.66±1.56)×106cm-2下降到 (3.45±0.33)×106cm-2,约下降25.97%。pH 7.6 时,珊瑚虫黄藻数量由(4.66±1.56)×106cm-2下降到(2.62±0.33)×106cm-2,约下降43.78%。pH 为7.6、7.8 时,珊瑚净光合速率分别为3.45、2.62 μmol·cm-2·h-1,与对照组相比,分别下降20.09%和50.22%。低pH 对珊瑚呼吸速率也有显著影响,在pH 为7.6、7.8 条件下,与pH 8.0 相比分别下降20.83%和47.28%。pH 8.2 组适当提高pH 对珊瑚摄食率、共生虫黄藻密度、净光合速率、呼吸速率影响不大,与pH 8.0 组间均无显著差异(P> 0.05)。
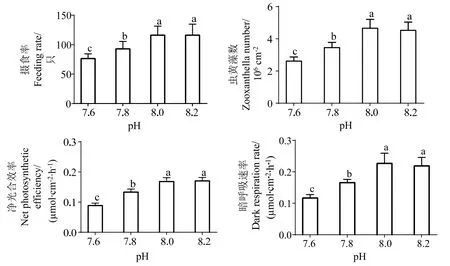
图3 不同pH 条件下珊瑚水螅体摄食率、虫黄藻密度及光合速率变化Fig.3 Changes of coral polyp feeding rate,zooxanthellae density and photosynthetic rate under different pH conditions
2.3 不同氮源及氮磷比对盾形陀螺珊瑚摄食及共生藻光合生理的影响
由图4 可知,提高无机盐浓度可增强造礁石珊瑚的摄食行为,NO3、NH4和NH4+P 组珊瑚的摄食率比对照组分别提高32.27%、29.00%和22.72%,NO3+P 组珊瑚摄食率提升最多,为34.42%,不同处理组间无显著差异 (P> 0.05),各处理组均与对照组有显著差异 (P< 0.05)。
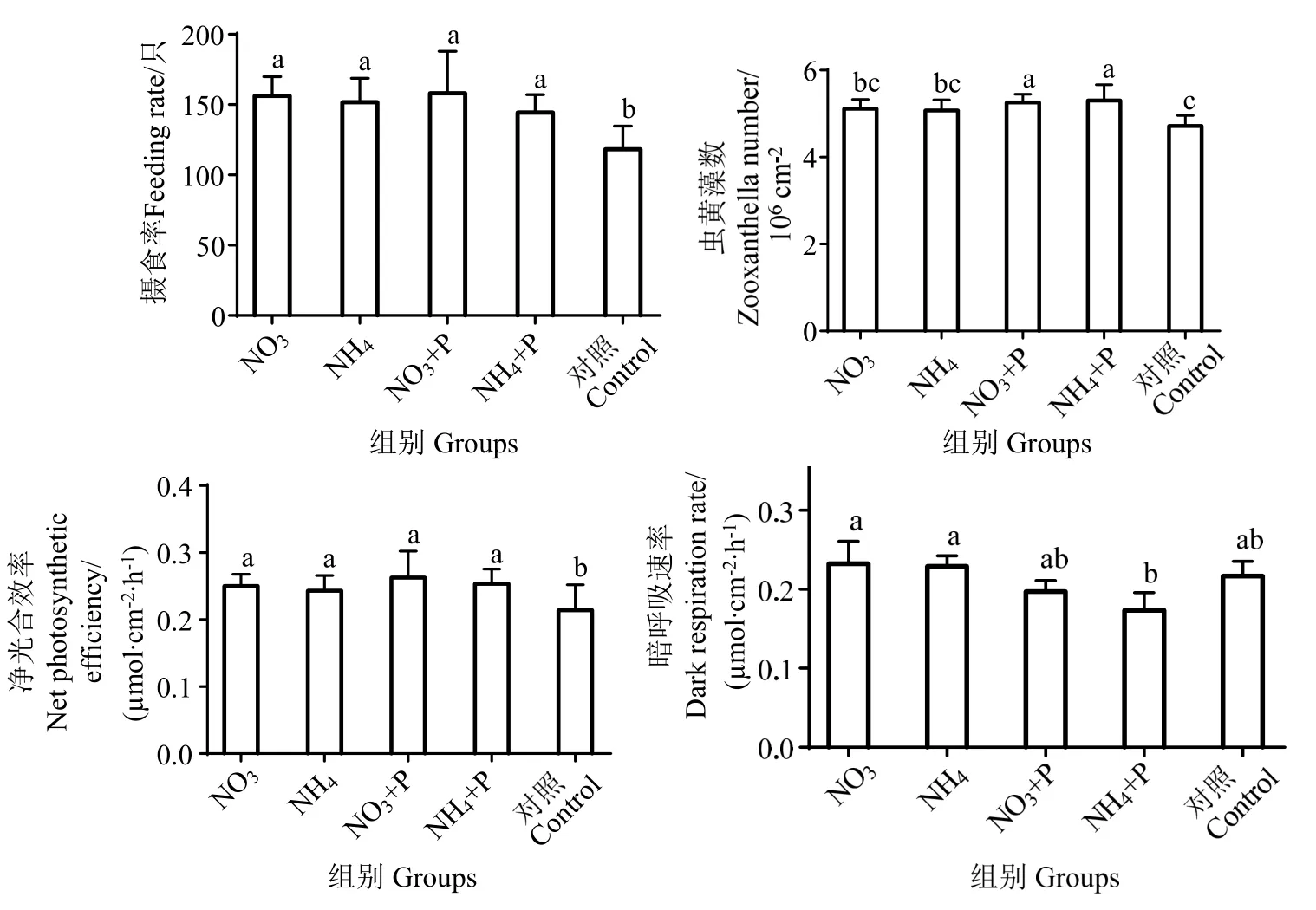
图4 不同氮源及氮磷比对盾形陀螺珊瑚生活指标的影响Fig.4 Different letters in the same group indicate significant differences
氮磷失衡 (100∶1) 条件下珊瑚共生虫黄藻数量与对照组相比略微提升,但差异不明显 (P>0.05),氮磷均衡 (20∶1) 条件下珊瑚共生虫黄藻数量与对照组相比显著提升(P< 0.05)。
与对照组相比,添加无机营养条件下珊瑚的净光合速率显著提高(P< 0.05),NH4+P 组平均值提升最多,为17.11%,各处理组间无显著差异 (P>0.05)。无机营养同样对虫黄藻的呼吸作用有影响,其中NH4+P 组降低最多,为25.02%。
3 讨论
健康的造礁石珊瑚多生长于光照充足的海域,其新陈代谢的能量需求主要来源于虫黄藻的光合作用产物[15]。珊瑚的异养营养在氮、磷等必需元素的获取中起关键作用[16],尤其是当珊瑚自养营养的供给受限时,异养营养可一定程度上弥补其营养缺失,甚至成为主要营养方式[3]。而异养营养过程受多种环境因子的影响,所以探究环境变化对珊瑚异养营养的影响对应对环境变化导致珊瑚白化现象有重要意义。
本研究中,随着温度的升高,盾形陀螺珊瑚共生虫黄藻数量下降,摄食率提高,说明珊瑚在高温胁迫环境下,需要通过增加异养营养来弥补因虫黄藻丢失而导致的能量供应不足。Ferrier-Pagès 等[10]认为,异养摄食有助于珊瑚维持其与虫黄藻的共生关系,异养摄食可能为虫黄藻提供氮和其他必要的营养物质,或减少了珊瑚对光合产物的需求,使虫黄藻有更多营养物质维持自身正常的生理过程[17]。Palardy 等[18]研究表明,表孔珊瑚(Montipora capitata)在高温条件下摄食行为显著增加。但不同造礁石珊瑚在高温环境下表现出的摄食行为不一致,Ferrier-Pagès 等[10]对比Stylophora pistillata、Turbinaria reniformis、Galaxea fascicularis3 种珊瑚在高温条件下摄食率的变化情况,发现不同珊瑚在高温条件下的摄食率存在差异,Stylophora pistillata在31 ℃条件下摄食率显著下降,而其他两种珊瑚的摄食率在31 ℃条件下有所上升。本研究中,温度升高显著提高了盾形陀螺珊瑚的摄食率,在28 ℃条件下珊瑚虫黄藻数量并未出现显著下降,但摄食率依旧有所提升,这说明温度不仅影响了珊瑚的能量供应,还可能通过提高珊瑚共生藻光合速率间接增加珊瑚用于捕食的能量分配。光合速率与呼吸速率体现了珊瑚共生虫黄藻的光合能力,这两项数值的变化可能是珊瑚摄食率变化的原因之一。
除温度外,pH 对造礁石珊瑚的异养营养也有一定的影响。在酸化条件下,珊瑚钙化需要消耗更多能量[19],进而减少珊瑚在捕食方面的能量分配,导致储存在珊瑚组织中的营养物质被快速消耗,最终导致捕食量和钙化的减少。本研究表明,在低pH情况下,盾形陀螺珊瑚对轮虫的摄食率显著降低,同时珊瑚共生虫黄藻数量、净光合速率、呼吸速率均显著降低,说明低pH 条件下,盾形陀螺珊瑚自养营养及主动摄食能力均受较大影响。这与Edmunds 等[7]酸化条件下所喂食珊瑚的组织量在3周内减少10%的结果相吻合,而在正常情况下,异养营养的输入使珊瑚组织量增加[20]。因此推测,异养营养的输入可在一定时间内维持珊瑚钙化速率,但长期暴露在酸化条件下,珊瑚摄食率的下降导致钙化速率大幅度下降,同时随着组织中营养物质的消耗,珊瑚捕食能力逐渐减弱。
本研究中,无机营养可显著增强盾形陀螺珊瑚的异养摄食率,并可提高珊瑚共生虫黄藻数量,使珊瑚净光合速率增加。Leal 等[21]指出,海葵(Aiptasiapallida)的共生虫黄藻碳移位率和宿主猎物摄取呈正相关,认为自养能量是宿主满足摄食和消化代谢需求的必需营养方式。本研究有相似结果,虫黄藻数量及光合作用速率的增加可为珊瑚提供更多的自养营养,进一步增强珊瑚捕食行为;同时,长时间温度过高 (32 ℃) 将导致珊瑚在10 d 后共生虫黄藻数量减少、摄食能力显著下降。这两者间的相同点是虫黄藻数量的变化,所以当无机营养盐浓度升高时,随着虫黄藻光合作用的增强,更多的光合产物可为珊瑚提供更多的能量,这可能影响珊瑚的能量分配过程,当能量充足时珊瑚会分配更多的能量用于捕食行为,以获取更多的“额外”营养物质。
在珊瑚中,寄主组织和共生体的N、P 比率约为50∶1[22],远高于海水中N、P 比16∶1。本研究中,氮磷比20∶1 条件下,珊瑚共生虫黄藻数量与对照组相比显著提升 (P< 0.05),同时提升幅度比氮磷比100∶1 条件下高,说明高无机盐条件下氮磷比维持在一定比例有利于维持珊瑚健康的生理状态。有研究指出,不平衡的硝酸盐和磷的比例加剧了几种珊瑚的磷饥饿和白化[23-24],同时减少了共生虫黄藻向珊瑚转移的光合产物数量[25]。
本研究中,温度、pH、无机盐浓度均会影响珊瑚的摄食率,表明珊瑚摄食能力会因环境因子变化而变化。因此,珊瑚触手的大小及密度可能并不是影响珊瑚摄食能力的唯一因素,海水环境及珊瑚健康状态与珊瑚的摄食能力密切相关。确定珊瑚最佳的喂食条件有利于提高人工培育下珊瑚的生长速率并缩短育苗时间,有助于进一步推进珊瑚礁修复工作。
